O6-Methylguanine-Methyltransferase (MGMT) Promoter Methylation Status in Glioma Stem-Like Cells is Correlated to Temozolomide Sensitivity Under Differentiation-Promoting Conditions

Abstract
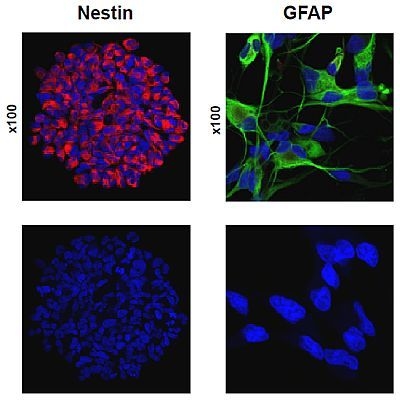
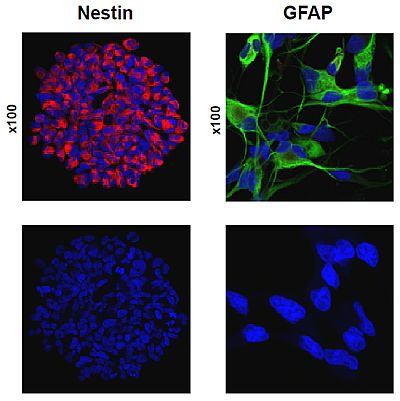
:
1. Introduction
2. Results and Discussion
2.1. Primary Culture of Glioma Stem-Like Cells in Undifferentiated and Differentiated States
2.2. Temozolomide Sensitivity
2.3. MGMT Promoter Methylation Status in GBM Tumors, Neurospheres and Differentiated Cells
