Gelam Honey Has a Protective Effect against Lipopolysaccharide (LPS)-Induced Organ Failure

Abstract
:1. Introduction
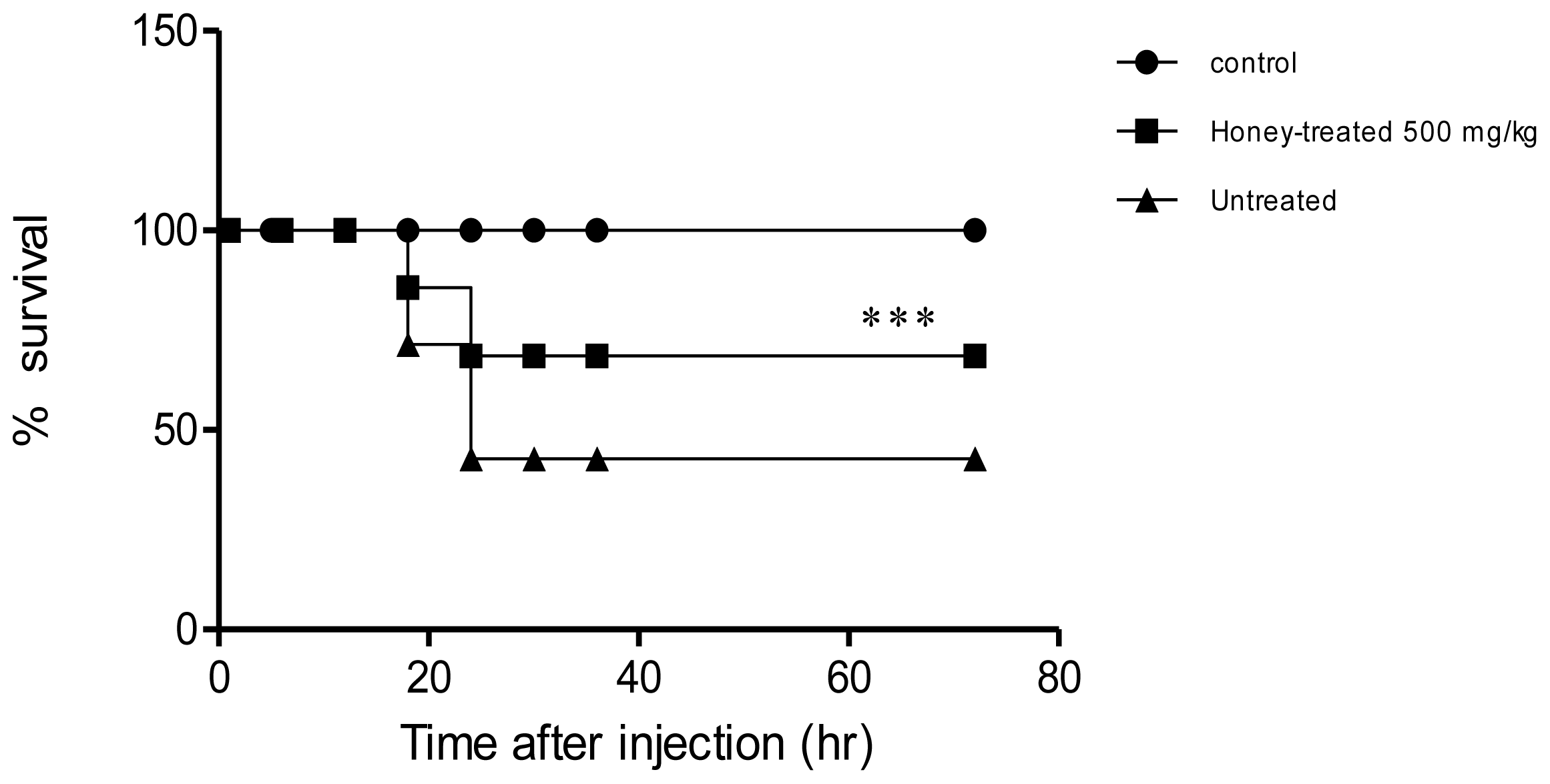
