High Resolution Crystal Structures of the Cerebratulus lacteus Mini-Hb in the Unligated and Carbomonoxy States


 ,
,
Abstract
:
1. Introduction
2. Results and Discussion
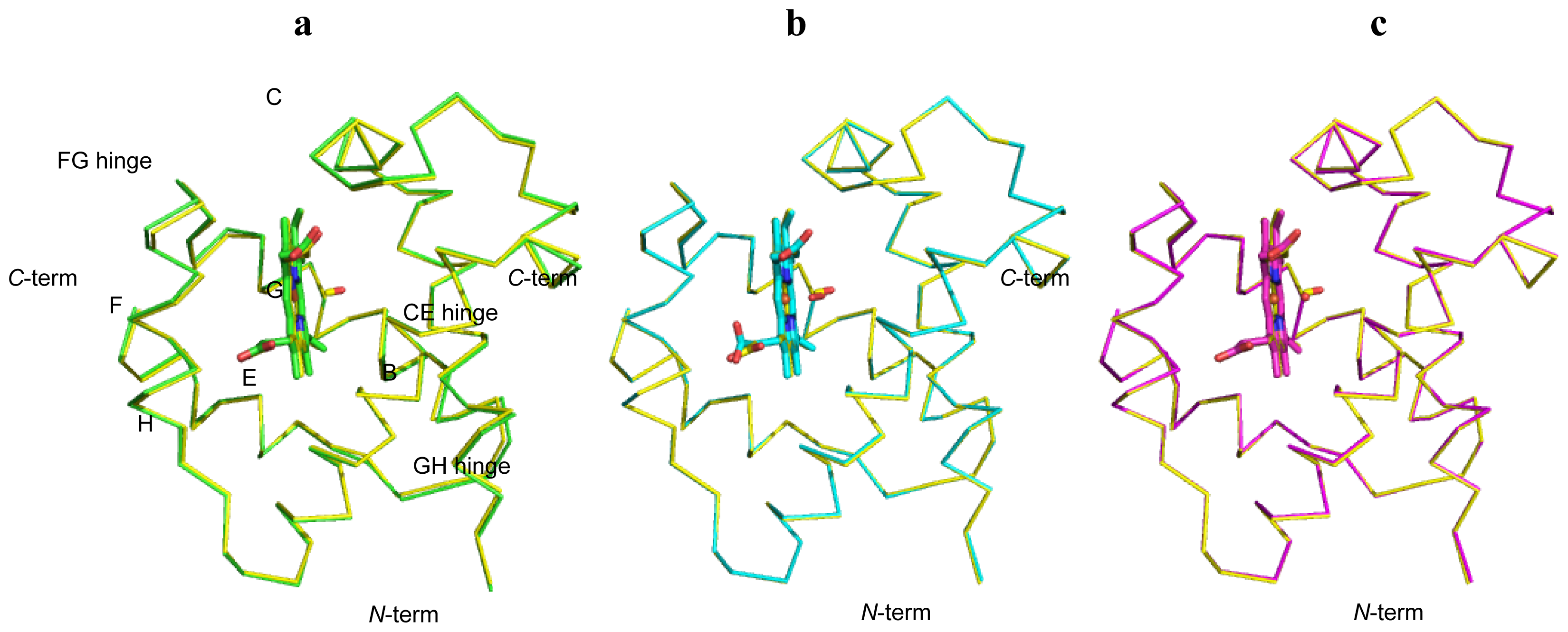
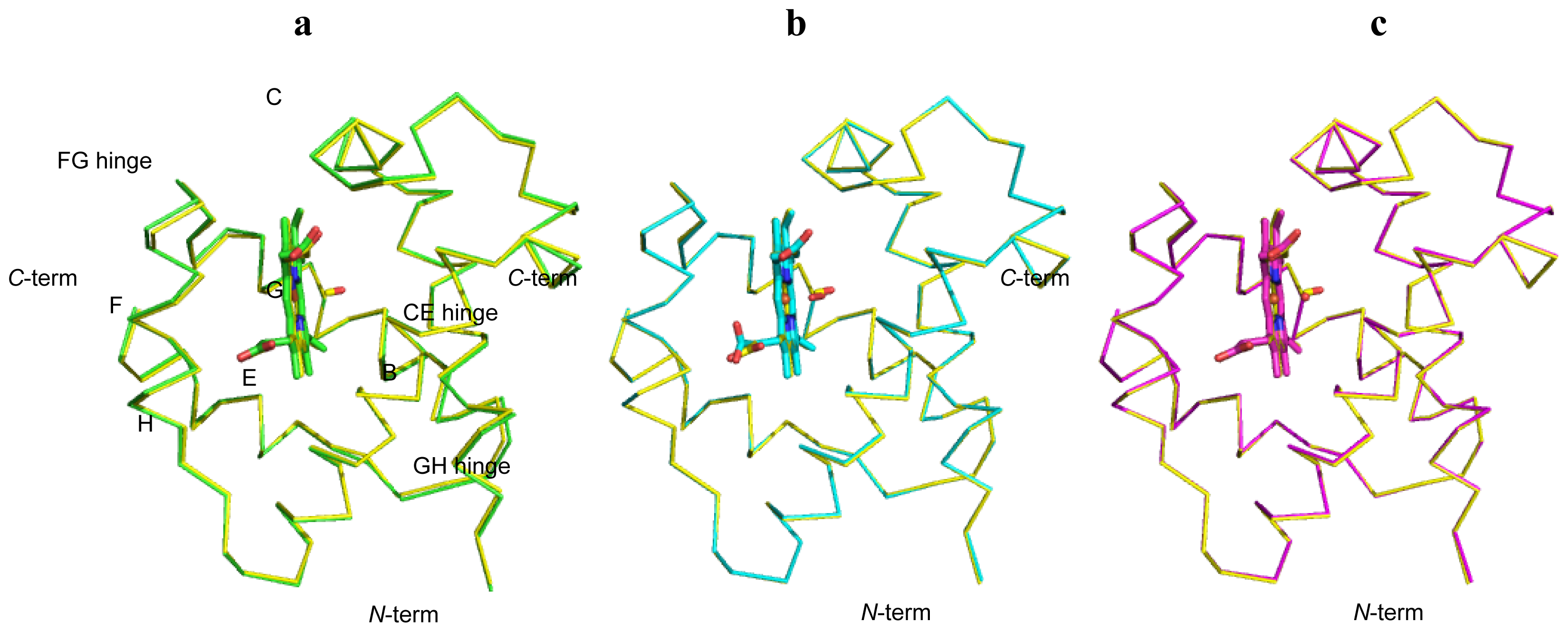
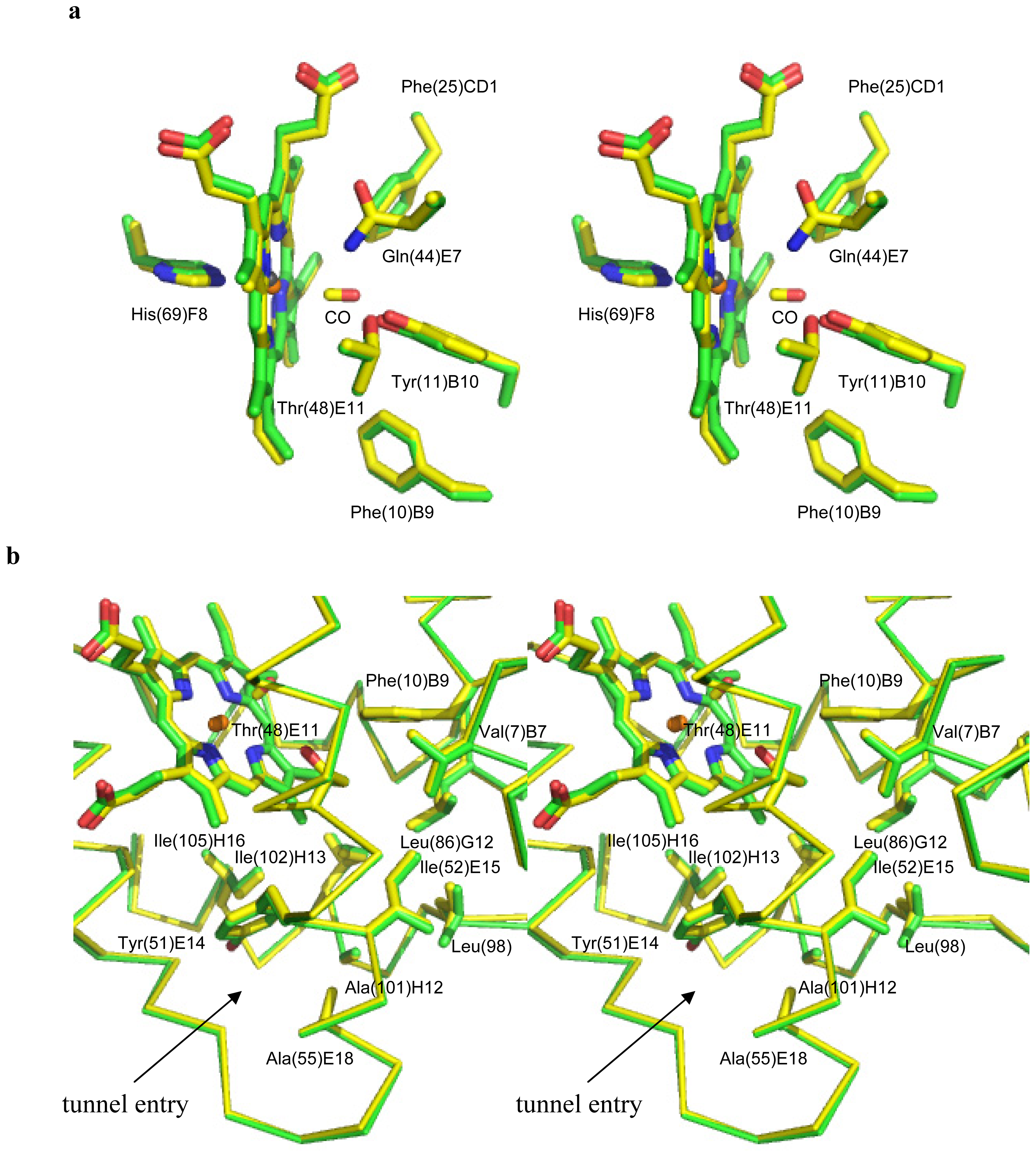