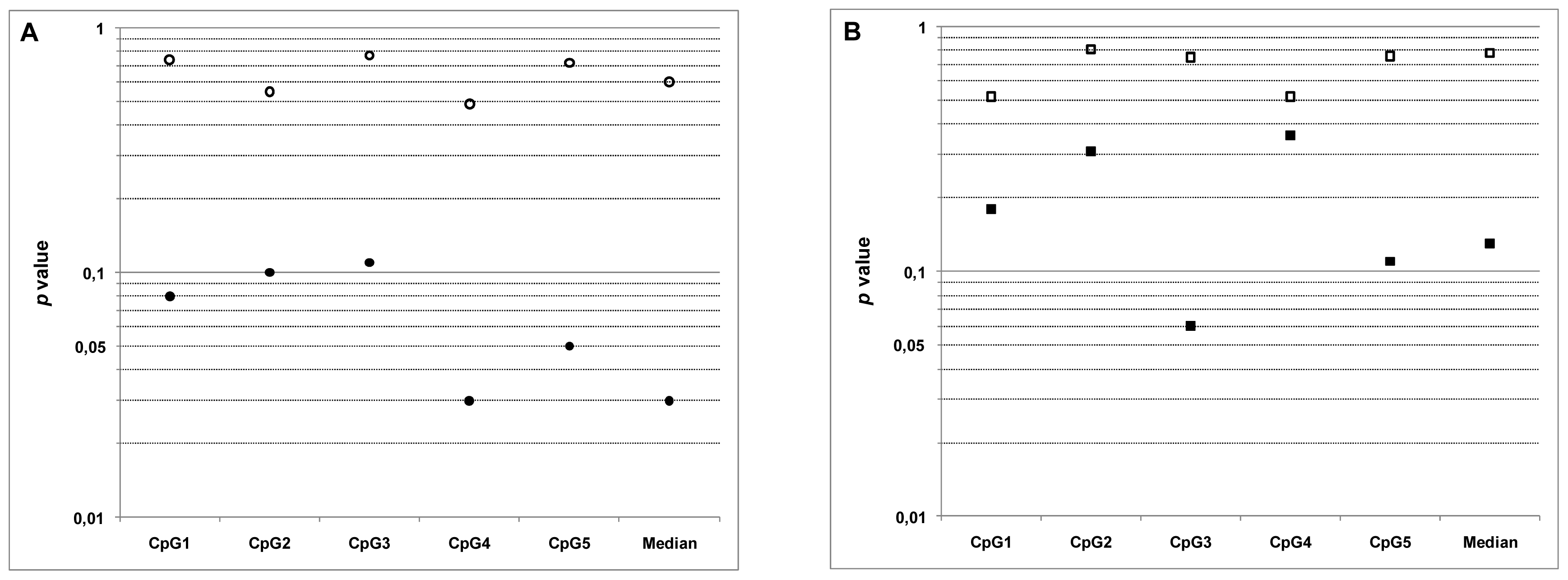
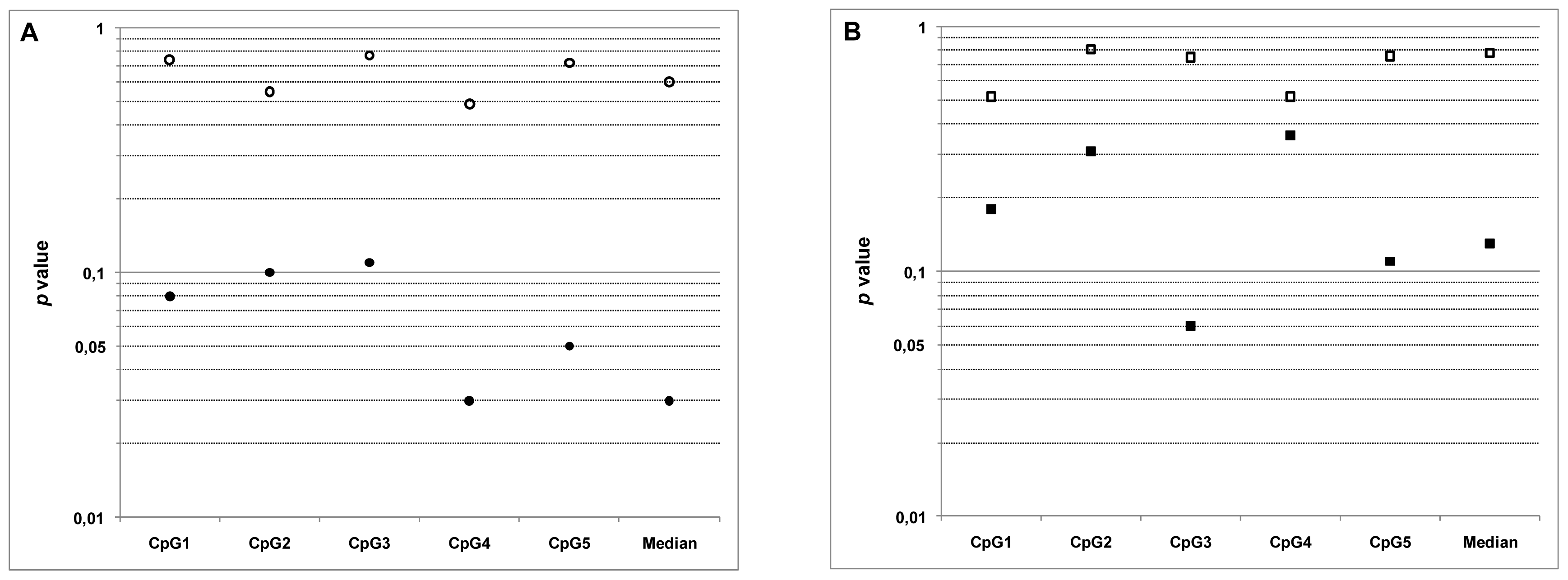
2.4. Relationship between MGMT Promoter Methylation and Sensitivity to Temozolomide
3. Experimental Section
3.1. GBM Samples
3.2. RT-qPCR
3.3. Proliferation and Cytotoxicity Assays
3.4. DNA Extraction and Bisulfitation
3.5. Quantification of Methylation by Pyrosequencing
3.6. Statistical Analysis
4. Conclusions
Supplementary Materials
ijms-13-06983-s001.pdfAcknowledgments
- Conflict of InterestThe authors declare no conflicts of interest related to this work.