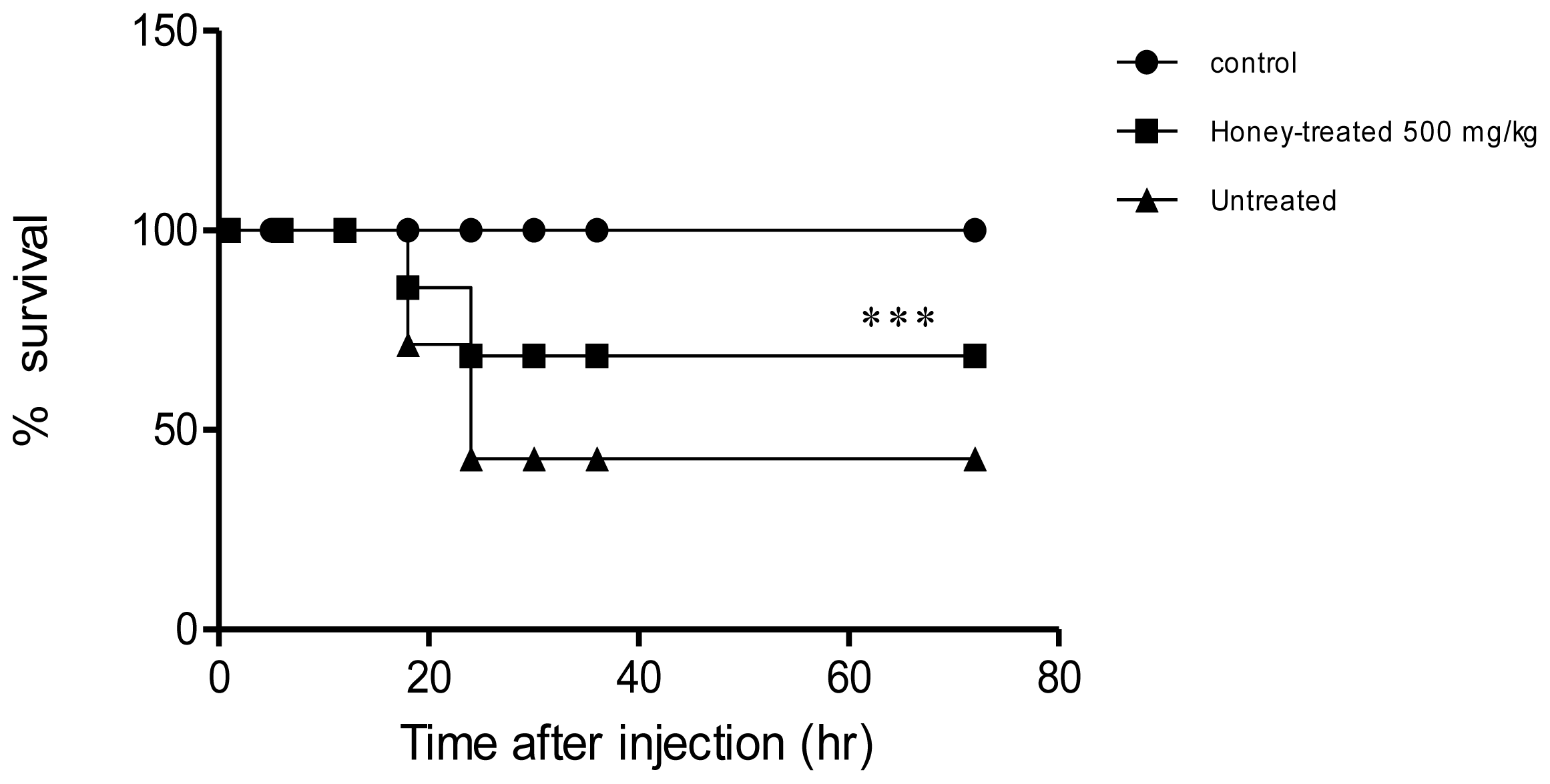
2. Results
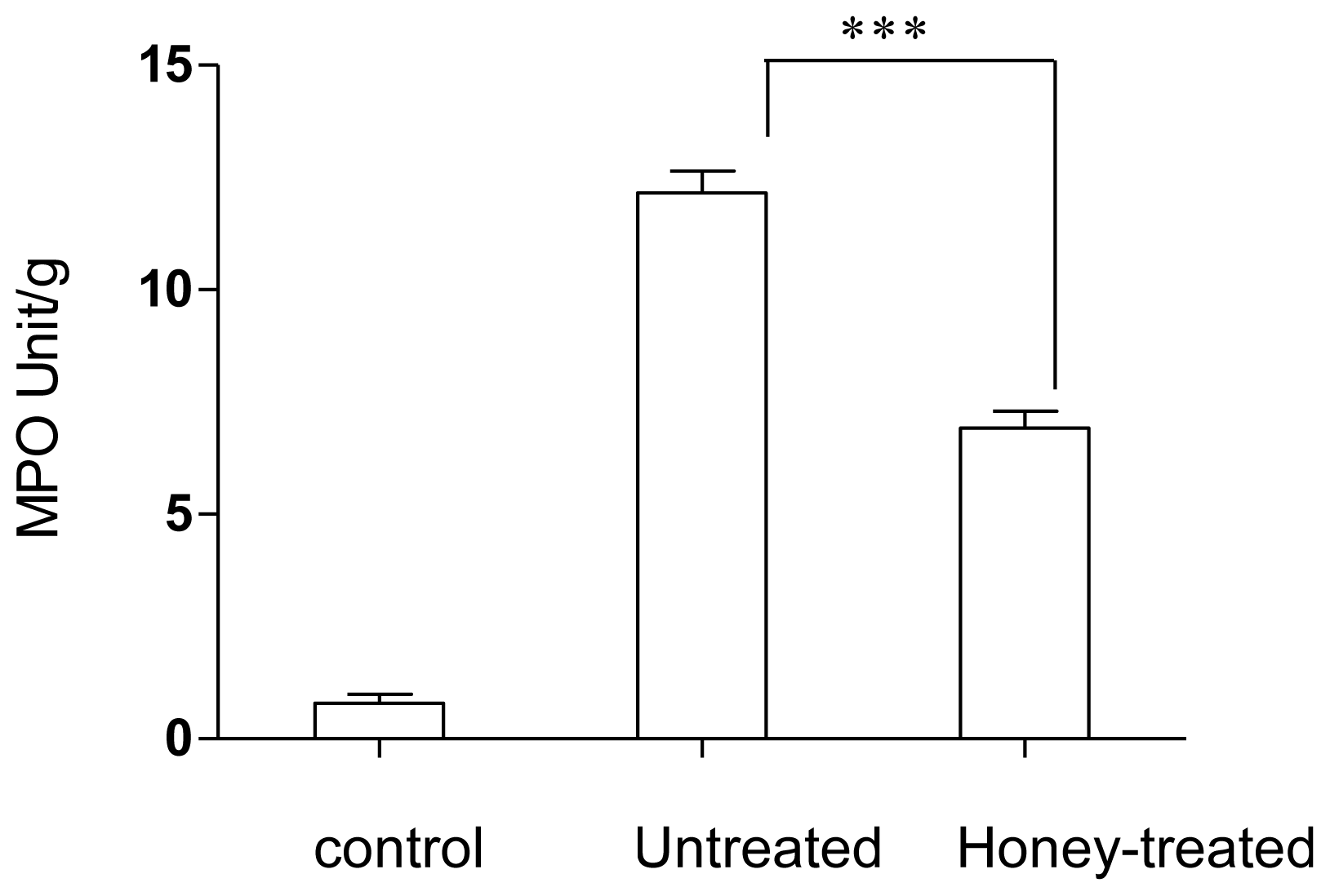
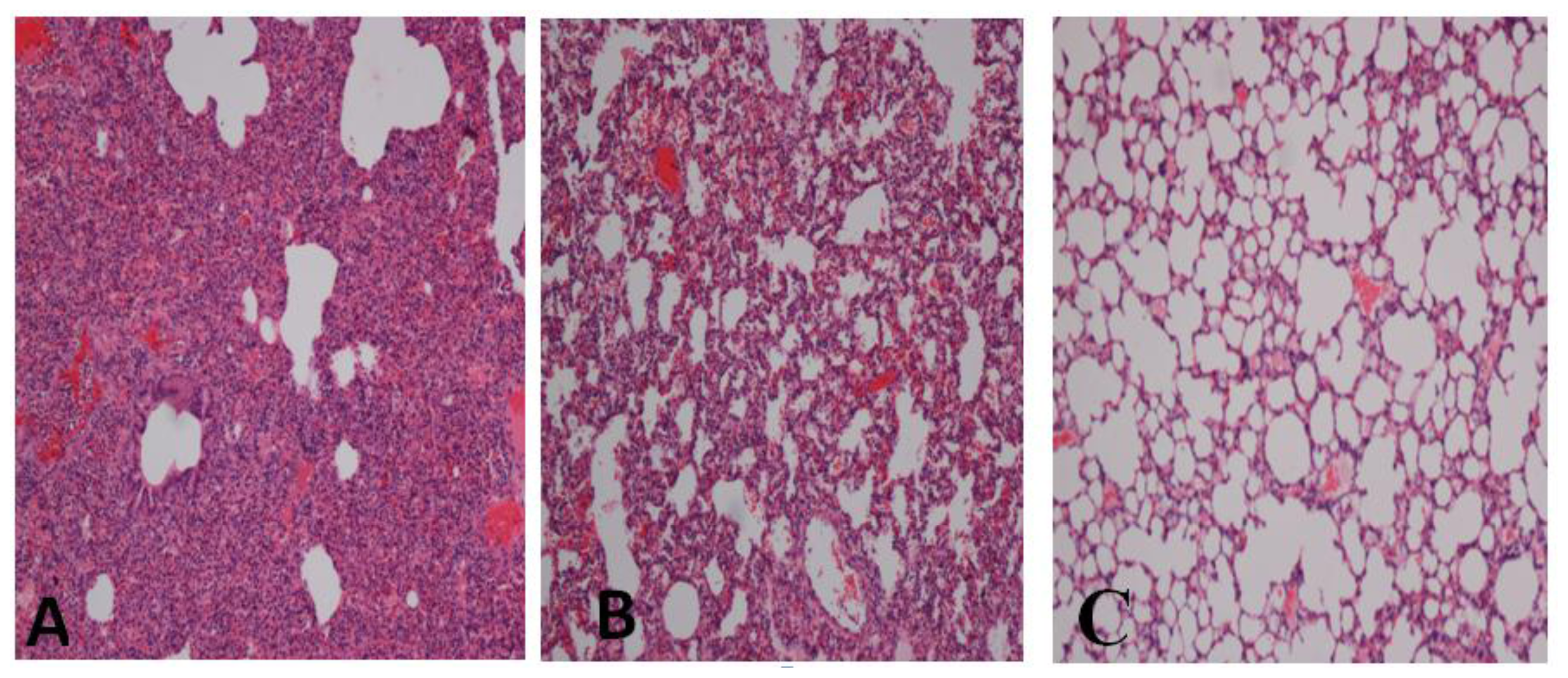
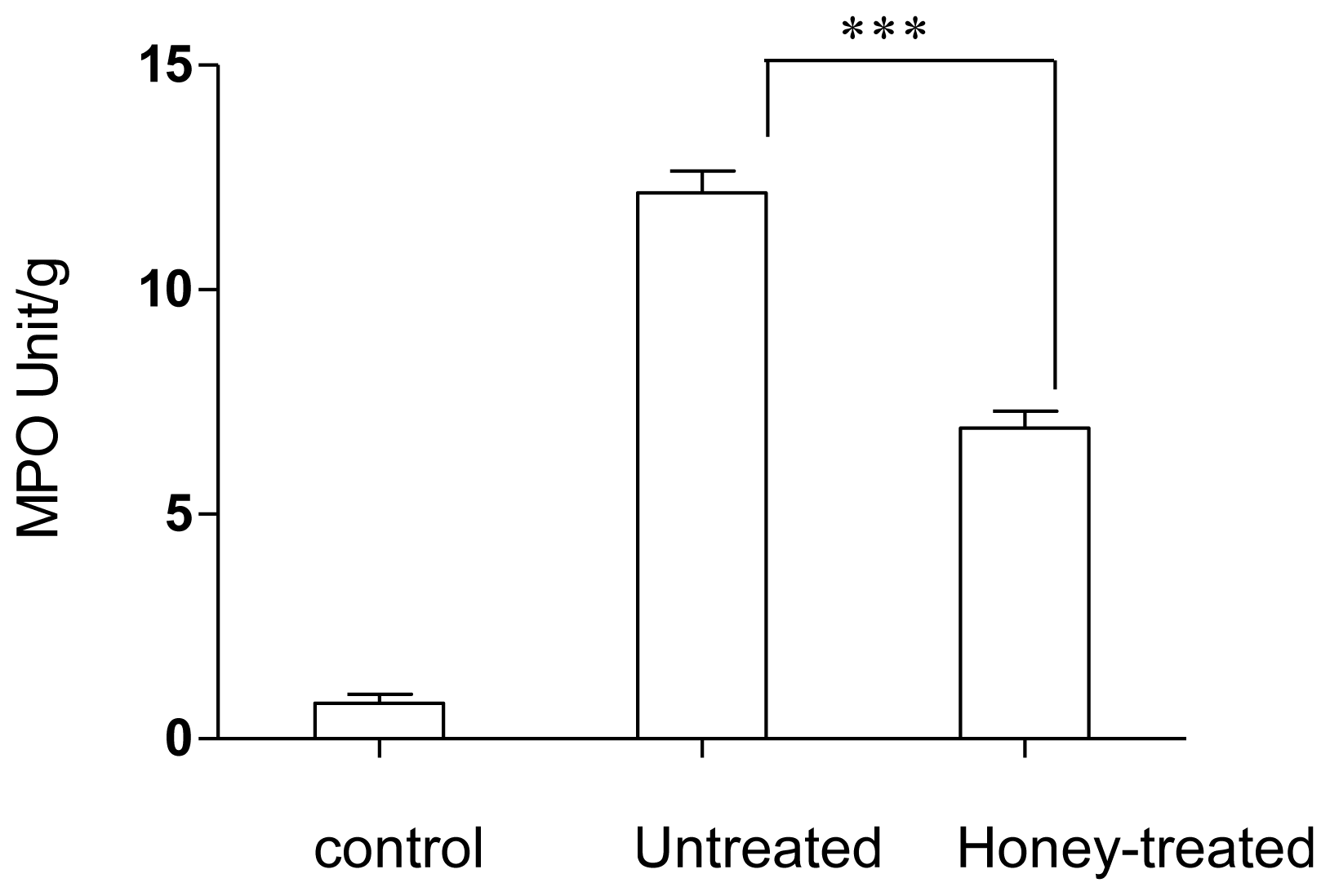