2.1. Overall Structures of CerHb in the Unligated and Carbomonoxy States
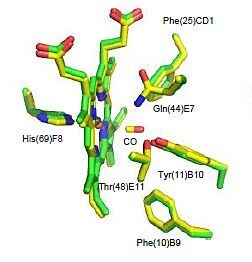
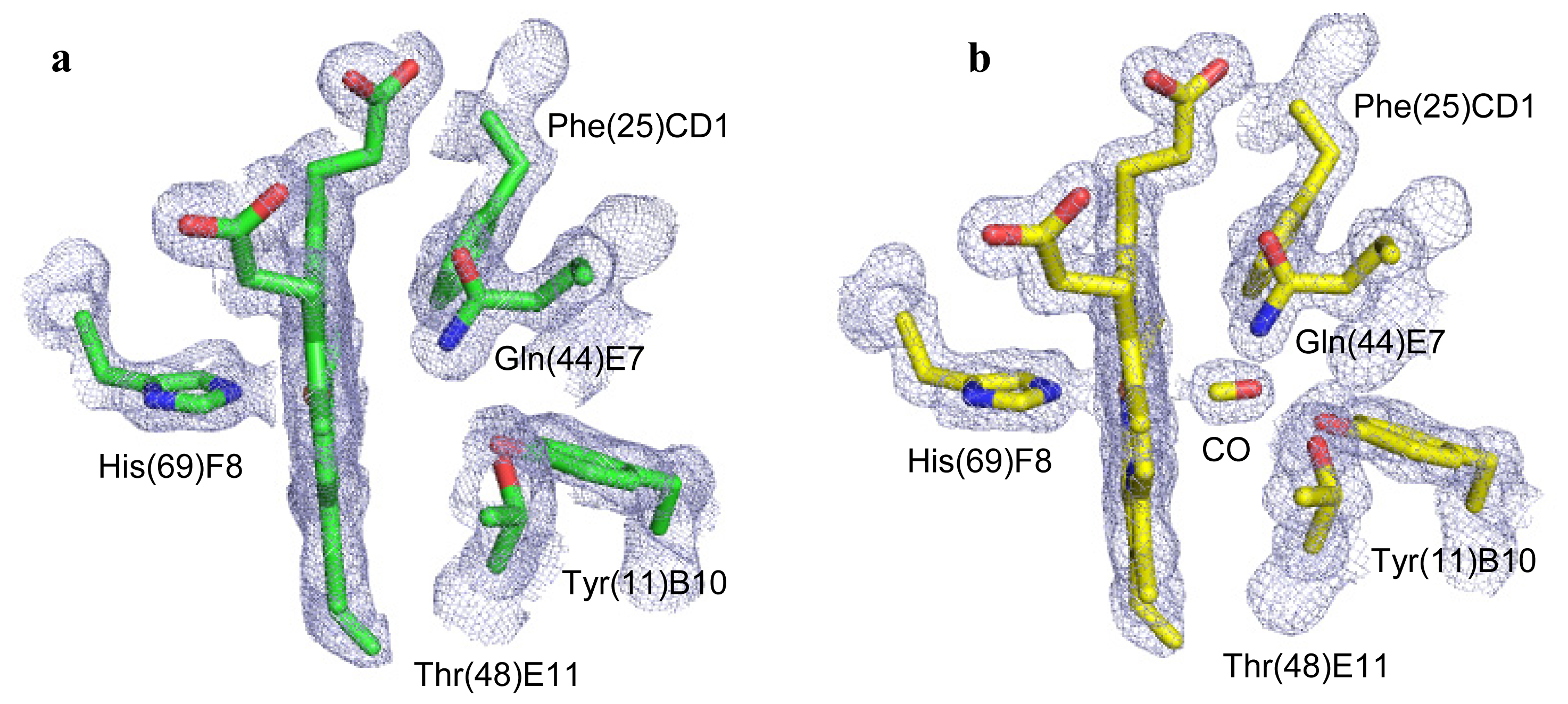
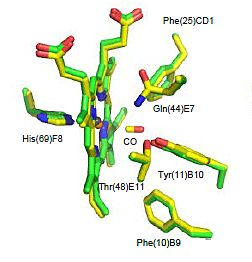
