Bone Marrow-Derived HipOP Cell Population Is Markedly Enriched in Osteoprogenitors

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
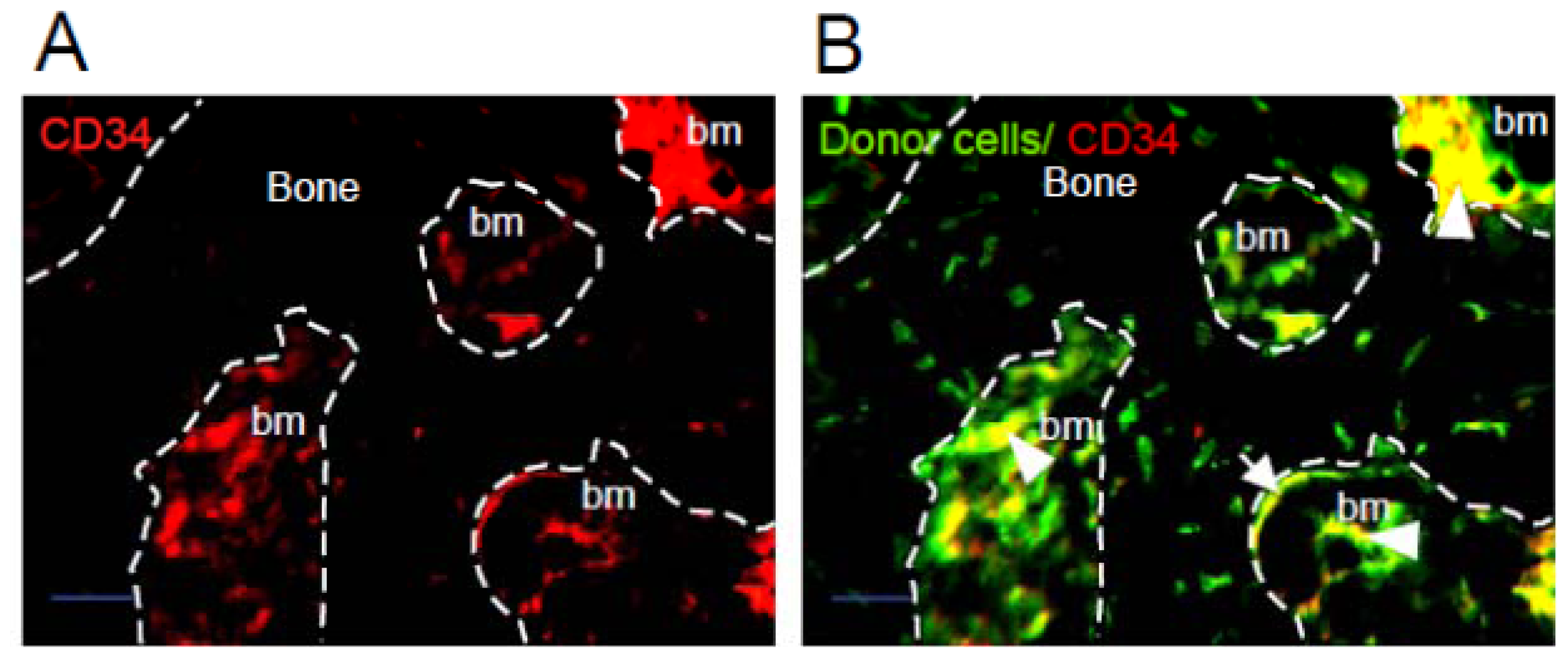
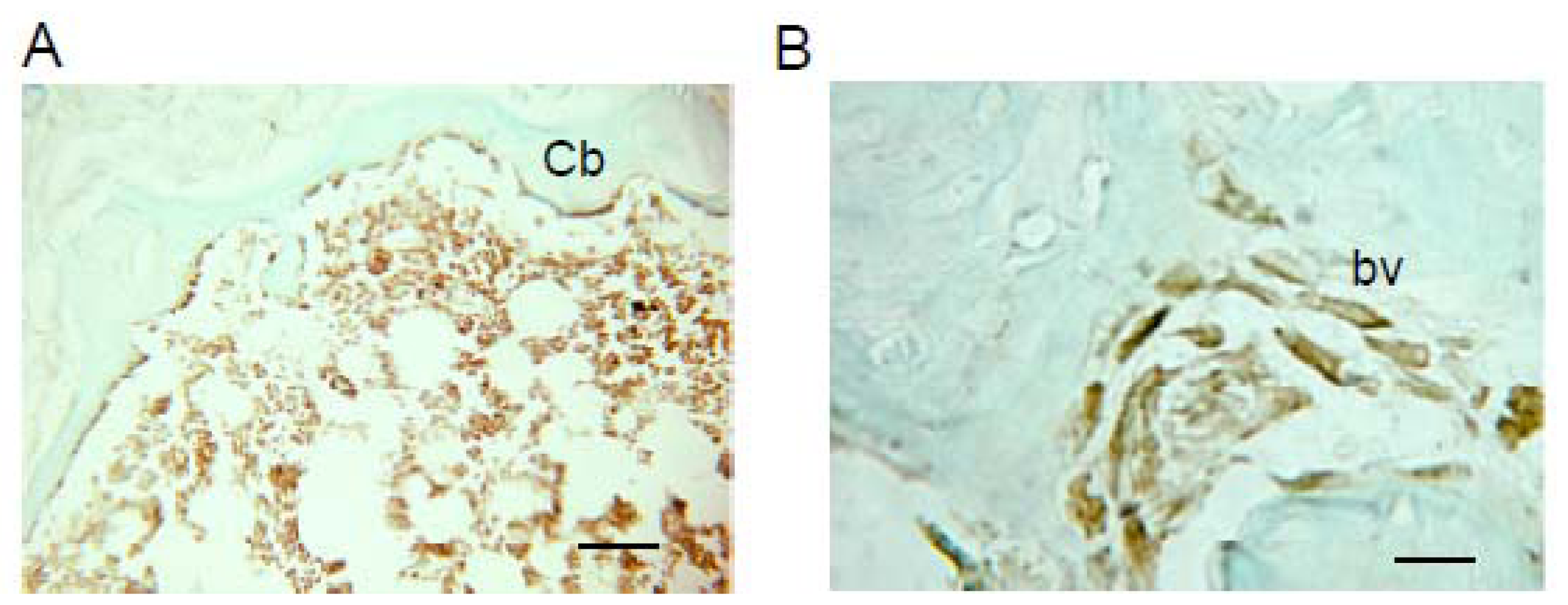
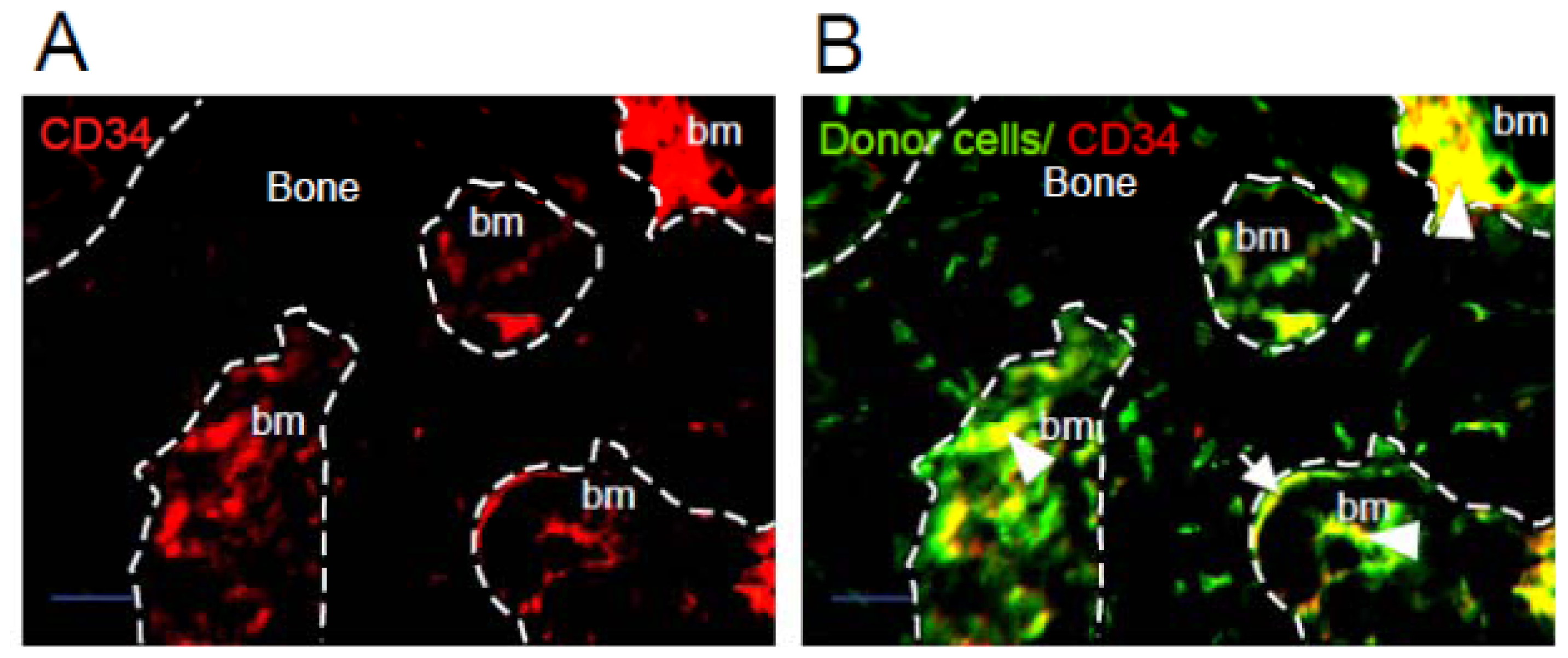
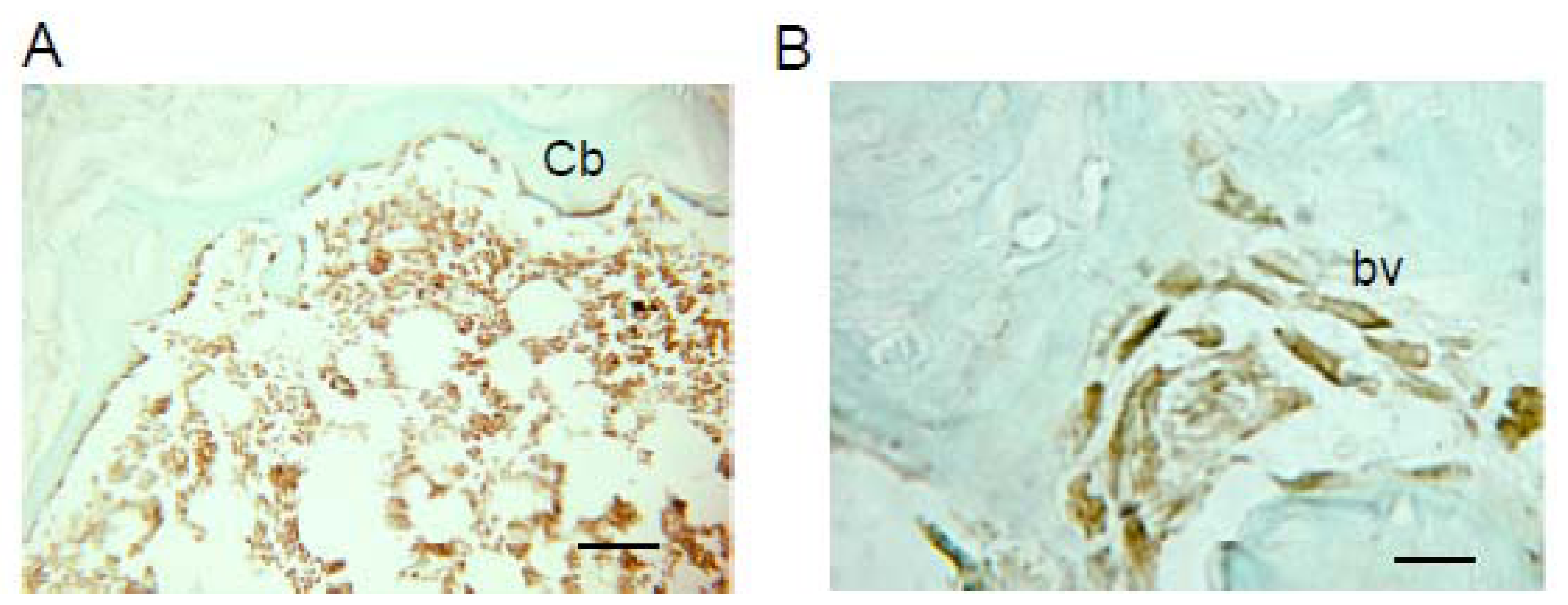
2.1. Donor Cells Generate Multiple Lineages Throughout the Transplants
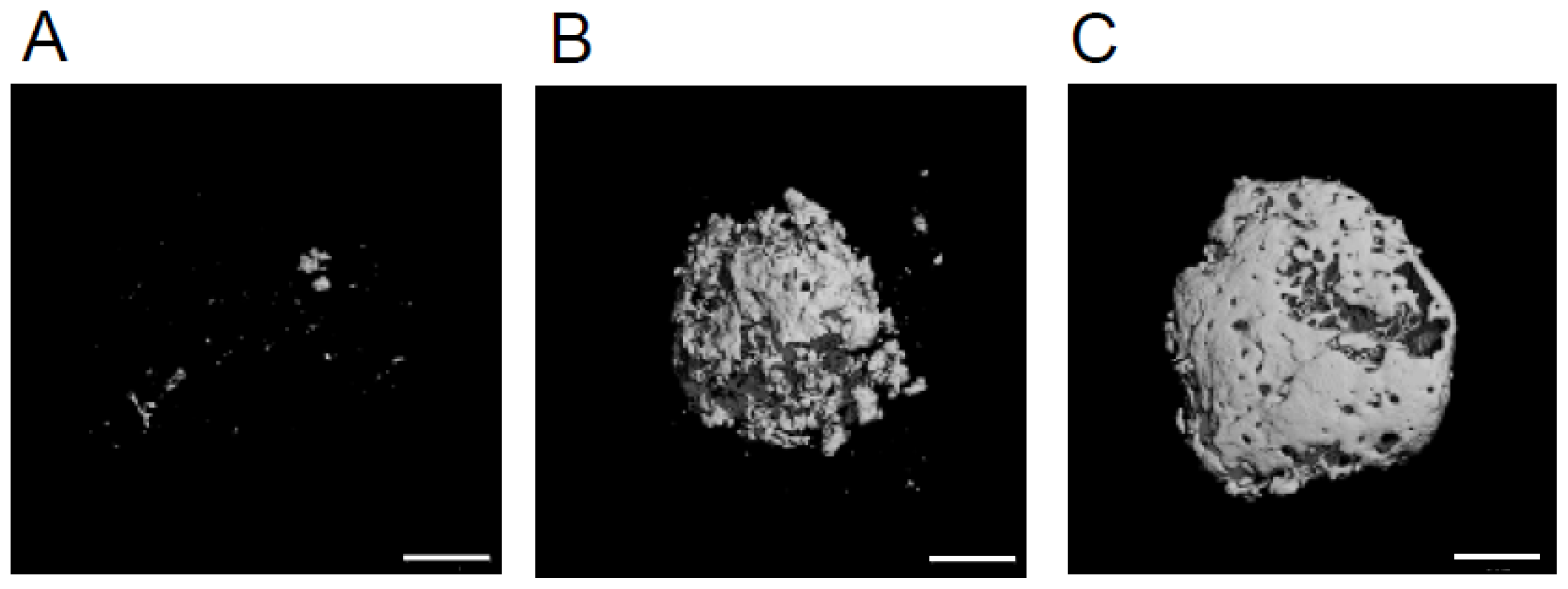
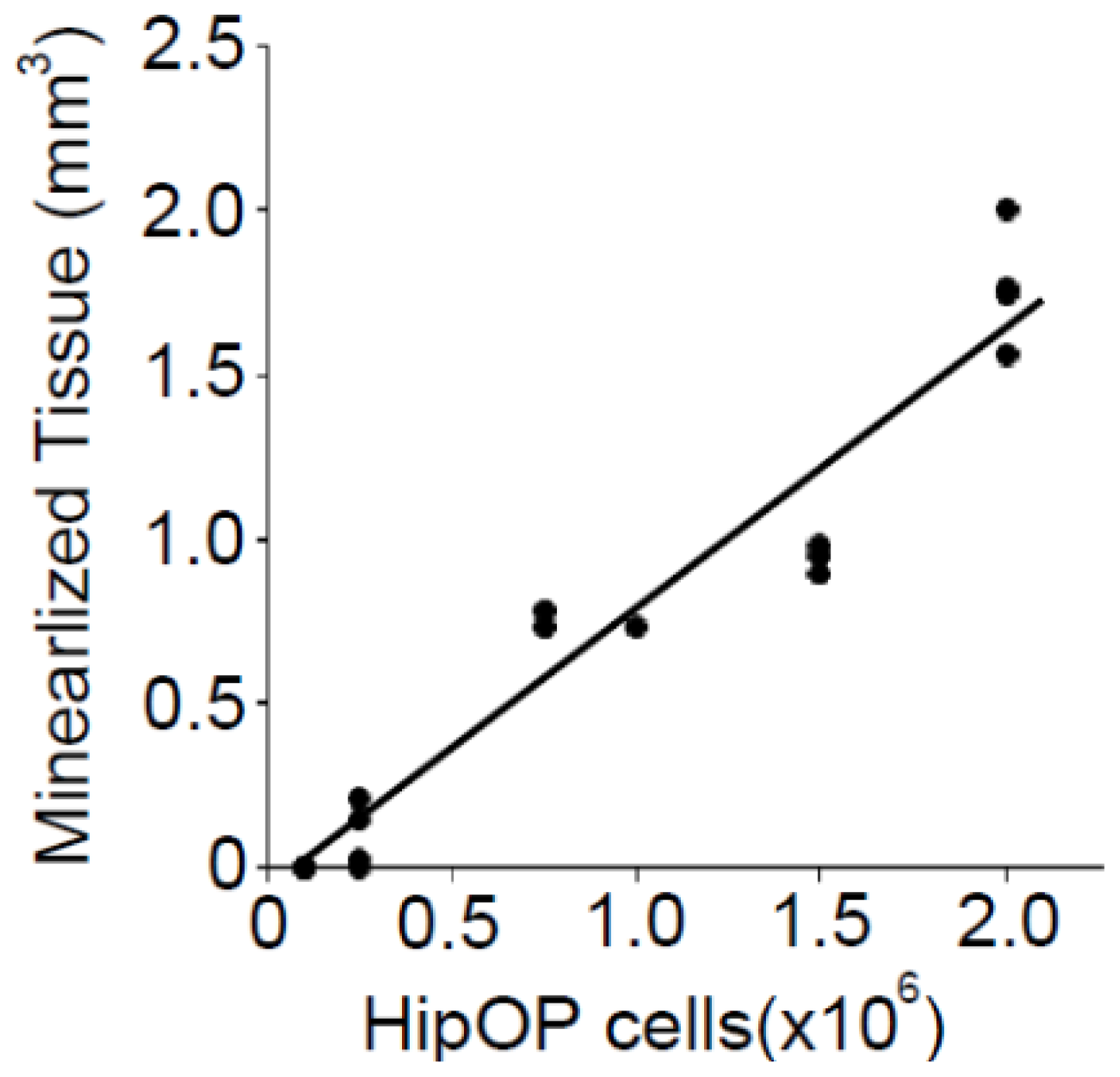
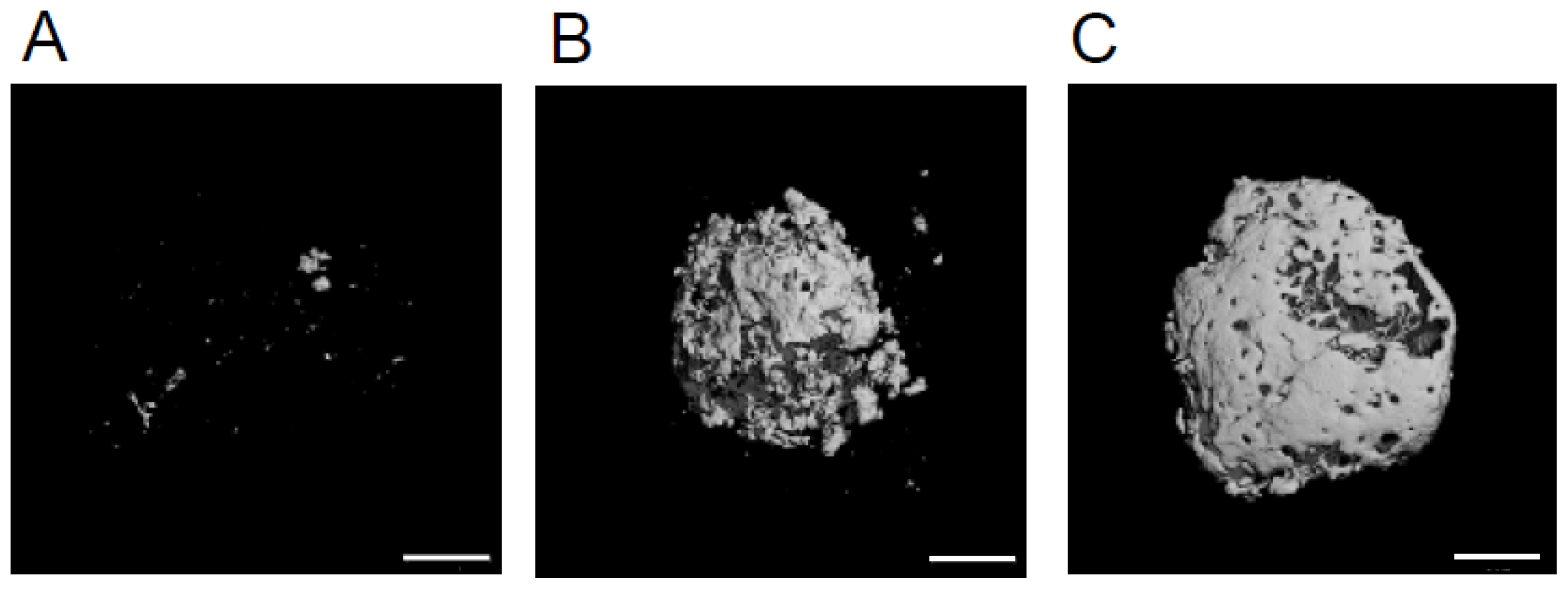
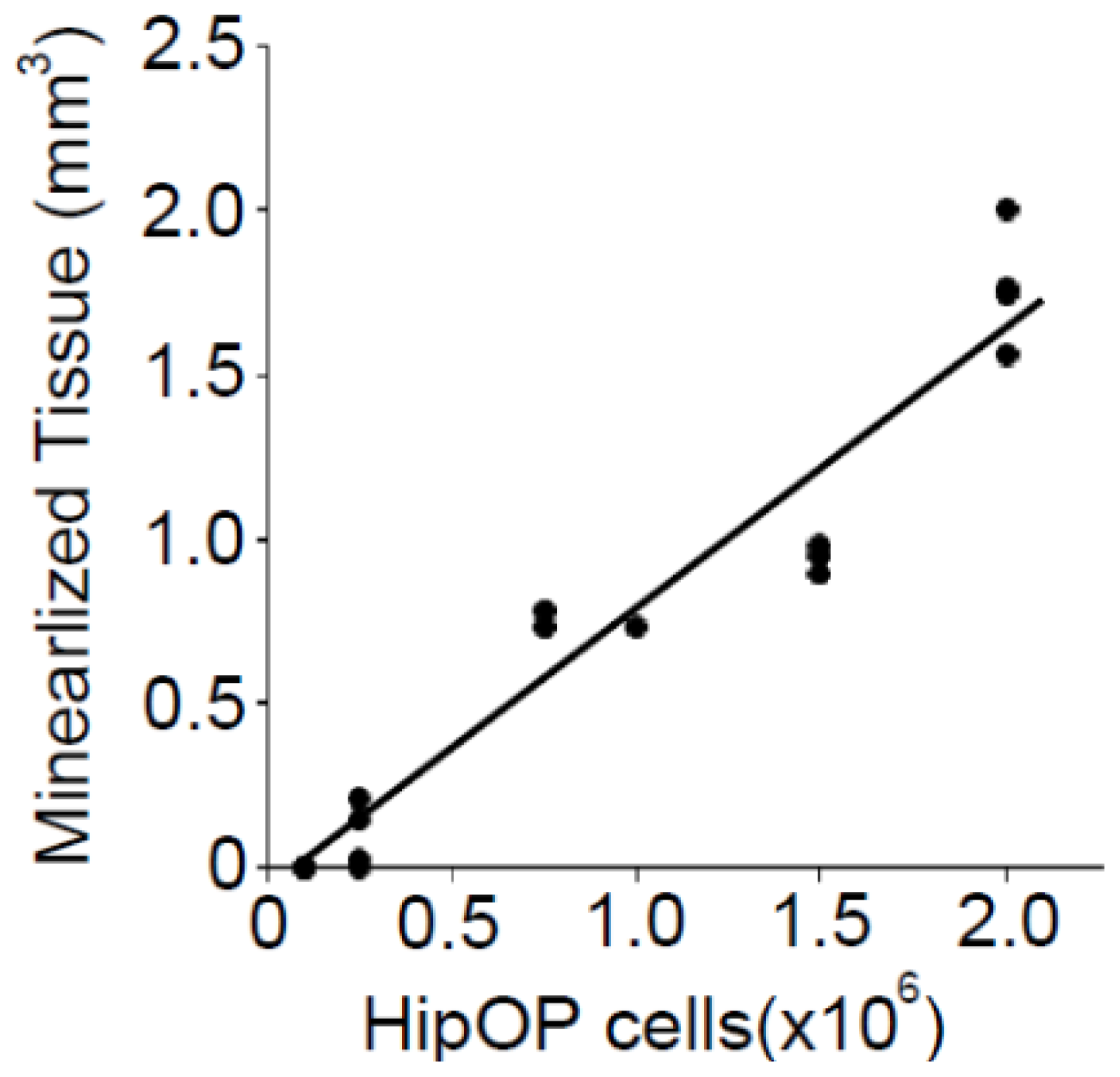
2.2. The Volume of Mineralized Tissue Produced Is Dependent on Transplanted by HipOP Cell Number
