Fragment C of Tetanus Toxin: New Insights into Its Neuronal Signaling Pathway

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
- direct inhibition of motoneurons by impulses in Group I a afferent fibers of antagonistic muscles;
- inhibition by impulses in the Group I b afferent fibers from muscles of the same limb;
- inhibition of extensor motoneurons by impulses in Groups II and III muscle afferent fibers and in cutaneous afferent fibers; and
- inhibition of motoneurons following the activation of Renshaw cells by volleys in axon collaterals.
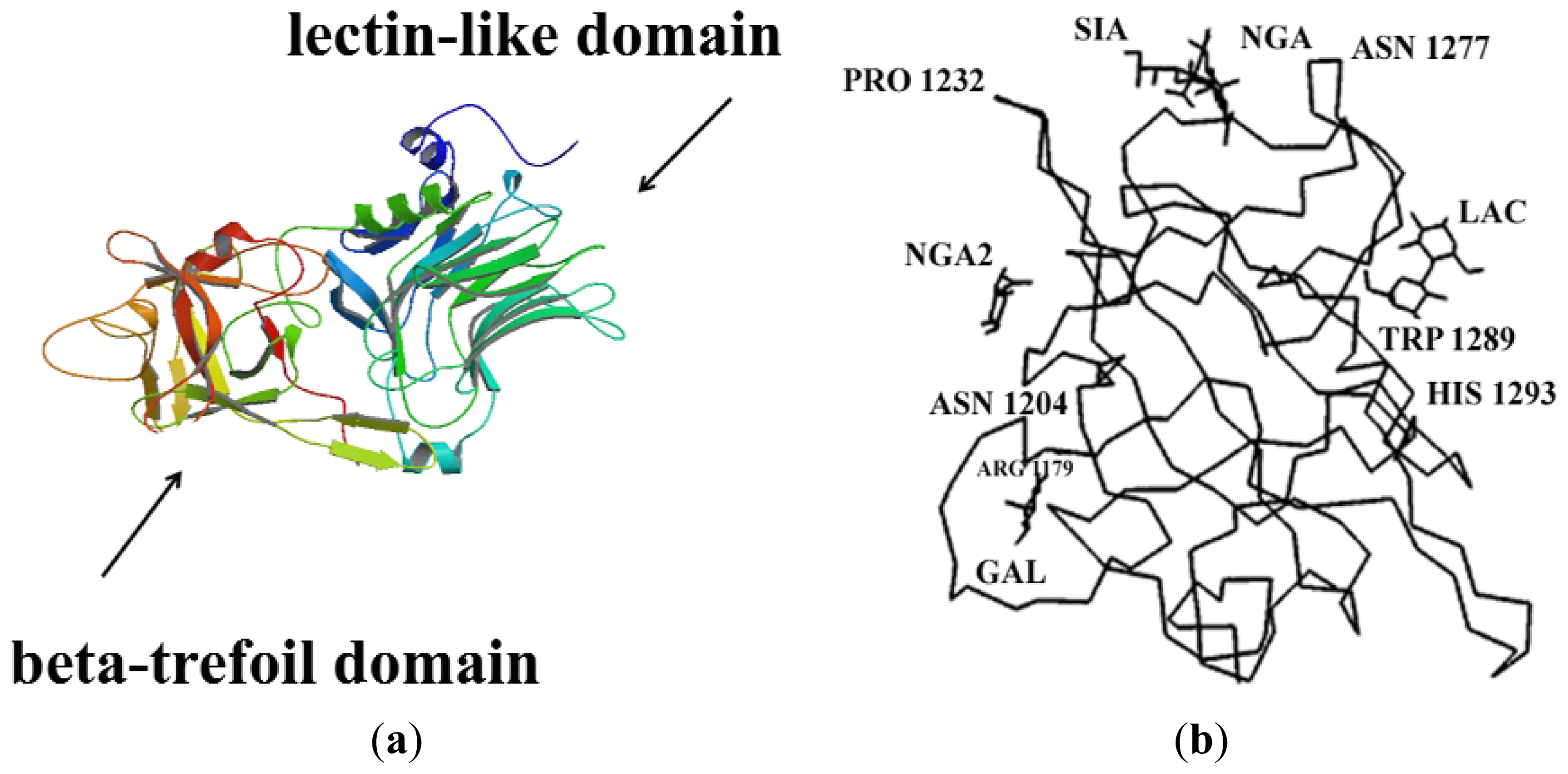
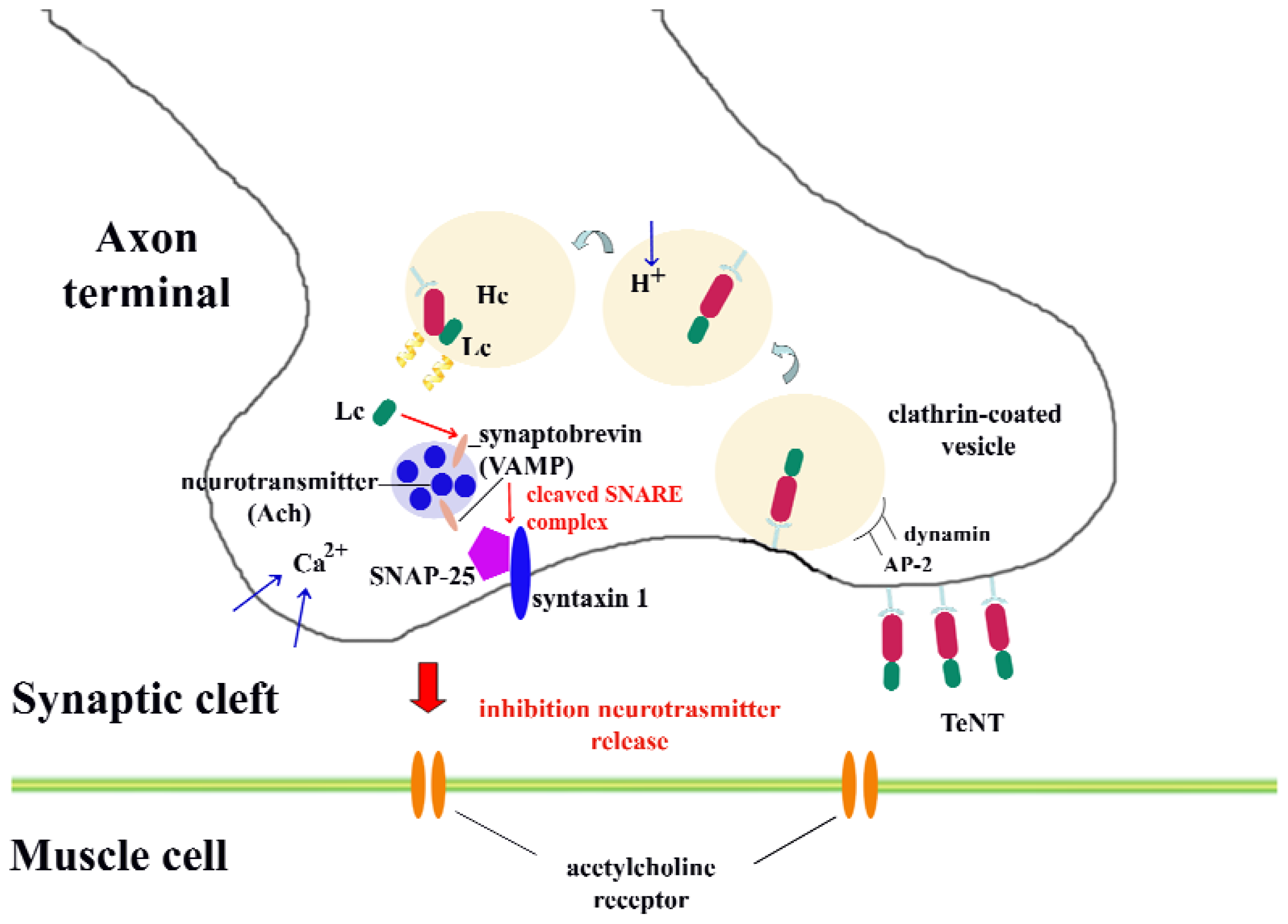
2. Molecular Structure and Properties of Fragment C: Toward the Discovery of Fragment C
- cell binding, mainly mediated by the ganglioside-recognition domain in the C-terminal region of HC;
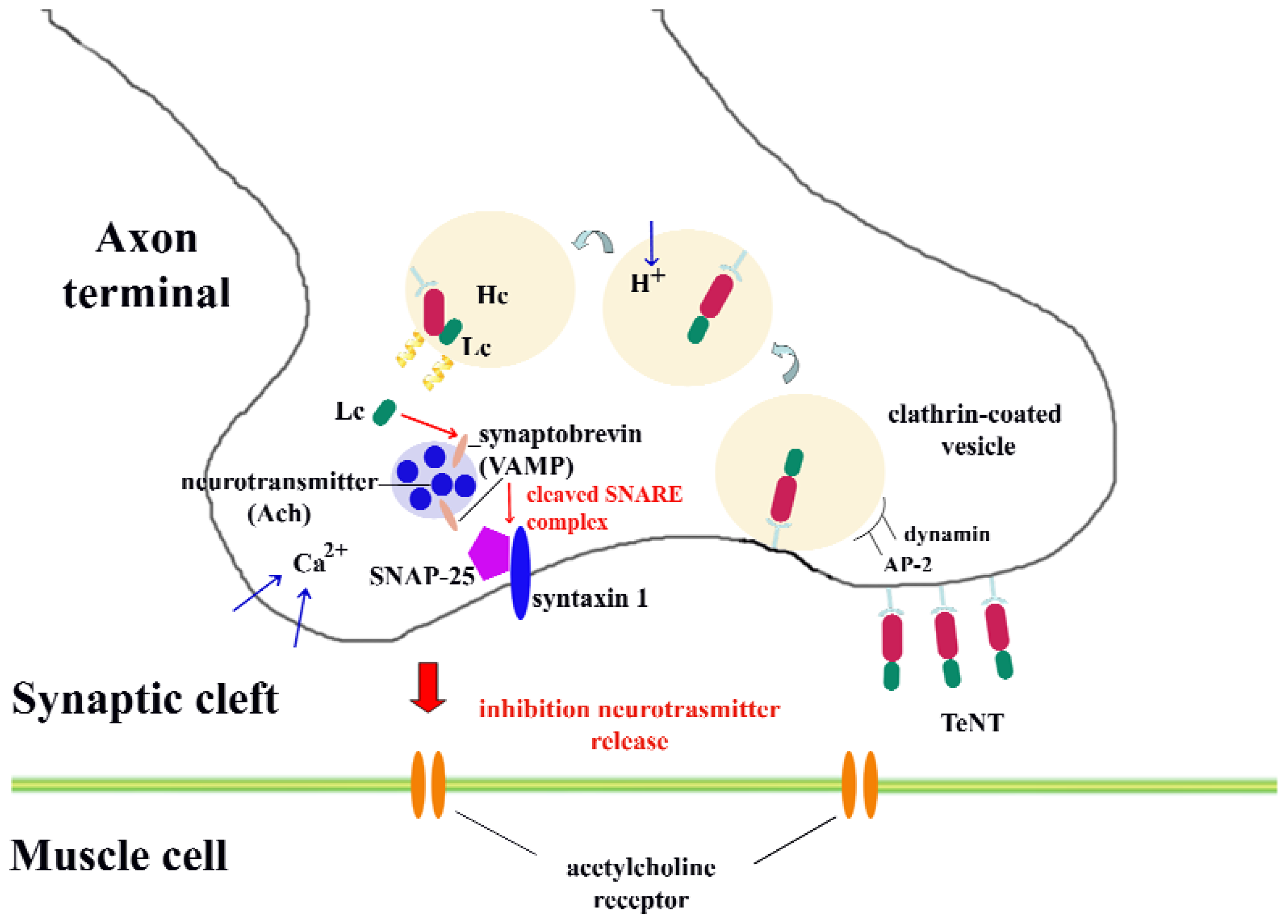
- internalization into neuronal cells. Different internalization mechanisms have been described: the mechanism that tetanus toxin possibly follows to internalize into neurons may be a clathrin-mediated pathway, which is dependent on the proteins dynamin, AP-2 and AP180 [30]. Other potential receptor molecules will be discussed in the next section.
- membrane translocation from mature endosomes into the neuronal cytoplasm. After incorporation of the toxin within endosomes, a structural change of the toxin is induced by the acidification of the endosomal environment; thus, a membrane-spanning pore forms. At this step, the oligomerization of four toxin amphipathic alpha-helices is required for channel formation; and
- target recognition and catalytic cleavage of neuronal substrates. Once the toxin reaches the cytoplasm, it specifically cleaves neuronal proteins integral to vesicular trafficking and neurotransmitter release. In particular, the synaptic vesicle protein synaptobrevin (VAMP) is the target of tetanus toxin. This protein belongs to a family of proteins that facilitate exocytosis in neurons known as SNARE proteins. The other members of this family are syntaxin and SNAP-25, which are the main molecular targets of botulinum toxin. SNARE proteins are formed by coiled-coil interactions of the alpha-helices of its members, which is required for membrane fusion [31–33].