References
- Gerson, S.L. MGMT: Its role in cancer aetiology and cancer therapeutics. Nat. Rev. Cancer 2004, 4, 296–307. [Google Scholar]
- Drablos, F.; Feyzi, E.; Aas, P.A.; Vaagbø, C.B.; Kavli, B.; Bratlie, M.S.; Peña-Diaz, J.; Otterlei, M.; Slupphaug, G.; Krokan, H.E. Alkylation damage in DNA and RNA repair mechanisms and medical significance. DNA Repair 2004, 3, 1389–1407. [Google Scholar]
- Kaina, B.; Christmann, M.; Naumann, S.; Roos, W.P. MGMT: Key node in the battle against genotoxicity, carcinogenicity and apoptosis induced by alkylating agents. DNA Repair 2007, 6, 1079–1099. [Google Scholar]
- Hegi, M.E.; Diserens, A.C.; Gorlia, T.; Hamou, M.F.; de Tribolet, N.; Weller, M.; Kros, J.M.; Hainfellner, J.A.; Mason, W.; Mariani, L.; et al. MGMT gene silencing and benefit from temozolomide in glioblastoma. N. Engl. J. Med 2005, 352, 997–1003. [Google Scholar]
- Chinot, O.L.; Barrié, M.; Fuentes, S.; Eudes, N.; Lancelot, S.; Metellus, P.; Muracciole, X.; Braguer, D.; Ouafik, L.; Martin, P.M.; et al. Correlation between O6-methylguanine-DNA methyltransferase and survival in inoperable newly diagnosed glioblastoma patients treated with neoadjuvant temozolomide. J. Clin. Oncol 2007, 25, 1470–1475. [Google Scholar]
- Martinez, R.; Martin-Subero, J.I.; Rohde, V.; Kirsch, M.; Alaminos, M.; Fernandez, A.F.; Ropero, S.; Schackert, G.; Esteller, M.; et al. A microarray-based DNA methylation study of glioblastoma multiforme. Epigenetics 2009, 4, 255–264. [Google Scholar]
- van den Bent, M.J.; Dubbink, H.J.; Sanson, M.; van der Lee-Haarloo, C.R.; Hegi, M.; Jeuken, J.W.; Ibdaih, A.; Brandes, A.A.; Taphoorn, M.J.; Frenay, M.; et al. MGMT promoter methylation is prognostic but not predictive for outcome to adjuvant PCV chemotherapy in anaplastic oligodendroglial tumors: A report from EORTC Brain Tumor Group Study 26951. J. Clin. Oncol 2009, 27, 5881–5886. [Google Scholar]
- Gorlia, T.; van den Bent, M.J.; Hegi, M.E.; Mirimanoff, R.O.; Weller, M.; Cairncross, J.G.; Eisenhauer, E.; Belanger, K.; Brandes, A.A.; Allgeier, A.; et al. Nomograms for predicting survival of patients with newly diagnosed glioblastoma: Prognostic factor analysis of EORTC and NCIC trial 26981-22981/CE.3. Lancet Oncol 2007, 9, 29–38. [Google Scholar]
- European Organisation for Research and Treatment of Cancer Brain Tumour and Radiation Oncology Groups; the National Cancer Institute of Canada Clinical Trials Group. Effects of radiotherapy with concomitant and adjuvant temozolomide versus radiotherapy alone on survival in glioblastoma in a randomised phase III study: 5-year analysis of the EORTC-NCIC trial. Lancet Oncol. 2009, 10, 459–466.