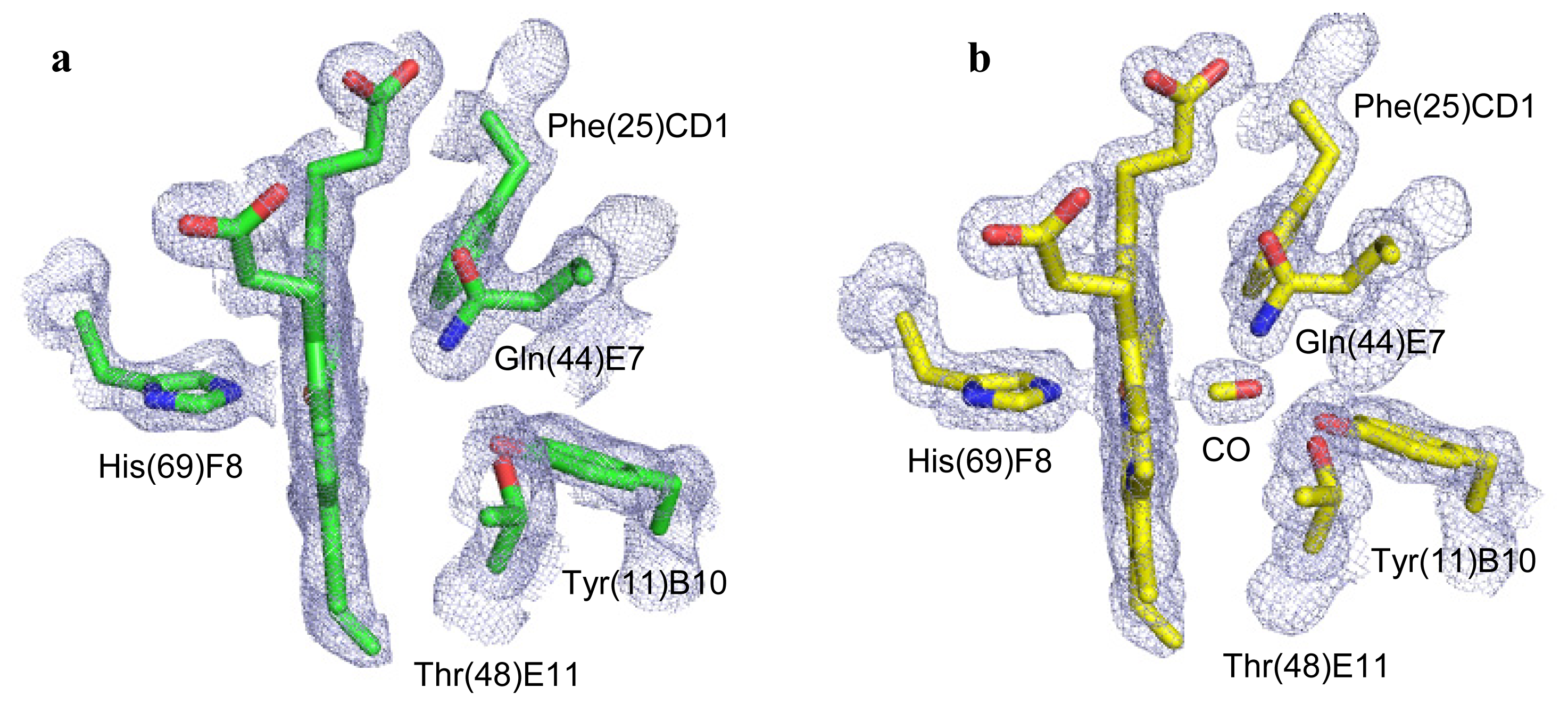
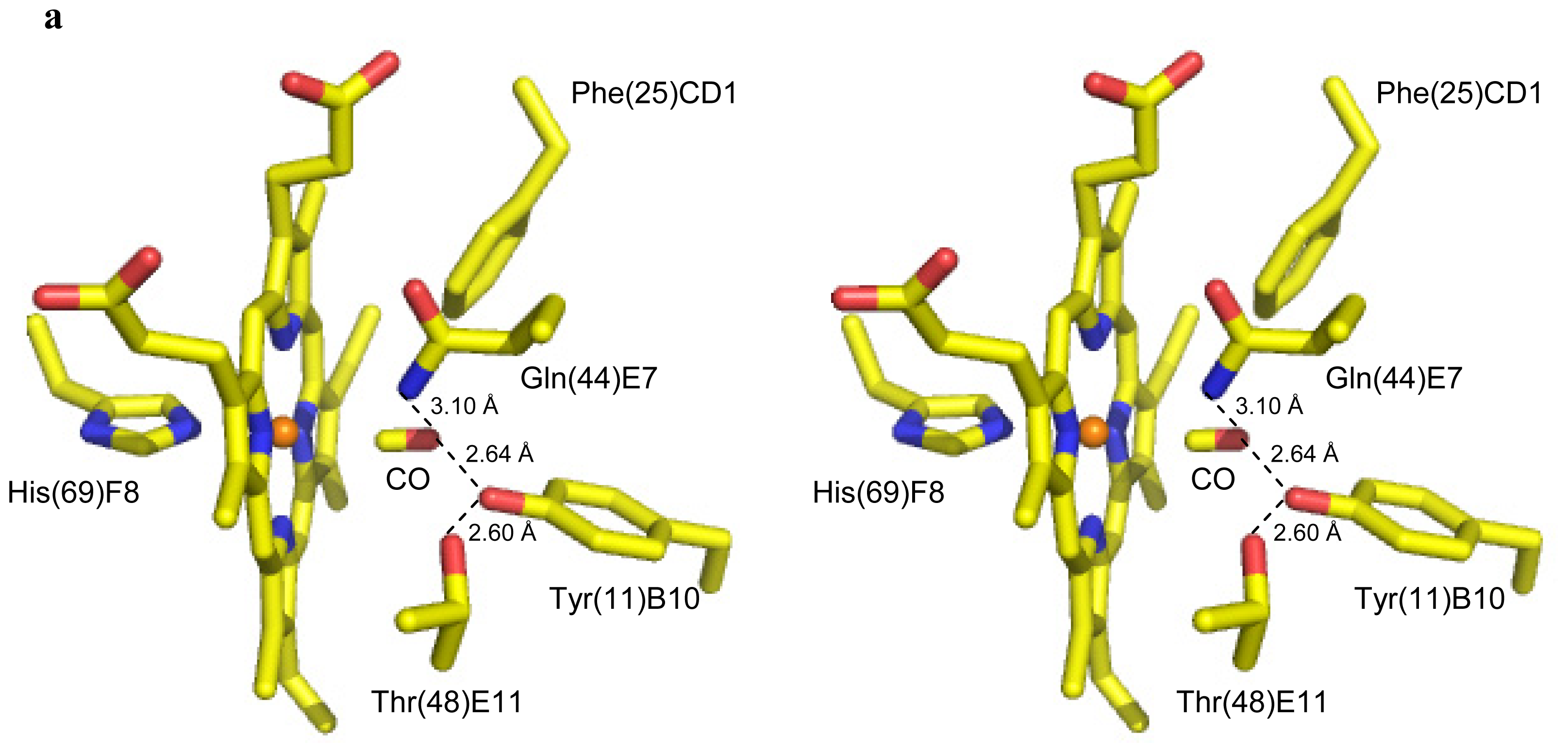
2.2. CO Binding at the CerHb Distal Site