2.1. Molecular Properties of Fragment C
2.1.1. Molecular Binding
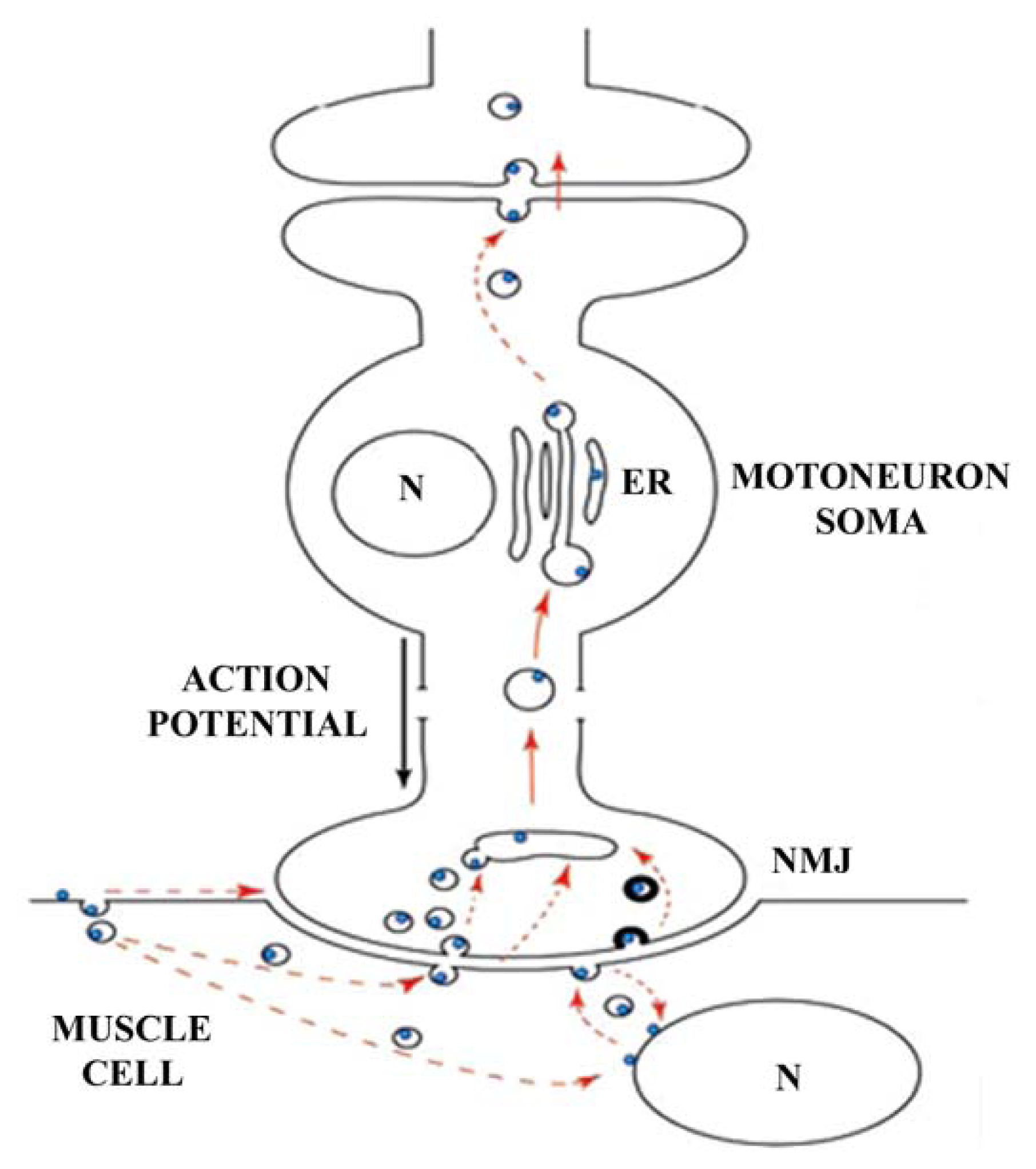
2.1.2. Retrograde Axonal Transport
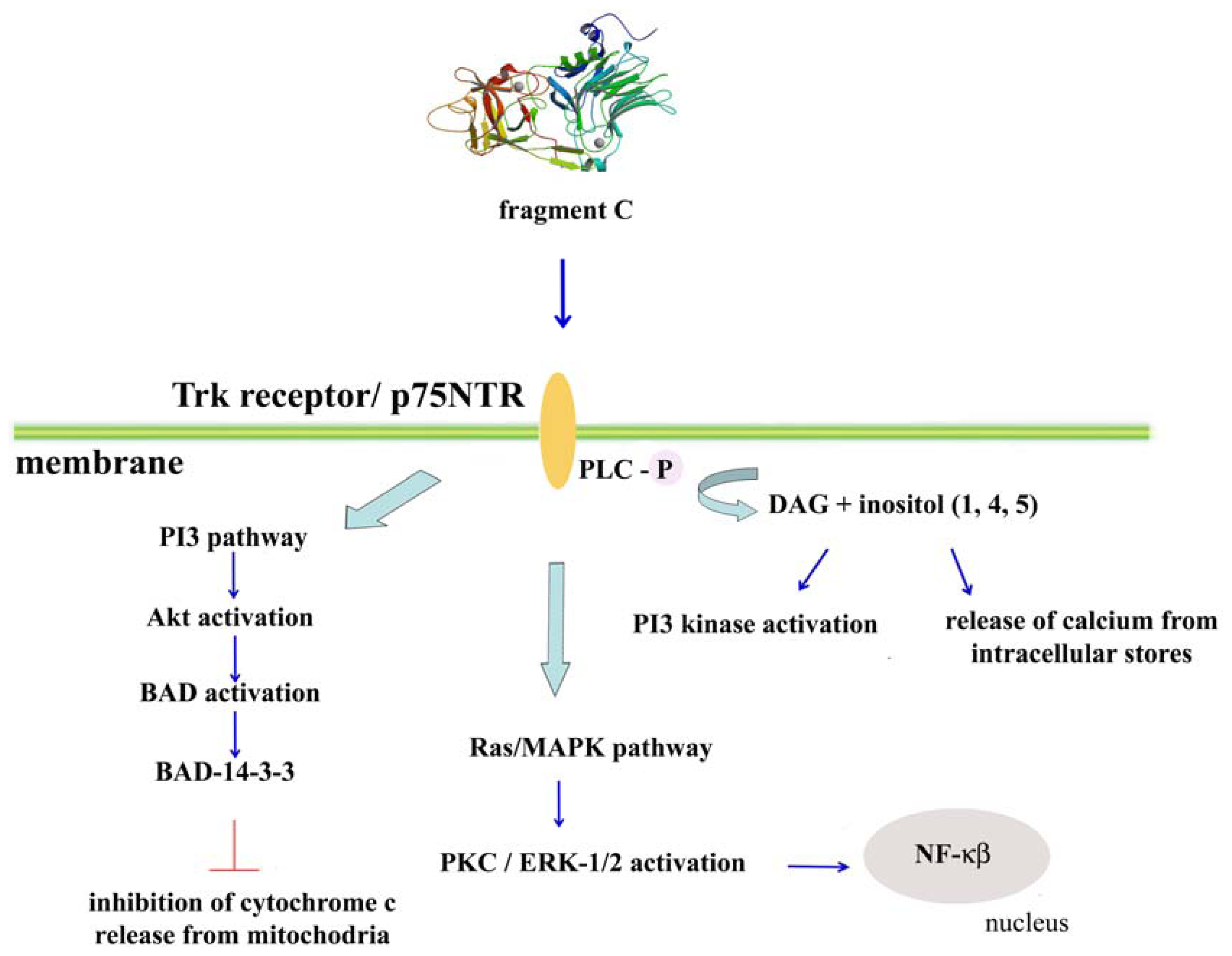
3. Possible Signaling Pathways for Fragment C of Tetanus Toxin
4. Conclusions
Acknowledgments
- Conflict of InterestThe authors declare no conflict of interest.