- Blanc, J.L.; Wager, M.; Guilhot, J.; Kusy, S.; Bataille, B.; Chantereau, T.; Lapierre, F.; Larsen, C.J.; Karayan-Tapon, L. Correlation of clinical features and methylation status of MGMT gene promoter in glioblastomas. J. Neurooncol 2004, 68, 275–283. [Google Scholar]
- Kamiryo, T.; Tada, K.; Shiraishi, S.; Shinojima, N.; Kochi, M.; Ushio, Y. Correlation between promoter hypermethylation of the O6-methylguanine-deoxyribonucleic acid methyltransferase gene and prognosis in patients with high-grade astrocytic tumors treated with surgery, radiotherapy, and 1-(4-amino-2-methyl-5-pyrimidinyl)methyl-3-(2-chloroethyl)-3-nitrosourea-based chemotherapy. Neurosurgery 2004, 54, 349–357. [Google Scholar]
- Eramo, A.; Ricci-Vitiani, L.; Zeuner, A.; Pallini, R.; Lotti, F.; Sette, G.; Pilozzi, E.; Larocca, L.M.; Peschle, C.; de Maria, R. Chemotherapy resistance of glioblastoma stem cells. Cell Death Differ 2006, 13, 1238–1241. [Google Scholar]
- Liu, G.; Yuan, X.; Zeng, Z.; Tunici, P.; Ng, H.; Abdulkadir, I.R.; Lu, L.; Irvin, D.; Black, K.L.; Yu, J.S. Analysis of gene expression and chemoresistance of CD133+ cancer stem cells in glioblastoma. Mol. Cancer 2006, 5. [Google Scholar] [CrossRef]
- Pistollato, F.; Abbadi, S.; Rampazzo, E.; Persano, L.; Della Puppa, A.; Frasson, C.; Sarto, E.; Scienza, R.; D’avella, D.; Basso, G. Intratumoral hypoxic gradient drives stem cells distribution and MGMT expression in glioblastoma. Stem Cells 2010, 28, 851–862. [Google Scholar]
- Murat, A.; Migliavacca, E.; Gorlia, T.; Lambiv, W.L.; Shay, T.; Hamou, M.F.; de Tribolet, N.; Regli, L.; Wick, W.; Kouwenhoven, M.C.; et al. Stem cell-related “self-renewal” signature and high epidermal growth factor receptor expression associated with resistance to concomitant chemoradiotherapy in glioblastoma. J. Clin. Oncol 2008, 26, 3015–3024. [Google Scholar]
- Piccirillo, S.G.; Reynolds, B.A.; Zanetti, N.; Lamorte, G.; Binda, E.; Broggi, G.; Brem, H.; Olivi, A.; Dimeco, F.; Vescovi, A.L. Bone morphogenetic proteins inhibit the tumorigenic potential of human brain tumour-initiating cells. Nature 2006, 444, 761–765. [Google Scholar]
- Campos, B.; Wan, F.; Farhadi, M.; Ernst, A.; Zeppernick, F.; Tagscherer, K.E.; Ahmadi, R.; Lohr, J.; Dictus, C.; Gdynia, G.; et al. Differentiation therapy exerts antitumor effects on stem-like glioma cells. Clin. Cancer Res 2010, 16, 2715–2728. [Google Scholar]
- Wakimoto, H.; Kesari, S.; Farrell, C.J.; Curry, W.T., Jr; Zaupa, C.; Aghi, M.; Kuroda, T.; Stemmer-Rachamimov, A.; Shah, K.; Liu, T.C.; et al. Human glioblastoma-derived cancer stem cells: Establishment of invasive glioma models and treatment with oncolytic herpes simplex virus vectors. Cancer Res. 2009, 69, 3472–3481. [Google Scholar]
- Blough, M.D.; Westgate, M.R.; Beauchamp, D.; Kelly, J.J.; Stechishin, O.; Ramirez, A.L.; Weiss, S.; Cairncross, J.G. Sensitivity to temozolomide in brain tumor initiating cells. Neuro-Oncology 2010, 12, 756–760. [Google Scholar]