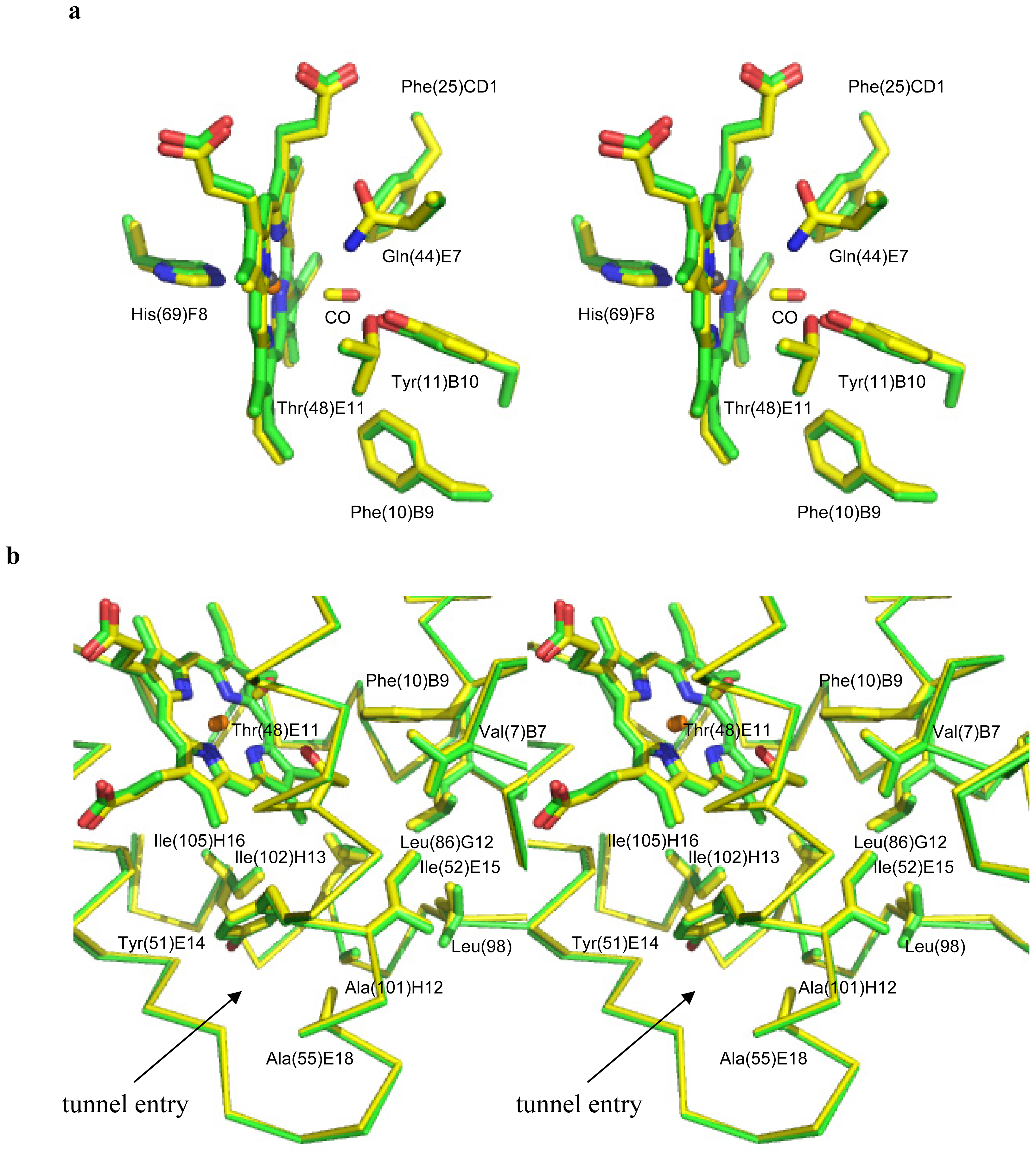
2.3. Ligand Diffusion Pathway through the CerHb Apolar Tunnel
3. Experimental Section
3.1. Crystallization of CerHb and Derivative Preparation
3.2. Data Collection, Phasing and Refinement
4. Conclusions
Acknowledgments
- Conflict of InterestThe authors declare no conflict of interest.