3. Discussion
4. Experimental Section
4.1. Isolation of HipOPs
4.2. In Vivo Transplantation
4.3. Micro CT
4.4. Statistical Analysis
5. Conclusions
Acknowledgement
References
- Arai, F.; Suda, T. Maintenance of quiescent hematopoietic stem cells in the osteoblastic niche. Ann. N. Y. Acad. Sci 2007, 1106, 41–53. [Google Scholar]
- Frisch, B.J.; Porter, R.L.; Calvi, L.M. Hematopoietic niche and bone meet. Curr. Opin. Support. Palliat. Care 2008, 2, 211–217. [Google Scholar]
- Kiel, M.J.; Morrison, S.J. Uncertainty in the niches that maintain haematopoietic stem cells. Nat. Rev. Immunol 2008, 8, 290–301. [Google Scholar]
- Friedenstein, A.J.; Gorskaja, J.F.; Kulagina, N.N. Fibroblast precursors in normal and irradiated mouse hematopoietic organs. Exp. Hematol 1976, 4, 267–274. [Google Scholar]
- Pittenger, M.F.; Mackay, A.M.; Beck, S.C.; Jaiswal, R.K.; Douglas, R.; Mosca, J.D.; Moorman, M.A.; Simonetti, D.W.; Craig, S.; Marshak, D.R. Multilineage potential of adult human mesenchymal stem cells. Science 1999, 284, 143–147. [Google Scholar]
- Owen, M.; Friedenstein, A.J. Marrow stromal stem cells. Ciba Found. Symp 1988, 136, 42–60. [Google Scholar]