References
- Johnson, J.L.; Francis, G. Taxonomy of the clostridia: Ribosomal ribonucleic acid homologies among the species. J. Gen. Microbiol 1975, 88, 229–244. [Google Scholar]
- Johnson, E.A. Clostridial toxins as therapeutic agents: Benefits of natures’s most toxic proteins. Annu. Rev. Microbiol 1999, 53, 551–575. [Google Scholar]
- Anderson, J.F.; Leake, J.P. A method of producing tetanus toxin. J. Med. Res 1915, 33, 239–241. [Google Scholar]
- Noguchi, H. The nature of the antitetanic action of eosin. J. Exp. Med 1907, 9, 281–290. [Google Scholar]
- Flexner, S.; Noguchi, H. The effect of eosin upon tetanus toxin and upon tetanus in rats and guinea-pigs. J. Exp. Med 1906, 8, 1–7. [Google Scholar]
- Cowie, D.M.; Greenthal, R.M. Studies on the nature of the action of non-specific protein in disease processes. III. Non-specific proteins and soluble toxin (diphtheria-tetanus). J. Med. Res 1922, 43, 21–28. [Google Scholar]
- Sherrington, C.S. On reciprocal innervation of antagonistic muscles. VIIIth note. Proc. R. Soc. B 1905, 76, 269–297. [Google Scholar]
- Acheson, G.H.; Oscar, M.D.; Ratnoff, D.; Schoenbach, E.B. The localized action on the spinal cord of intramuscularly injected tetanus toxin. J. Exp. Med 1942, 75, 465–480. [Google Scholar]
- Brooks, V.B.; Curtis, D.R.; Eccles, J.C. The action of tetanus toxin on the inhibition of motor neurons. J. Physiol 1957, 135, 655–672. [Google Scholar]
- Firor, W.M.; Lamont, A. The apparent alteration of tetanus toxin within the spinal cord of dogs. Ann. Surg 1938, 108, 941–957. [Google Scholar]
- Martini, E.; Torda, C.; Zironi, A. The effect of tetanus toxin on the choline esterase activity of the muscles of rats. J. Physiol 1939, 96, 168–171. [Google Scholar]
- Harvey, A.M. The peripheral action of tetanus toxin. J. Physiol 1939, 96, 348–365. [Google Scholar]
- Manwaring, W.H. Types of tetanus toxin. Cal. West. Med 1943, 59, 306–307. [Google Scholar]
- Ipsen, J. The effect of environmental temperature on the reaction of mice to tetanus toxin. J. Immunol 1951, 66, 687–694. [Google Scholar]
- Wright, E.A. The effect of the injection of tetanus toxin into the central nervous system of rabbits. J. Immunol 1953, 71, 41–44. [Google Scholar]
- Roaf, M.D.; Sherrington, C.S. Experiments in examination of the locked jaw induced by tetanus toxin. J. Physiol 1906, 34, 315–331. [Google Scholar]
- Wassermann, A.; Takaki, T. Über Tetanusantitoxische Eigenschaften des normalen Centralnervensystems. Berl. Klin. Wochenschr 1898, 35, 5–6. [Google Scholar]
- Landsteiner, K.; Botteri, A. Über Verbindungen von Tetanustoxin mit Lipoiden IV. Zbl. Bakt. Orig 1906, 42, 562. [Google Scholar]
- Van Heyningen, W.E. The fixation of tetanus toxin by nervous tissue. J. Gen. Microbiol 1959, 20, 291–300. [Google Scholar]
- Van Heyningen, W.E. Chemical assay of the tetanus toxin receptor in nervous tissue. J. Gen. Microbiol 1959, 20, 301–309. [Google Scholar]
- Van Heyningen, W.E.; Miller, P.A. The fixation of tetanus toxin by ganglioside. J. Gen. Microbiol 1961, 24, 107–119. [Google Scholar]
- Van Heyningen, W.E. Binding of ganglioside by the chains of tetanus toxin. FEBS Lett 1976, 68, 5–7. [Google Scholar]
- Sugiyama, H. Clostridium botulinum neurotoxin. Microbiol. Rev 1980, 44, 419–448. [Google Scholar]
- Johnson, E.A. Clostridial toxins as therapeutic agents: Benefits of nature’s most toxic proteins. Ann. Rev. Microbiol 1999, 53, 551–575. [Google Scholar]
- Pellizari, R.; Rossetto, O.; Schiavo, G.; Montecucco, C. Tetanus and botulinum neurotoxins: Mechanism of action and therapeutic uses. Philos. Trans. R. Soc. Lond. B 1999, 354, 259–268. [Google Scholar]
- Montal, M. Botulinum neurotoxin. Annu. Rev. Biochem 2010, 79, 591–617. [Google Scholar]
- Habermann, E.; Dreyer, F. Clostridial neurotoxins: Handling and action at the cellular and molecular level. Curr. Top. Microbiol. Immunol 1986, 129, 93–179. [Google Scholar]