- Singh, S.K.; Hawkins, C.; Clarke, I.D.; Squire, J.A.; Bayani, J.; Hide, T.; Henkelman, R.M.; Cusimano, M.D.; Dirks, P.B. Identification of human brain tumour initiating cells. Nature 2004, 432, 396–401. [Google Scholar]
- Galli, R.; Binda, E.; Orfanelli, U.; Cipelletti, B.; Gritti, A.; de Vitis, S.; Fiocco, R.; Foroni, C.; Dimeco, F.; Vescovi, A. Isolation and characterization of tumorigenic, stem-like neural precursors from human glioblastoma. Cancer Res 2004, 64, 7011–7021. [Google Scholar]
- Ogden, A.T.; Waziri, A.E.; Lochhead, R.A.; Fusco, D.; Lopez, K.; Ellis, J.A.; Kang, J.; Assanah, M.; McKhann, G.M.; Sisti, M.B.; et al. Identification of A2B5+CD133− tumor-initiating cells in adult human gliomas. Neurosurgery 2008, 62, 505–515. [Google Scholar]
- Chen, R.; Nishimura, M.C.; Bumbaca, S.M.; Kharbanda, S.; Forrest, W.F.; Kasman, I.M.; Greve, J.M.; Soriano, R.H.; Gilmour, L.L.; Rivers, C.S.; et al. A hierarchy of self-renewing tumor-initiating cell types in glioblastoma. Cancer Cell 2010, 17, 362–375. [Google Scholar]
- Villalva, C.; Martin-Lannerée, S.; Cortes, U.; Dkhissi, F.; Wager, M.; Le Corf, A.; Tourani, J.M.; Dusanter-Fourt, I.; Turhan, A.G.; Karayan-Tapon, L. STAT3 is essential for the maintenance of neurosphere-initiating tumor cells in patients with glioblastomas: A potential for targeted therapy? Int. J. Cancer 2011, 128, 826–838. [Google Scholar]
- Beier, D.; Röhrl, S.; Pillai, D.R.; Schwarz, S.; Kunz-Schughart, L.A.; Leukel, P.; Proescholdt, M.; Brawanski, A.; Bogdahn, U.; Trampe-Kieslich, A.; et al. Temozolomide preferentially depletes cancer stem cells in glioblastoma. Cancer Res 2008, 68, 5706–5715. [Google Scholar]
- Karayan-Tapon, L.; Quillien, V.; Guilhot, J.; Wager, M.; Fromont, G.; Saikali, S.; Etcheverry, A.; Hamlat, A.; Loussouarn, D.; Campion, L.; et al. Prognostic value of O6-methylguanine-DNA methyltransferase status in glioblastoma patients, assessed by five different methods. J. Neuro-Oncol 2009, 97, 311–322. [Google Scholar]
- Yoshino, A.; Ogino, A.; Yachi, K.; Ohta, T.; Fukushima, T.; Watanabe, T.; Katayama, Y.; Okamoto, Y.; Naruse, N.; Sano, E.; et al. Gene expression profiling predicts response to temozolomide in malignant gliomas. Int. J. Oncol 2010, 36, 1367–1377. [Google Scholar]
- Burger, P.C.; Kleihues, P. Cytologic composition of the untreated glioblastoma with implications for evaluation of needle biopsies. Cancer 1989, 63, 2014–2023. [Google Scholar]
- Costa, B.M.; Caeiro, C.; Guimarães, I.; Martinho, O.; Jaraquemada, T.; Augusto, I.; Castro, L.; Osório, L.; Linhares, P.; Honavar, M.; et al. Prognostic value of MGMT promoter methylation in glioblastoma patients treated with temozolomide-based chemoradiation: A Portuguese multicentre study. Oncol. Rep 2010, 23, 1655–1662. [Google Scholar]
- European Organisation for Research and Treatment of Cancer Brain Tumor and Radiotherapy Groups; the National Cancer Institute of Canada Clinical Trials Group. Radiotherapy plus concomitant and adjuvant temozolomide for glioblastoma. N. Engl. J. Med. 2005, 352, 987–996.