References
- Bolognesi, M.; Bordo, D.; Rizzi, M.; Tarricone, C.; Ascenzi, P. Nonvertebrate hemoglobins: structural bases for reactivity. Prog. Biophys. Mol. Biol 1997, 68, 29–68. [Google Scholar]
- Vandergon, T.L.; Riggs, C.K.; Gorr, T.A.; Colacino, J.M.; Riggs, A.F. The mini-hemoglobins in neural and body wall tissue of the nemertean worm, Cerebratulus lacteus. J. Biol. Chem 1998, 273, 16998–17011. [Google Scholar]
- Pesce, A.; Nardini, M.; Dewilde, S.; Geuens, E.; Yamauchi, K.; Ascenzi, P.; Riggs, A.F.; Moens, L.; Bolognesi, M. The 109 residue nerve tissue minihemoglobin from Cerebratulus lacteus highlights striking structural plasticity of the α-helical globin fold. Structure 2002, 10, 725–735. [Google Scholar]
- Pesce, A.; Nardini, M.; Ascenzi, P.; Geuens, E.; Dewilde, S.; Moens, L.; Bolognesi, M.; Riggs, A.F.; Hale, A.; Deng, P.; et al. ThrE11 regulates O2 affinity in Cerebratulus lacteus mini-hemoglobin. J. Biol. Chem 2004, 279, 33662–33672. [Google Scholar]
- Martí, M.A.; Bikiel, D.E.; Crespo, A.; Nardini, M.; Bolognesi, M.; Estrin, D.A. Two distinct heme distal site states define Cerebratulus lacteus mini-hemoglobin oxygen affinity. Proteins Struct. Funct. Bioinforma 2006, 62, 641–648. [Google Scholar]
- Draghi, F.; Miele, A.E.; Travaglini-Allocatelli, C.; Vallone, B.; Brunori, M.; Gibson, Q.H.; Olson, J.S. Controlling ligand binding in myoglobin by mutagenesis. J. Biol. Chem 2002, 277, 7509–7519. [Google Scholar]
- Salter, M.D.; Nienhaus, K.; Nienhaus, G.U.; Dewilde, S.; Moens, L.; Pesce, A.; Nardini, M.; Bolognesi, M.; Olson, J.S. The apolar channel in Cerebratulus lacteus hemoglobin is the route for O2 entry and exit. J. Biol. Chem 2008, 283, 35689–35702. [Google Scholar]
- Pesce, A.; Nardini, M.; Dewilde, S.; Capece, L.; Martí, M.A.; Congia, S.; Salter, M.D.; Blouin, G.C.; Estrin, D.A.; Ascenzi, P.; et al. Ligand migration in the apolar tunnel of Cerebratulus lacteus mini-hemoglobin. J. Biol. Chem 2011, 286, 5347–5358. [Google Scholar]
- Srajer, V.; Ren, Z.; Teng, T.Y.; Schmidt, M.; Ursby, T.; Bourgeois, D.; Pradervand, C.; Schildkamp, W.; Wulff, M.; Moffat, K. Protein conformational relaxation and ligand migration in myoglobin: a nanosecond to millisecond molecular movie from time-resolved Laue X-ray diffraction. Biochemistry 2001, 40, 13802–13815. [Google Scholar]
- Laskowski, R.A.; MacArthur, M.W.; Moss, D.S.; Thornton, J.M. PROCHECK: a program to check the stereochemical quality of protein structures. J. Appl. Crystallogr 1993, 26, 283–291. [Google Scholar]
- Kachalova, G.S.; Popov, A.N.; Bartunik, H.D. A steric mechanism for inhibition of CO binding to heme proteins. Science 1999, 284, 473–476. [Google Scholar]
- Vojtechovsky, J.; Chu, K.; Berendzen, J.; Sweet, R.M.; Schlichting, I. Crystal structures of myoglobin-ligand complexes at near-atomic resolution. Biophys. J 1999, 77, 2153–2174. [Google Scholar]
- Pesce, A.; Nardini, M.; Dewilde, S.; Ascenzi, P.; Riggs, A.F.; Yamauchi, K.; Geuens, E.; Moens, L.; Bolognesi, M. Crystallization and preliminary X-ray analysis of neural haemoglobin from the nemertean worm Cerebratulus lacteus. Acta Crystallogr. D Biol. Crystallogr 2001, 57, 1897–1899. [Google Scholar]
- Leslie, A.G. The integration of macromolecular diffraction data. Acta Crystallogr. D Biol. Crystallogr 2006, 62, 48–57. [Google Scholar]
- Evans, P. Scaling and assessment of data quality. Acta Crystallogr. D Biol. Crystallogr 2006, 62, 72–82. [Google Scholar]