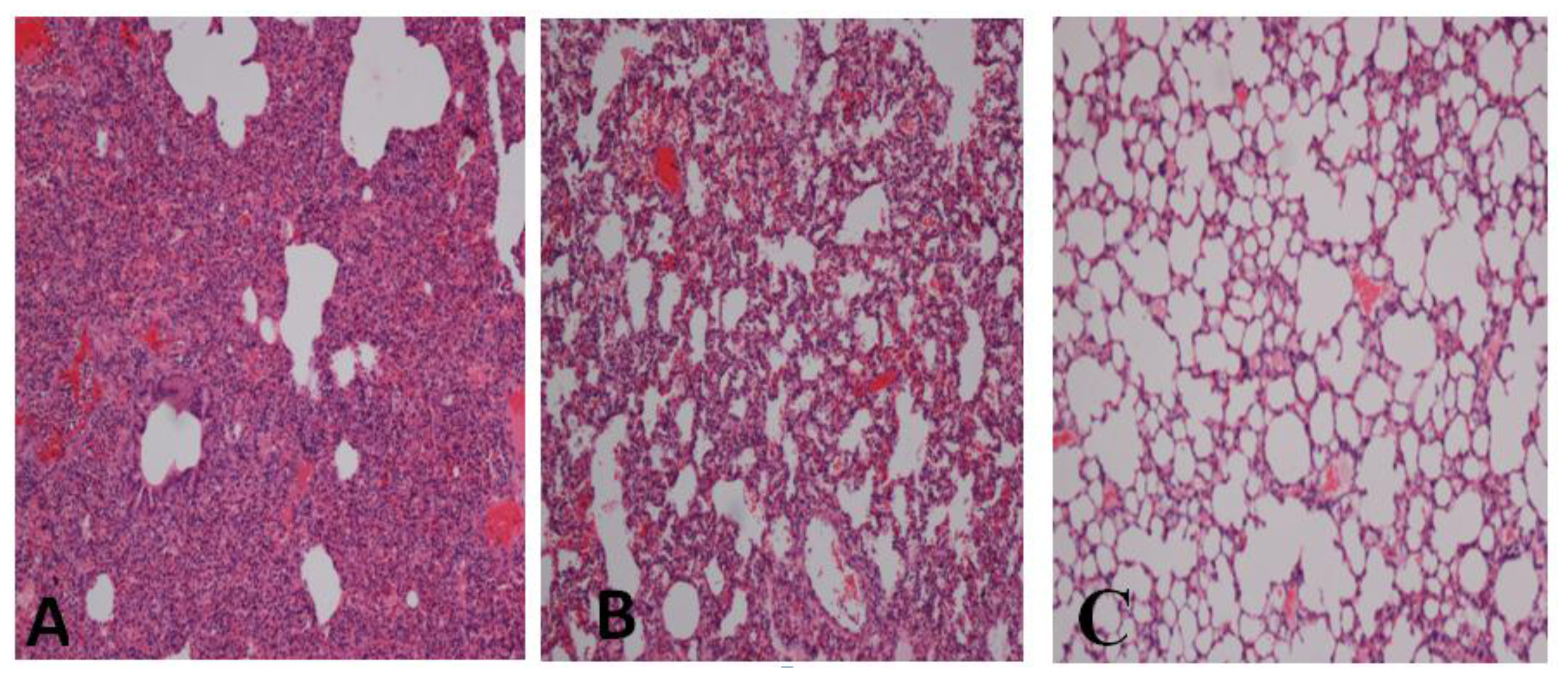
2.1. Effect of Gelam Honey on Biochemical and Hematological Tests, Histopathology, and MPO Activity
3. Discussion
4. Experimental Section
4.1. Preparation of Honey
4.2. Animals
4.3. Toxicity Tests
4.4. Induction of an Immune Response in Rabbits by LPS Stimulation and Treatment with Honey
4.5. Biochemical Analysis
4.6. Myeloperoxidase Assay
4.7. Histopathology
4.8. Statistical Analysis
5. Conclusions
Acknowledgments
- Conflict of InterestThe authors declare no conflict of interest.