{kind=link}
{kind=link}
| Patient # | Sex | Age (Years) | Tumor location | OMS classification | Overall survival (Months) | Cell line | Index of differentitaion |
|---|---|---|---|---|---|---|---|
| 1 | M | 69 | Left fronto-parietal | Grade IV | 15 | GBM1 | 83.5 |
| 2 | M | 57 | Right fronto-temporal | Grade IV | 10 | GBM2 | 31.6 |
| 3 | M | 53 | Right parieto-occipital | Grade IV | 10 | GBM3 | 6.75 |
| 4 | M | 51 | Left frontal | Grade IV | 13 | GBM4 | 34.3 |
| 5 | M | 56 | Left temporo-frontal | Grade IV | 6 | GBM5 | 2.26 |
| 6 | M | 69 | Right basi-frontal | Grade IV | More than 18 | GBM6 | 382 |
| 7 | M | 59 | Left temporal | Grade IV | 10 | GBM7 | 12.4 |
| 8 | F | 65 | Right frontal | Grade IV | 13 | GBM8 | 4.52 |
| 9 | M | 61 | Right temporal | Grade IV | More than 18 | GBM9 | 7.71 |
| 10 | M | 56 | Left parieto-occipital | Grade IV | 10 | GBM10 | 19.3 |
| Cell line | Neurospheres | Differentiated cells | ||||
|---|---|---|---|---|---|---|
| IC10 (μM) | IC20 (μM) | IC50 (μM) | IC10 (μM) | IC20 (μM) | IC50 (μM) | |
| GBM1 | 69 | 194 | 516 | 121 | 267 | 493 |
| GBM2 | 84 | 194 | 474 | 259 | 503 | 1236 |
| GBM3 | 185 | 346 | 830 | 128 | 239 | 570 |
| GBM4 | 133 | 277 | 707 | 60 | 174 | 515 |
| GBM5 | 50 | 134 | 367 | 74 | 180 | 354 |
| GBM6 | 401 | 908 | 1480 | 94 | 212 | 567 |
| GBM7 | 186 | 311 | 686 | 110 | 227 | 577 |
| GBM8 | 181 | 280 | 574 | 283 | 557 | 784 |
| GBM9 | 65 | 185 | 495 | 80 | 190 | 413 |
| GBM10 | 202 | 349 | 787 | 894 | 1770 | 3950 |
| Median | 157 | 278.5 | 630 | 115.5 | 233 | 568.5 |
| Cell Line | CpG1 | CpG2 | CpG3 | CpG4 | CpG5 | Median CpG | NS-IC50 | DC-IC50 | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| NS | DC | Tumor | NS | DC | Tumor | NS | DC | Tumor | NS | DC | Tumor | NS | DC | Tumor | NS | DC | Tumor | |||
| GBM1 | 3 | 14 | 1 | 7 | 33 | 2 | 4 | 20 | 2 | 5 | 31 | 1 | 3 | 13 | 2 | 4 | 20 | 2 | 516 | 493 |
| GBM2 | 1 | 1 | 1 | 2 | 3 | 3 | 2 | 2 | 2 | 1 | 2 | 2 | 2 | 1 | 2 | 2 | 2 | 2 | 474 | 1236 |
| GBM3 | 16 | 11 | 7 | 42 | 28 | 14 | 41 | 22 | 11 | 31 | 18 | 11 | 41 | 18 | 12 | 41 | 18 | 11 | 830 | 570 |
| GBM4 | 1 | 1 | 2 | 3 | 2 | 4 | 2 | 1 | 2 | 3 | 3 | 4 | 2 | 2 | 2 | 2 | 2 | 2 | 707 | 515 |
| GBM5 | 72 | 76 | 13 | 85 | 88 | 54 | 37 | 56 | 13 | 29 | 52 | 15 | 33 | 49 | 24 | 37 | 56 | 15 | 367 | 354 |
| GBM6 | 97 | 96 | 53 | 98 | 97 | 52 | 56 | 84 | 12 | 95 | 99 | 35 | 88 | 94 | 47 | 95 | 96 | 47 | 1480 | 567 |
| GBM7 | 1 | 1 | 1 | 2 | 2 | 2 | 1 | 2 | 2 | 2 | 8 | 2 | 1 | 1 | 2 | 1 | 2 | 2 | 686 | 577 |
| GBM8 | 1 | 1 | 1 | 3 | 2 | 3 | 2 | 2 | 2 | 3 | 10 | 3 | 1 | 2 | 1 | 2 | 2 | 2 | 574 | 784 |
| GBM9 | 94 | 95 | 10 | 97 | 98 | 29 | 87 | 89 | 28 | 99 | 98 | 35 | 94 | 94 | 25 | 94 | 95 | 28 | 495 | 413 |
| GBM10 | 2 | 2 | 2 | 13 | 4 | 5 | 3 | 2 | 2 | 14 | 3 | 7 | 3 | 2 | 2 | 3 | 2 | 2 | 787 | 3950 |