- Chen, S.; Karalewitz, A.P.A.; Barbieri, J.T. Insights into the different catalytic activities of Clostridium neurotoxins. Biochemistry 2012, 51, 3941–3947. [Google Scholar]
- Neubauer, V.; Helting, T.B. Structure of tetanus toxin: The arrangement of papain digestion products within the heavy chain-light chain framework of extracellular toxin. Biochim. Biophys. Acta 1981, 27, 141–148. [Google Scholar]
- Deinhardt, K.; Berninghausen, O.; Willison, H.J.; Hopkins, C.R.; Schiavo, G. Tetanus toxin is internalized by a sequential clathrin-dependent mechanism initiated within lipid microdomains and independent of epsin (eosin?) 1. J. Cell Biol 2006, 174, 459–471. [Google Scholar]
- Mochida, S. Protein-protein interactions in neurotransmitter release. Neurosci. Res 2000, 36, 175–182. [Google Scholar]
- Humeau, Y.; Doussau, F.; Grant, N.J.; Poulain, B. How botulinum and tetanus neurotoxins block neurotransmitter release. Biochimie 2000, 82, 427–446. [Google Scholar]
- Ungar, D.; Hughson, F.M. SNARE protein structure and function. Annu. Rev. Cell Dev. Biol 2003, 19, 493–517. [Google Scholar]
- Boquet, P.; Duflot, E.; Hauttecoeur, B. Low pH induces a hydrophobic domain in the tetanus toxin molecule. Eur. J. Biochem 1984, 144, 339–344. [Google Scholar]
- Simpson, L.L.; Hoch, D.H. Neuropharmacological characterization of fragment B from tetanus toxin. J. Pharmacol. Exp. Ther 1985, 232, 223–227. [Google Scholar]
- Menestrina, G.; Forti, S.; Gambale, F. Interaction of tetanus toxin with lipid vesicles. Effects of pH, surface charge and transmembrane potential on the kinetics of channel formation. Biophys. J 1989, 55, 393–405. [Google Scholar]
- Calappi, E.; Masserini, M.; Schiavo, G.; Montecucco, C.; Tettamanti, G. Lipid interaction of tetanus toxin. A calorimetric and fluorescence spectroscopy study. FEBS 1992, 309, 107–110. [Google Scholar]
- Habermann, E.; Albus, U. Interaction between tetanus toxin and rabbit kidney: A comparison with rat brain preparations. J. Neurochem 1986, 46, 1219–1226. [Google Scholar]
- Lazarovici, P.; Yanai, P.; Llavín, E. Molecular interactions between micellar polysialogangliosides and affinity-purified tetanotoxins in aqueous solution. J. Biol. Chem 1987, 262, 2645–2651. [Google Scholar]
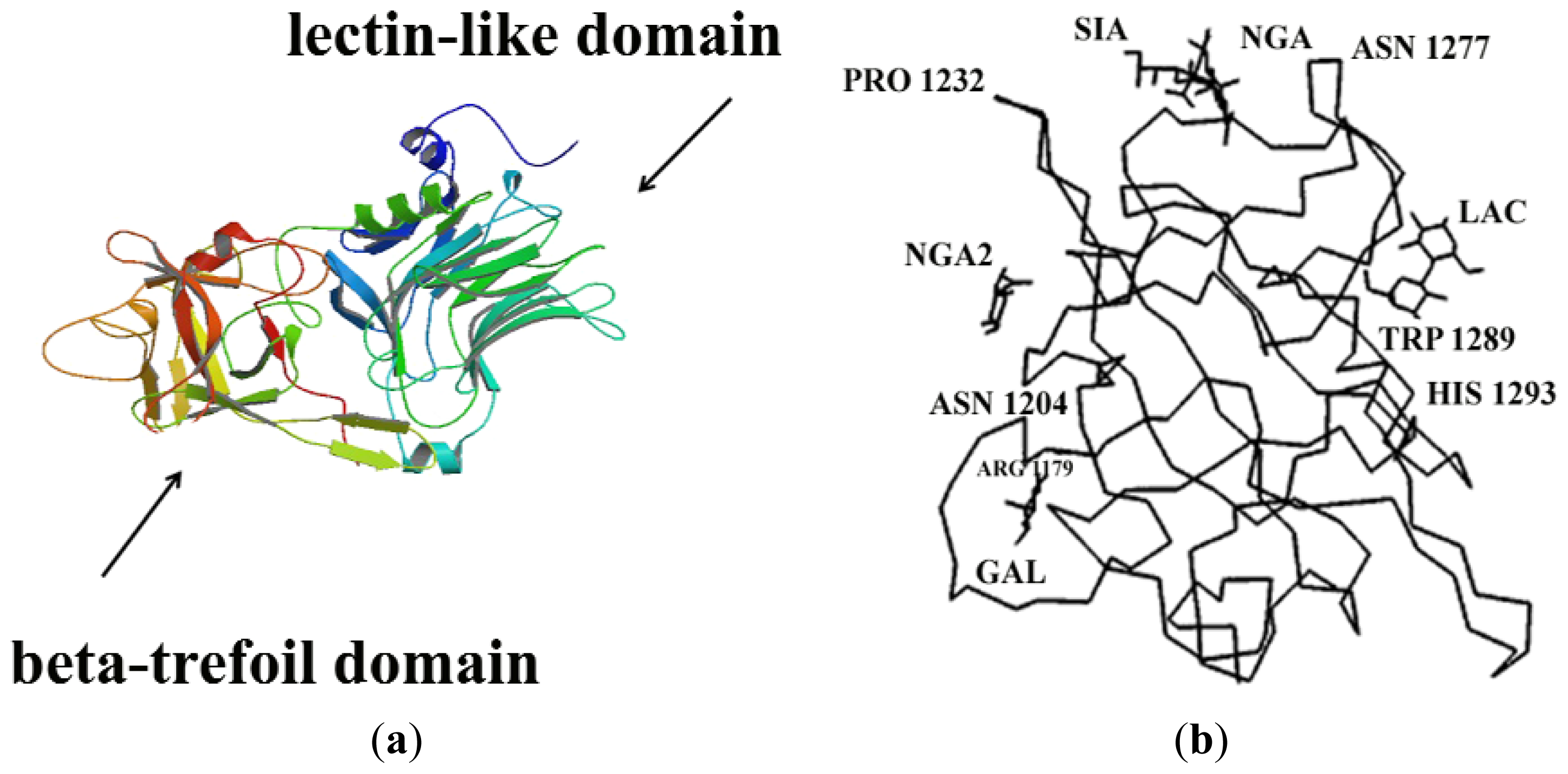
- Emsley, P.; Fotinou, C.; Black, I.; Fairweather, N.F.; Charles, I.G.; Watts, C.; Hewitt, E.; Isaacs, N.W. The structures of the HC fragment of tetanus toxin with carbohydrate subunit complexes provide insight into ganglioside binding. J. Biol. Chem 2000, 275, 8889–8894. [Google Scholar]
- Sinha, K.; Box, M.; Lalli, G.; Schiavo, G.; Schneider, H.; Groves, M.; Siligardi, G.; Fairweather, N. Analysis of mutants of tetanus toxin HC fragment: Ganglioside binding, cell binding and retrograde axonal transport properties. Mol. Microbiol 2000, 37, 1041–1051. [Google Scholar]
- Louch, H.A.; Buczko, E.S.; Woody, M.A.; Venable, R.M.; Vann, W.F. Identification of a binding site for ganglioside on the receptor binding domain of tetanus toxin. Biochemistry 2002, 41, 13644–13652. [Google Scholar]
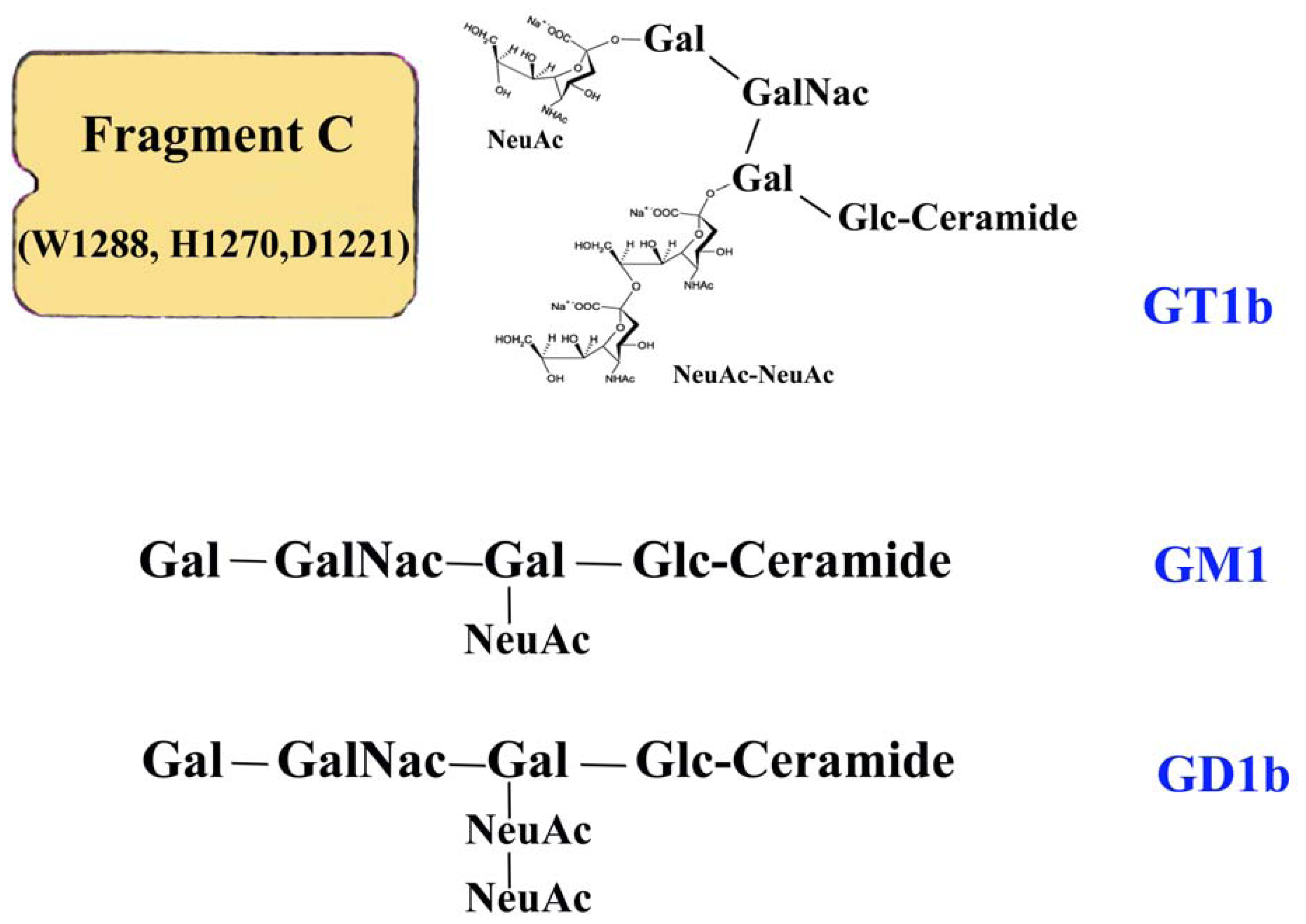
- Conway, P.M.C.; Whittal, R.M.; Baldwin, M.A.; Burlingame, A.L.; Balhorn, R. Electrospray mass spectrometry of NeuAc oligomers associated with the C fragment of the tetanus toxin. J. Am. Soc. Mass Spectrom 2006, 17, 967–976. [Google Scholar]
- Siade, A.L.; Schoeniger, J.S.; Sasaki, D.Y.; Yip, C.M. In situ canning probe microscopy studies of tetanus toxin-membrane interacions. Biophys. J 2006, 91, 4565–4574. [Google Scholar]
- Helting, T.B.; Zwisler, O.; Wiegandt, H. Structure of tetanus toxin. II. Toxin binding to ganglioside. J. Biol. Chem 1977, 252, 194–198. [Google Scholar]
- Sutton, J.M.; Chow-Worn, O.; Spaven, L.; Silman, N.J.; Hallis, B.; Shone, C.C. Tyrosyne-1290 of tetanus neurotoxin plays a key role in its binding to gangliosides and functional binding to neurons. FEBS Lett 2001, 493, 45–49. [Google Scholar]