References
- Lazaridou, A.; Biliaderis, C.G.; Bacandritsos, N.; Sabatini, A.G. Composition, thermal and rheological behaviour of selected greek honeys. J. Food Eng 2004, 64, 9–21. [Google Scholar]
- Gheldof, N.; Wang, X.H.; Engeseth, N.J. Buckwheat honey increases serum antioxidant capacity in humans. J. Agric. Food Chem 2003, 51, 1500–1505. [Google Scholar]
- White, R. The benefits of honey in wound management. Nurs. Stand 2005, 20, 57–64, quiz 66. [Google Scholar]
- Molan, P.C. The evidence supporting the use of honey as a wound dressing. Int. J. Low. Extrem. Wounds 2006, 5, 40–54. [Google Scholar]
- Tonks, A.J.; Dudley, E.; Porter, N.G.; Parton, J.; Brazier, J.; Smith, E.L.; Tonks, A. A 5.8-kda component of manuka honey stimulates immune cells via TLR4. J. Leukoc. Biol 2007, 82, 1147–1155. [Google Scholar]
- Molan, P.C. Potential of honey in the treatment of wounds and burns. Am. J. Clin. Dermatol 2001, 2, 13–19. [Google Scholar]
- Majtan, J.; Kovacova, E.; Bilikova, K.; Simuth, J. The immunostimulatory effect of the recombinant apalbumin 1-major honeybee royal jelly protein-on tnfalpha release. Int. Immunopharmacol 2006, 6, 269–278. [Google Scholar]
- Leong, A.G.; Herst, P.M.; Harper, J.L. Indigenous new zealand honeys exhibit multiple anti-inflammatory activities. Innate Immun 2011. [Google Scholar] [CrossRef]
- Kassim, M.; Achoui, M.; Mansor, M.; Yusoff, K.M. The inhibitory effects of gelam honey and its extracts on nitric oxide and prostaglandin E(2) in inflammatory tissues. Fitoterapia 2010, 81, 1196–1201. [Google Scholar]
- Kassim, M.; Achoui, M.; Mustafa, M.R.; Mohd, M.A.; Yusoff, K.M. Ellagic acid, phenolic acids, and flavonoids in malaysian honey extracts demonstrate in vitro anti-inflammatory activity. Nutr. Res 2010, 30, 650–659. [Google Scholar]
- Wang, H.; Li, W.; Li, J.; Rendon-Mitchell, B.; Ochani, M.; Ashok, M.; Yang, L.; Yang, H.; Tracey, K.J.; Wang, P.; et al. The aqueous extract of a popular herbal nutrient supplement, angelica sinensis, protects mice against lethal endotoxemia and sepsis. J. Nutr 2006, 136, 360–365. [Google Scholar]
- Bone, R.C.; Grodzin, C.J.; Balk, R.A. Sepsis: A new hypothesis for pathogenesis of the disease process. Chest 1997, 112, 235–243. [Google Scholar]
- Bohlinger, I.; Leist, M.; Gantner, F.; Angermuller, S.; Tiegs, G.; Wendel, A. DNA fragmentation in mouse organs during endotoxic shock. Am. J. Pathol 1996, 149, 1381–1393. [Google Scholar]
- Angus, D.C.; Linde-Zwirble, W.T.; Lidicker, J.; Clermont, G.; Carcillo, J.; Pinsky, M.R. Epidemiology of severe sepsis in the united states: Analysis of incidence, outcome, and associated costs of care. Crit. Care Med 2001, 29, 1303–1310. [Google Scholar]
- Cohen, J. The immunopathogenesis of sepsis. Nature 2002, 420, 885–891. [Google Scholar]
- Al-Jabri, A.A. Honey, milk and antibiotics. Afr. J. Biotechnol 2005, 4, 1580–1587. [Google Scholar]
- Molan, P.C. The potential of honey to promote oral wellness. Gen. Dent 2001, 49, 584–589. [Google Scholar]
- Van Deventer, S.J.; ten Cate, J.W.; Tytgat, G.N. Intestinal endotoxemia. Clinical significance. Gastroenterology 1988, 94, 825–831. [Google Scholar]