| Median | 2.5 | 6.5 | 2 | 10 | 16 | 4.5 | 3.5 | 11 | 2 | 9.5 | 14 | 5.5 | 3 | 7.5 | 2 | 3.5 | 10 | 2 | 630 | 568.5 |
| CpG1 * | CpG2 * | CpG3 * | CpG4 * | CpG5 * | Median CpG # | |
|---|---|---|---|---|---|---|
| Meth NS/Meth DC | p < 0.0001 | p = 0.0007 | p = 0.0003 | p = 0.004 | p = 0.0003 | p = 0.0003 |
| r = 0.99 | r = 0.88 | r = 0.91 | r = 0.81 | r = 0.90 | r = 0.90 | |
| Meth Tum/Meth NS | p = 0.002 | p = 0.002 | p = 0.002 | p = 0.002 | p = 0.0004 | p = 0.0007 |
| r = 0.85 | r = 0.85 | r = 0.86 | r = 0.86 | r = 0.90 | r = 0.88 | |
| Meth Tum/Meth DC | p = 0.007 | p = 0.03 | p = 0.0004 | p = 0.05 | p = 0.003 | p = 0.0005 |
| r = 0.79 | r = 0.68 | r = 0.90 | r = 0.62 | r = 0.83 | r = 0.90 | |
| CpG1 * | CpG2 * | CpG3 * | CpG4 * | CpG5 * | Median CpG # | |
|---|---|---|---|---|---|---|
| IC50 NS/Meth NS | p = 0.74 | p = 0.55 | p = 0.77 | p = 0.49 | p = 0.72 | p = 0.60 |
| r = 0.12 | r = 0.21 | r = 0.10 | r = 0.25 | r = 0.13 | r = 0.19 | |
| IC50 DS/Meth DS | p = 0.08 | p = 0.10 | p = 0.11 | p = 0.03 | p = 0.05 | p = 0.03 |
| r = −0.58 | r = −0.55 | r = −0.54 | r = −0.67 | r = −0.62 | r = −0.68 | |
| IC50 NS/Meth Tum | p = 0.52 | p = 0.81 | p = 0.75 | p = 0.52 | p = 0.76 | p = 0.78 |
| r = 0.23 | r = 0.09 | r = −0.12 | r = 0.23 | r = 0.10 | r = 0.10 | |
| IC50 DS/Meth Tum | p = 0.18 | p = 0.31 | p = 0.06 | p = 0.36 | p = 0.11 | p = 0.13 |
| r = −0.46 | r = −0.36 | r = −0.61 | r = −0.32 | r = −0.54 | r = −0.51 | |
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Villalva, C.; Cortes, U.; Wager, M.; Tourani, J.-M.; Rivet, P.; Marquant, C.; Martin, S.; Turhan, A.G.; Karayan-Tapon, L. O6-Methylguanine-Methyltransferase (MGMT) Promoter Methylation Status in Glioma Stem-Like Cells is Correlated to Temozolomide Sensitivity Under Differentiation-Promoting Conditions. Int. J. Mol. Sci. 2012, 13, 6983-6994. https://doi.org/10.3390/ijms13066983
Villalva C, Cortes U, Wager M, Tourani J-M, Rivet P, Marquant C, Martin S, Turhan AG, Karayan-Tapon L. O6-Methylguanine-Methyltransferase (MGMT) Promoter Methylation Status in Glioma Stem-Like Cells is Correlated to Temozolomide Sensitivity Under Differentiation-Promoting Conditions. International Journal of Molecular Sciences. 2012; 13(6):6983-6994. https://doi.org/10.3390/ijms13066983
Chicago/Turabian StyleVillalva, Claire, Ulrich Cortes, Michel Wager, Jean-Marc Tourani, Pierre Rivet, Celine Marquant, Sebastien Martin, Ali G. Turhan, and Lucie Karayan-Tapon. 2012. "O6-Methylguanine-Methyltransferase (MGMT) Promoter Methylation Status in Glioma Stem-Like Cells is Correlated to Temozolomide Sensitivity Under Differentiation-Promoting Conditions" International Journal of Molecular Sciences 13, no. 6: 6983-6994. https://doi.org/10.3390/ijms13066983