- The 1.61 angstrom structure of the tetanus toxin ganglioside binding region: Solved by MAD and Mir phase combination. Available online: www.pdb.org accessed on 31 May 2012.
- Rummel, A.; Bade, S.; Alves, J.; Bigalke, H.; Binz, T. Two carbohydrate binding sites in the HCC-domain of tetanus neurotoxin are required for toxicity. J. Mol. Biol 2003, 326, 835–847. [Google Scholar]
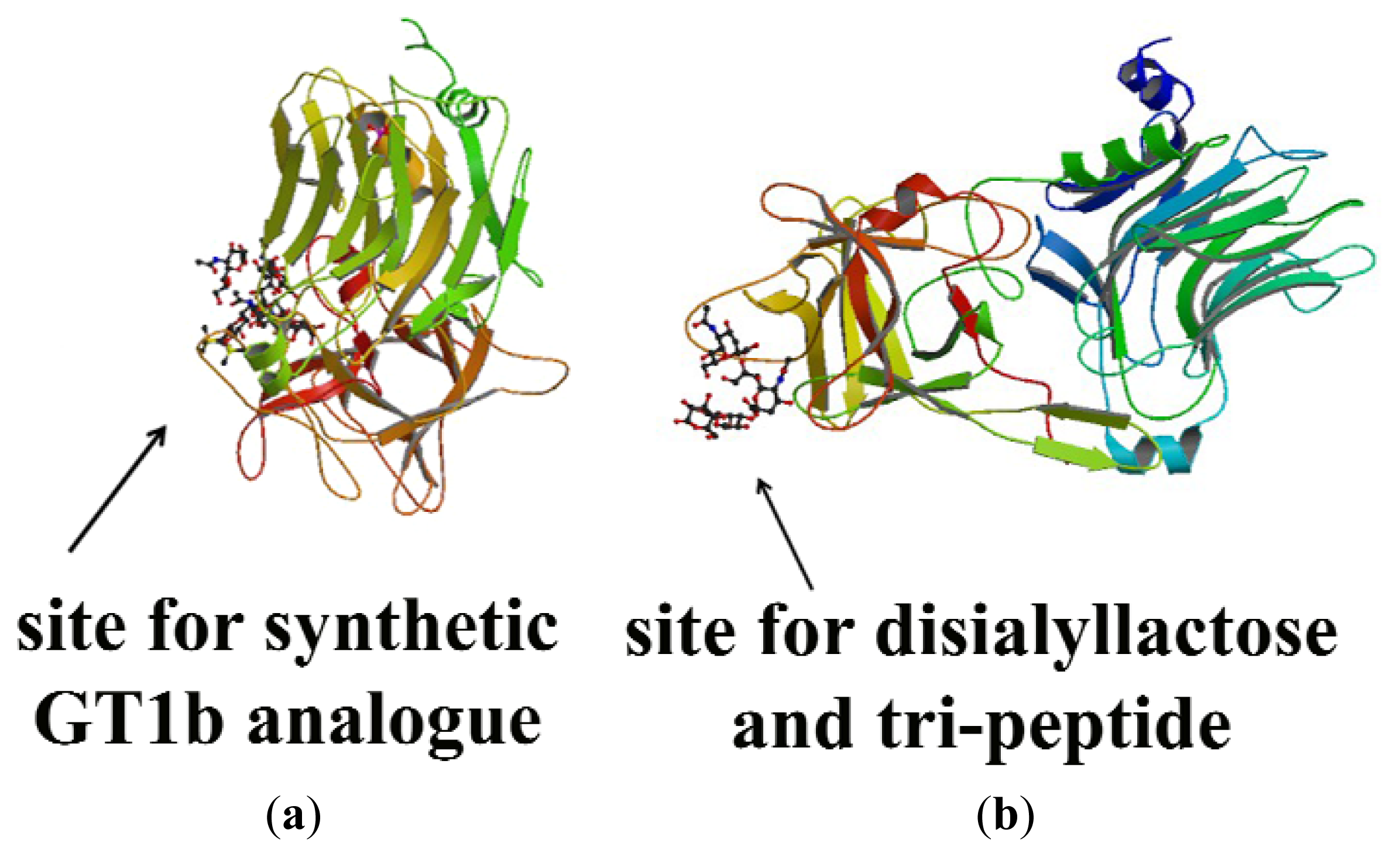
- Jayaraman, S.; Eswaramoorthy, S.; Kumaran, D.; Swaminathan, S. Common binding site for disialyllactose and tri-peptide in C-fragment of tetanus neurotoxin. Proteins Struct. Funct. Bioinform 2005, 61, 288–295. [Google Scholar]
- Cosman, M.; Lightstone, F.C.; Krishnan, V.V.; Zeller, L.; Prieto, M.C.; Roe, D.C.; Balhorn, R. Identification of novel small molecules that bind to two different sites on the surface of tetanus toxin C fragment. Chem. Res. Toxicol 2002, 15, 1218–1228. [Google Scholar]
- Fotinou, C.; Emsley, P.; Black, I.; Ando, H.; Ishida, H.; Kiso, M.; Sinha, K.A.; Fairweather, N.F.; Isaacs, W. The crystal structure of the tetanus-toxin HC fragment complexed with a synthetic GT1b analogue suggests cross-linking between ganglioside receptors and the toxin. J. Biol. Chem 2001, 276, 32274–32281. [Google Scholar]
- Jayaraman, S.; Swaramoorthy, S.; Kumaran, D.; Swaminathan, S. Common binging site for disialyllactose and tri-peptide in C-fragment of tetanus neurotoxin. Proteins 2005, 61, 288–295. [Google Scholar]
- Chen, C.; Fu, Z.; Kim, J.-J.P.; Barbieri, J.T.; Baldwin, M.R. Gangliosides as high affinity receptors for tetanus neurotoxin. J. Biol. Chem 2009, 284, 26569–26577. [Google Scholar]
- Herreros, J.; Lalli, G.; Montecucco, C.; Schiavo, G. Tetanus toxin fragment C binds to a protein present in neuronal cell lines and motorneurons. J. Neurochem 2000, 74, 1941–1950. [Google Scholar]
- Herreros, J.; Ng, T.; Schiavo, G. Lipid rafts act as specialized domains for tetanus toxin binding and internalization into neurons. Mol. Biol. Cell 2001, 12, 2947–2960. [Google Scholar]
- Deinhardt, K.; Salinas, K.; Verastigui, C.; Watson, R.; Worth, D.; Hanrahan, S.; Bucci, C.; Schiavo, G. Rab5 and Rab7 control endocytic sorting along the axonal retrograde transport pathway. Neuron 2006, 52, 293–305. [Google Scholar]
- Evinger, C.; Erichsen, J.T. Transsynaptic retrograde transport of fragment C of tetanus toxin demonstrated by immunohistochemical localization. Brain Res 1986, 380, 383–388. [Google Scholar]
- Manning, K.A.; Erichsen, J.T.; Evinger, C. Retrograde transneuronal transport properties of fragment C of tetanus toxin. Neuroscience 1990, 34, 251–263. [Google Scholar]
- Fishman, P.S.; Carrigan, D.R. Retrograde transneuronal transfer of the fragment C of tetanus toxin. Brain Res 1987, 406, 275–279. [Google Scholar]
- Coen, L.; Osta, R.; Maury, M.; Brûlet, P. Construction of hybrid proteins that migrate retrogradely and transynaptically into the central nervous system. Proc. Natl. Acad. Sci. USA 1997, 94, 9400–9405. [Google Scholar]
- Miana-Mena, F.J.; Muñoz, M.J.; Ciriza, J.; Soria, J.; Brûlet, P.; Zaragoza, P.; Osta, R. Fragment C tetanus toxin: A putative activity-dependent neuroanatomical tracer. Acta Neurobiol. Exp 2003, 63, 211–218. [Google Scholar]
- Miana-Mena, F.J.; Muñoz, M.J.; Roux, S.; Ciriza, J.; Zaragoza, P.; Brûlet, P.; Osta, R. A non-viral vector for targeting gene therapy to motoneurons in the CNS. Neurodegener. Dis 2004, 1, 101–108. [Google Scholar]
- Miana-Mena, F.J.; Roux, S.; Benichou, J.C.; Osta, R.; Brûlet, P. Neuronal activity-dependent membrane traffic at the neuromuscular junction. Proc. Natl. Acad. Sci. USA 2002, 99, 3234–3239. [Google Scholar]
- Barati, S.; Chegini, F.; Hurtado, P.; Rush, R.A. Hybrid tetanus toxin C fragment-diphtheria toxin translocation domain allows specific gene transfer into PC12 cells. Exp. Neurol 2002, 177, 75–87. [Google Scholar]