- Wenzel, R.P.; Pinsky, M.R.; Ulevitch, R.J.; Young, L. Current understanding of sepsis. Clin. Infect. Dis 1996, 22, 407–412. [Google Scholar]
- Tracey, K.J.; Lowry, S.F.; Beutler, B.; Cerami, A.; Albert, J.D.; Shires, G.T. Cachectin/tumor necrosis factor mediates changes of skeletal muscle plasma membrane potential. J. Exp. Med 1986, 164, 1368–1373. [Google Scholar]
- Schafer, T.; Scheuer, C.; Roemer, K.; Menger, M.D.; Vollmar, B. Inhibition of p53 protects liver tissue against endotoxin-induced apoptotic and necrotic cell death. FASEB J 2003, 17, 660–667. [Google Scholar]
- Essani, N.A.; McGuire, G.M.; Manning, A.M.; Jaeschke, H. Endotoxin-induced activation of the nuclear transcription factor kappa B and expression of E-selectin messenger RNA in hepatocytes, Kupffer cells, and endothelial cells in vivo. J. Immunol 1996, 156, 2956–2963. [Google Scholar]
- Dustin, M.L.; Springer, T.A. Role of lymphocyte adhesion receptors in transient interactions and cell locomotion. Annu. Rev. Immunol 1991, 9, 27–66. [Google Scholar]
- Luce, J.M. Acute lung injury and the acute respiratory distress syndrome. Crit. Care Med 1998, 26, 369–376. [Google Scholar]
- Vincent, J.L.; Sakr, Y.; Ranieri, V.M. Epidemiology and outcome of acute respiratory failure in intensive care unit patients. Crit. Care Med 2003, 31, S296–S299. [Google Scholar]
- Bernard, G.R.; Artigas, A.; Brigham, K.L.; Carlet, J.; Falke, K.; Hudson, L.; Lamy, M.; Legall, J.R.; Morris, A.; Spragg, R. The american-european consensus conference on ards. Definitions, mechanisms, relevant outcomes, and clinical trial coordination. Am. J. Respir. Crit. Care Med 1994, 149, 818–824. [Google Scholar]
- Braude, S.; Nolop, K.B.; Hughes, J.M.; Barnes, P.J.; Royston, D. Comparison of lung vascular and epithelial permeability indices in the adult respiratory distress syndrome. Am. Rev. Respir. Dis 1986, 133, 1002–1005. [Google Scholar]
- Patterson, C.E.; Barnard, J.W.; Lafuze, J.E.; Hull, M.T.; Baldwin, S.J.; Rhoades, R.A. The role of activation of neutrophils and microvascular pressure in acute pulmonary edema. Am. Rev. Respir. Dis 1989, 140, 1052–1062. [Google Scholar]
- Tomashefski, J.F., Jr. Pulmonary pathology of the adult respiratory distress syndrome. Clin. Chest Med 1990, 11, 593–619. [Google Scholar]
- Singh, R.; Barden, A.; Mori, T.; Beilin, L. Advanced glycation end-products: A review. Diabetologia 2001, 44, 129–146. [Google Scholar]
- Wang, L.F.; Patel, M.; Razavi, H.M.; Weicker, S.; Joseph, M.G.; McCormack, D.G.; Mehta, S. Role of inducible nitric oxide synthase in pulmonary microvascular protein leak in murine sepsis. Am. J. Respir. Crit. Care Med 2002, 165, 1634–1639. [Google Scholar]
- Tate, R.M.; Repine, J.E. Neutrophils and the adult respiratory distress syndrome. Am. Rev. Respir. Dis 1983, 128, 552–559. [Google Scholar]
- Kindt, G.C.; Gadek, J.E.; Weiland, J.E. Initial recruitment of neutrophils to alveolar structures in acute lung injury. J. Appl. Physiol 1991, 70, 1575–1585. [Google Scholar]
- Doerschuk, C.M. Mechanisms of leukocyte sequestration in inflamed lungs. Microcirculation 2001, 8, 71–88. [Google Scholar]