- Owen, M. Stromal stem cells: Marrow-derived osteogenic precursors. J. Cell Sci. Suppl 1988, 10, 63–76. [Google Scholar]
- Prockop, D.J. Marrow stromal cells as stem cells for nonhematopoietic tissues. Science 1997, 276, 71–74. [Google Scholar]
- Bianco, P.; Kuznetsov, S.A.; Riminucci, M.; Gehron Robey, P. Postnatal skeletal stem cells. Method Enzymol 2006, 419, 117–148. [Google Scholar]
- Phinney, D.G.; Kopen, G.; Isaacson, R.L.; Prockop, D.J. Plastic adherent stromal cells from the bone marrow of commonly used strains of inbred mice: Variations in yield, growth, and differentiation. J. Cell Biochem 1999, 72, 570–585. [Google Scholar]
- Bearpark, A.D.; Gordon, M.Y. Adhesive properties distinguish sub-populations of haemopoietic stem cells with different spleen colony-forming and marrow repopulating capacities. Bone Marrow Transpl 1989, 4, 625–628. [Google Scholar]
- Simmons, P.J.; Masinovsky, B.; Longenecker, B.M.; Berenson, R.; Torok-Storb, B.; Gallatin, W.M. Vascular cell adhesion molecule-1 expressed by bone marrow stromal cells mediates the binding of hematopoietic progenitor cells. Blood 1992, 80, 388–395. [Google Scholar]
- Deryugina, E.I.; Muller-Sieburg, C.E. Stromal cells in long-term cultures: Keys to the elucidation of hematopoietic development? Crit. Rev. Immunol 1993, 13, 115–150. [Google Scholar]