- Oliveira, H.; Fernandez, R.; Pires, L.R.; Martins, M.C.L.; Simões, S.; Barbosa, M.A.; Pêgo, A.P. Targeted gene delivery into peripheral sensorial neurons mediated by self-assembled vectors composed of poly (ethylene imine) and tetanus toxin fragment C. J. Control Release 2010, 143, 350–358. [Google Scholar]
- Larsen, K.E.; Benn, S.C.; Ay, I.; Chian, R.J.; Celia, S.A.; Remington, M.P.; Bejarano, M.; Liu, M.; Ross, J.; Carmillo, P.; et al. A glial cell line-derived neurotrophic factor (GDNF): Tetanus toxin fragment C protein conjugate improves delivery of GDNF to spinal cord motor neurons in mice. Brain Res 2006, 1120, 1–12. [Google Scholar]
- Ciriza, J.; Moreno-Igoa, M.; Calvo, A.C.; Yagüe, G.; Palacio, J.; Miana-Mena, F.J.; Muñoz, M.J.; Zaragoza, P.; Brûlet, P.; Osta, R. A genetic fusion GDNF-C fragment of tetanus toxin prolongs survival in a symptomatic mouse ALS model. Restor. Neurol. Neurosci 2008, 26, 459–465. [Google Scholar]
- Moreno-Igoa, M.; Calvo, A.C.; Ciriza, J.; Muñoz, M.J.; Zaragoza, P.; Osta, R. Non-viral gene delivery of the GDNF, either alone or fused to the C-fragment of tetanus toxin protein, prolongs survival in a mouse ALS model. Restor. Neurol. Neurosci 2012, 30, 69–80. [Google Scholar]
- Calvo, A.C.; Moreno-Igoa, M.; Mancuso, R.; Manzano, R.; Oliván, S.; Munoz, M.J.; Penas, C.; Zaragoza, P.; Navarro, X.; Osta, R. Lack of a synergistic effect of a non-viral ALS gene therapy based on BDNF and a TTC fusion molecule. Orphanet J. Rare Dis 2011. [Google Scholar] [CrossRef]
- Bordet, T.; Castelnau-Ptakhine, L.; Fauchereau, F.; Friocourt, G.; Kahn, A.; Haase, G. Neuronal targeting of cardiotrophin-1 by coupling with tetanus toxin C fragment. Mol. Cell. Neurosci 2001, 17, 842–854. [Google Scholar]
- Carlton, E.; Teng, Q.; Federici, T.; Yang, J.; Riley, J.; Boulis, N.M. Fusion of the tetanus toxin C fragment binding domain and Bcl-XL for protection of peripheral-nerve neurons. Neurosurgery 2008, 63, 1175–1184. [Google Scholar]
- Lalli, G.; Schiavo, G. Analysis of retrograde transport in motor neurons reveals common endocytic carriers for tetanus toxin and neurotrophin receptor p75NTR. J. Cell Biol 2002, 156, 233–239. [Google Scholar]
- Ibánez, C.F.; Simi, A. p75 neurotrophin receptor signaling in nervous system injury and degeneration: Paradox and opportunity. Trends Neurosci 2012. [Google Scholar] [CrossRef]
- Roux, S.; Saint Cloment, C.; Curie, T.; Girard, E.; Mena, F.J.; Barbier, J.; Osta, R.; Molgó, J.; Brûlet, P. Brain-derived neurotrophic factor facilitates in vivo internalization of tetanus neurotoxin C-terminal fragment fusion proteins in mature mouse motor nerve terminals. Eur. J. Neurosci 2006, 24, 1546–1554. [Google Scholar]
- Skeldal, S.; Matusica, D.; Nykjaer, A.; Coulson, E.J. Proteolytic processing of the p75 neurotrophin receptor: A prerequisite for signalling? Neuronal life, growth and death signalling are crucially regulated by intra-membrane proteolysis and trafficking of p75(NTR). Bioessays 2011, 33, 614–625. [Google Scholar]
- Skaper, S.D. The biology of neurotrophins, signalling pathways, and functional peptide mimetics of neurotrophins and their receptors. CNS Neurol. Disord. Drug Targets 2008, 7, 46–62. [Google Scholar]
- Deinhardt, K.; Reversi, A.; Berninghausen, O.; Hopkins, C.R.; Schiavo, G. Neurotrophins redirect p75NTR from a clathrin-independent to a clathrin-dependent endocytic pathway coupled to axonal transport. Traffic 2007, 8, 1736–1749. [Google Scholar]
- Wang, K.C.; Kim, J.A.; Sivasankaran, R.; Segal, R.; He, Z. p75 interacts with the Nogo receptor as a co-receptor for Nogo, MAG and OMgp. Nature 2002, 420, 74–78. [Google Scholar]
- Twiss, J.L.; Chang, J.H.; Schanen, N.C. Pathophysiological mechanisms for actions of the neurotrophins. Brain Pathol 2006, 16, 320–332. [Google Scholar]
- Aguilera, J.; Lopez, L.A.; Yavin, E. Tetanus toxin-induced protein kinase C activation and elevated serotonin levels in the perinatal rat brain. FEBS 1990, 263, 61–65. [Google Scholar]