- Vagin, A.; Teplyakov, A. Molecular replacement with MOLREP. Acta Crystallogr. D Biol. Crystallogr 1997, 66, 22–25. [Google Scholar]
- Murshudov, G.N.; Vagin, A.A.; Dodson, E.J. Refinement of macromolecular structures by the maximum-likelihood method. Acta Crystallogr. D Biol. Crystallogr 1997, 53, 240–255. [Google Scholar]
- Emsley, P.; Cowtan, K. Coot: model-building tools for molecular graphics. Acta Crystallogr. D Biol. Crystallogr 2004, 60, 2126–2132. [Google Scholar]
- Voss, N.R.; Gerstein, M. 3V: cavity, channel and cleft volume calculator and extractor. Nucl. Acid. Res 2010, 38, W555–W562. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| unligated-CerHb | CerHb-CO | |
|---|---|---|
| Data Collection Parameters: | ||
| Space group | P212121 | P212121 |
| Cell dimensions (Å) | a = 42.1 | a = 42.7 |
| b = 44.3 | b = 43.5 | |
| c = 62.1 | c = 60.1 | |
| Resolution (Å) | 25.4–1.9 (2.00–1.90) | 30.5–1.5 (1.58–1.50) |
| Observations | 55,282 | 129,825 |
| Unique reflections | 9,482 | 18,465 |
| Completeness (%) | 98.6 (100) a | 99.6 (100.0) |
| R-merge b (%) | 20.0 (35.7) | 8.2 (31.9) |
| I/σ(I) | 6.0 (4.2) | 13.4 (5.7) |
| Multiplicity | 5.8 (6.4) | 7.0 (7.3) |
| Refinement: | ||
| R-factor c/R-free (%) | 18.3/22.1 | 14.6/17.9 |
| Protein atoms in the a.u. d | 819 | 836 |
| Heme prostetic group | 1 | 1 |
| Carbon monoxide molecule | - | 1 |
| Water molecules | 115 | 123 |
| Sulfate ions | 2 | 2 |
| Acetate ions | 1 | 1 |
| Glycerol | 1 | 1 |
| Model quality: | ||
| Overall B-factor (Å2): | ||
| Protein and heme | 13.9 | 13.3 |
| Carbon monoxide molecule | - | 18.9 |
| Ions and glycerol | 37.9 | 23.8 |
| Water molecules | 25.8 | 27.1 |
| Rmsd from ideal values: | ||
| bond lengths (Å) | 0.014 | 0.010 |
| bond angles (°) | 1.3 | 1.3 |
| Ramachandran plot (%) e: | ||
| most favored regions | 96.8 | 96.8 |
| additional allowed regions | 3.2 | 3.2 |
| unligated-CerHb | CerHb-CO | CerHb-O2 | |
|---|---|---|---|
| Fe–CO (Å) | 2.09 | ||
| Fe–O2 (Å) | 1.94 | ||
| Fe–C–O angle (°) | 172 | ||
| Fe–O1–O2 angle (°) | 103 | ||
| Tyr(11)B10 OH---O (Å) | 2.64 | ||
| Tyr(11)B10 OH---O2 (Å) | 2.57 | ||
| Gln(44)E7 NE2---O (Å) | 3.10 | ||
| Gln(44)E7 NE2---O1 (Å) | 2.60 | ||
| Gln(44)E7 NE2---O2 (Å) | 3.34 | ||
| Tyr(11)B10 OH---Thr(48)E11 OG1 (Å) | 2.76 | 2.60 | 2.59 |
| Tyr(11)B10 OH---Gln(44)E7 NE2 (Å) | 3.13 | 3.06 | 3.24 |
| Gln(44)E7 NE2---Thr(48)E11 OG1 (Å) | 3.70 | 3.56 | 3.53 |
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Germani, F.; Pesce, A.; Venturini, A.; Moens, L.; Bolognesi, M.; Dewilde, S.; Nardini, M. High Resolution Crystal Structures of the Cerebratulus lacteus Mini-Hb in the Unligated and Carbomonoxy States. Int. J. Mol. Sci. 2012, 13, 8025-8037. https://doi.org/10.3390/ijms13078025
Germani F, Pesce A, Venturini A, Moens L, Bolognesi M, Dewilde S, Nardini M. High Resolution Crystal Structures of the Cerebratulus lacteus Mini-Hb in the Unligated and Carbomonoxy States. International Journal of Molecular Sciences. 2012; 13(7):8025-8037. https://doi.org/10.3390/ijms13078025
Chicago/Turabian StyleGermani, Francesca, Alessandra Pesce, Andrea Venturini, Luc Moens, Martino Bolognesi, Sylvia Dewilde, and Marco Nardini. 2012. "High Resolution Crystal Structures of the Cerebratulus lacteus Mini-Hb in the Unligated and Carbomonoxy States" International Journal of Molecular Sciences 13, no. 7: 8025-8037. https://doi.org/10.3390/ijms13078025