- Itoh, S.; Aubin, J.E. A novel purification method for multipotential skeletal stem cells. J. Cell Biochem 2009, 108, 368–377. [Google Scholar]
- Krebsbach, P.H.; Kuznetsov, S.A.; Satomura, K.; Emmons, R.V.; Rowe, D.W.; Robey, P.G. Bone formation in vivo: Comparison of osteogenesis by transplanted mouse and human marrow stromal fibroblasts. Transplantation 1997, 63, 1059–1069. [Google Scholar]
- Itoh, S.; Itoh, M.; Nishida, K.; Yamasaki, S.; Yoshida, Y.; Narimatsu, M.; Park, S.J.; Hibi, M.; Ishihara, K.; Hirano, T. Adapter molecule Grb2-associated binder 1 is specifically expressed in marginal zone B cells and negatively regulates thymus-independent antigen-2 responses. J. Immunol 2002, 168, 5110–5116. [Google Scholar]
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Itoh, S.; Matsushita, K.; Ikeda, S.; Yamamoto, Y.; Yamauchi, Y.; Yoshioka, S.; Yamamoto, R.; Ebisu, S.; Hayashi, M.; Aubin, J.E. Bone Marrow-Derived HipOP Cell Population Is Markedly Enriched in Osteoprogenitors. Int. J. Mol. Sci. 2012, 13, 10229-10235. https://doi.org/10.3390/ijms130810229
Itoh S, Matsushita K, Ikeda S, Yamamoto Y, Yamauchi Y, Yoshioka S, Yamamoto R, Ebisu S, Hayashi M, Aubin JE. Bone Marrow-Derived HipOP Cell Population Is Markedly Enriched in Osteoprogenitors. International Journal of Molecular Sciences. 2012; 13(8):10229-10235. https://doi.org/10.3390/ijms130810229
Chicago/Turabian StyleItoh, Shousaku, Kenta Matsushita, Shun Ikeda, Yumiko Yamamoto, Yukako Yamauchi, Seisuke Yoshioka, Reiko Yamamoto, Shigeyuki Ebisu, Mikako Hayashi, and Jane E. Aubin. 2012. "Bone Marrow-Derived HipOP Cell Population Is Markedly Enriched in Osteoprogenitors" International Journal of Molecular Sciences 13, no. 8: 10229-10235. https://doi.org/10.3390/ijms130810229