- Gil, C.; Ruiz-Meana, M.; Álava, M.; Yavin, E.; Aguilera, J. Tetanus toxin enhances protein kinase C activity translocation and increases polyphosphoinositide hydrolysis in rat cerebral cortex preparations. J. Neurochem 1998, 70, 1636–1643. [Google Scholar]
- Inserte, J.; Najib, A.; Pelliccioni, P.; Gil, C.; Aguilera, J. Inhibition by tetanus toxin of sodium-dependent, high-affinity [3H]5-hydroxitryptamine uptake in rat synaptosomes. Biochem. Pharmacol 1999, 57, 111–120. [Google Scholar]
- Gil, C.; Chaib, I.; Pelliccioni, P.; Aguilera, J. Activation of signal transduction pathways involving TrkA, PLCγ-1, PKC isoforms and ERK-1/2 by tetanus toxin. FEBS Lett 2000, 481, 177–182. [Google Scholar]
- Pelliccioni, P.; Gil, C.; Najib, A.; Sarri, E.; Picatoste, F.; Aguilera, J. Tetanus toxin modulates serotonin transport in rat-brain neuronal cultures. J. Mol. Neurosci 2001, 17, 303–310. [Google Scholar]
- Gil, C.; Chaib-Oukadour, I.; Blasi, J.; Aguilera, J. HC fragment (C-terminal portion of the heavy chain) of tetanus toxin activates protein kinase C isoforms and phosphoproteins involved in signal transduction. Biochem. J 2001, 356, 97–103. [Google Scholar]
- Gil, C.; Chaib-Oukadour, I.; Aguilera, J. C-terminal fragment of tetanus toxin heavy chain activates Akt and MEK/ERK signalling pathways in a Trk receptor-dependent manner in cultured cortical neurons. Biochem. J 2003, 373, 613–620. [Google Scholar]
- Chaib-Oukadour, I.; Gil, C.; Aguilera, J. The C-terminal domain of heavy chain of tetanus toxin rescues cerebellar granule neurons from apoptotic death: Involvement of phosphatidylinositol 3-kinase and mitogen-activated protein kinase pathways. J. Neurochem 2004, 90, 1227–1236. [Google Scholar]
- Mendieta, L.; Venegas, B.; Moreno, N.; Patricio, A.; Martínez, I.; Aguilera, J.; Limón, I.D. The carboxyl-terminal domain of the heavy chain of tetanus toxin prevents dopaminergic degeneration and improves motor behaviour in rats with striatal MPP+-lesions. Neurosci. Res 2009, 65, 98–106. [Google Scholar]
- Chaib-Oukadour, I.; Gil, C.; Rodríguez-Álvarez, J.; Ortega, A.; Aguilera, J. Tetanus toxin HC fragment reduces neuronal MPP+ toxicity. Mol. Cell. Neurosci 2009, 41, 297–303. [Google Scholar]
- Thorne, R.G.; Frey, W.H. Delivery of neurotrophic factors to the central nervous system: Pharmacokinetic considerations. Clin. Pharmacokinet 2001, 40, 907–946. [Google Scholar]
- Borasio, G.D.; Robberecht, W.; Leigh, P.N.; Emile, J.; Guiloff, R.J.; Jerusalem, F.; Silani, V.; Vos, P.E.; Wokke, J.H.; Dobbins, T. A placebo-controlled trial of insulin-like growth factor-I in amyotrophic lateral sclerosis. European ALS/IGF-I Study Group. Neurology 1998, 51, 583–586. [Google Scholar]
- Check, E. Harmful potential of viral vectors fuels doubts over gene therapy. Nature 2003, 423, 573–574. [Google Scholar]
- Moreno-Igoa, M.; Calvo, A.C.; Penas, C.; Manzano, R.; Oliván, S.; Muñoz, M.J.; Mancuso, R.; Zaragoza, P.; Aguilera, J.; Navarro, X.; et al. Fragment C of tetanus toxin, more than a carrier. Novel perspectives in non-viral ALS gene therapy. J. Mol. Med 2010, 88, 297–308. [Google Scholar]


© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Calvo, A.C.; Oliván, S.; Manzano, R.; Zaragoza, P.; Aguilera, J.; Osta, R. Fragment C of Tetanus Toxin: New Insights into Its Neuronal Signaling Pathway. Int. J. Mol. Sci. 2012, 13, 6883-6901. https://doi.org/10.3390/ijms13066883
Calvo AC, Oliván S, Manzano R, Zaragoza P, Aguilera J, Osta R. Fragment C of Tetanus Toxin: New Insights into Its Neuronal Signaling Pathway. International Journal of Molecular Sciences. 2012; 13(6):6883-6901. https://doi.org/10.3390/ijms13066883
Chicago/Turabian StyleCalvo, Ana C., Sara Oliván, Raquel Manzano, Pilar Zaragoza, José Aguilera, and Rosario Osta. 2012. "Fragment C of Tetanus Toxin: New Insights into Its Neuronal Signaling Pathway" International Journal of Molecular Sciences 13, no. 6: 6883-6901. https://doi.org/10.3390/ijms13066883