- Brown, D.M.; Drost, E.; Donaldson, K.; MacNee, W. Deformability and cd11/cd18 expression of sequestered neutrophils in normal and inflamed lungs. Am. J. Respir. Cell Mol. Biol 1995, 13, 531–539. [Google Scholar]
- Skoutelis, A.T.; Kaleridis, V.; Athanassiou, G.M.; Kokkinis, K.I.; Missirlis, Y.F.; Bassaris, H.P. Neutrophil deformability in patients with sepsis, septic shock, and adult respiratory distress syndrome. Crit. Care Med 2000, 28, 2355–2359. [Google Scholar]
- Goode, H.F.; Webster, N.R. Free radicals and antioxidants in sepsis. Crit. Care Med 1993, 21, 1770–1776. [Google Scholar]
- Novelli, G.P. Role of free radicals in septic shock. J. Physiol. Pharmacol 1997, 48, 517–527. [Google Scholar]
- Razavi, H.M.; Werhun, R.; Scott, J.A.; Weicker, S.; Wang, L.F.; McCormack, D.G.; Mehta, S. Effects of inhaled nitric oxide in a mouse model of sepsis-induced acute lung injury. Crit. Care Med 2002, 30, 868–873. [Google Scholar]
- Hogg, J.C.; Doerschuk, C.M. Leukocyte traffic in the lung. Annu. Rev. Physiol 1995, 57, 97–114. [Google Scholar]
- Downey, G.P.; Fialkow, L.; Fukushima, T. Initial interaction of leukocytes within the microvasculature: Deformability, adhesion, and transmigration. New Horiz 1995, 3, 219–228. [Google Scholar]
- Razavi, H.M.; Wang, L.F.; Weicker, S.; Rohan, M.; Law, C.; McCormack, D.G.; Mehta, S. Pulmonary neutrophil infiltration in murine sepsis: Role of inducible nitric oxide synthase. Am. J. Respir. Crit. Care Med 2004, 170, 227–233. [Google Scholar]
- Aoki, Y.; Ota, M.; Katsuura, Y.; Komoriya, K.; Nakagaki, T. Effect of activated human protein c on disseminated intravascular coagulation induced by lipopolysaccharide in rats. Arzneimittelforschung 2000, 50, 809–815. [Google Scholar]
- Chiou, W.F.; Ko, H.C.; Chen, C.F.; Chou, C.J. Evodia rutaecarpa protects against circulation failure and organ dysfunction in endotoxaemic rats through modulating nitric oxide release. J. Pharm. Pharmacol 2002, 54, 1399–1405. [Google Scholar]
- Chen, C.P.; Yokozawa, T.; Kitani, K. Beneficial effects of sanguisorbae radix in renal dysfunction caused by endotoxin in vivo. Biol. Pharm. Bull 1999, 22, 1327–1330. [Google Scholar]
- Cunha, F.Q.; Assreuy, J.; Moncada, S.; Liew, F.Y. Phagocytosis and induction of nitric oxide synthase in murine macrophages. Immunology 1993, 79, 408–411. [Google Scholar]
- Deaciuc, I.V.; D’Souza, N.B.; de Villiers, W.J.; Burikhanov, R.; Sarphie, T.G.; Hill, D.B.; McClain, C.J. Inhibition of caspases in vivo protects the rat liver against alcohol-induced sensitization to bacterial lipopolysaccharide. Alcohol. Clin. Exp. Res 2001, 25, 935–943. [Google Scholar]
- Hong, K.W.; Kim, K.E.; Rhim, B.Y.; Lee, W.S.; Kim, C.D. Effect of rebamipide on liver damage and increased tumor necrosis factor in a rat model of endotoxin shock. Dig. Dis. Sci 1998, 43, 154S–159S. [Google Scholar]
- Jiang, J.; Chen, H.; Diao, Y.; Tian, K.; Zhu, P.; Wang, Z. Distribution of endotoxins in tissues and circulation and its effects following hemorrhagic shock. Chin. Med. J. (Engl. ) 1998, 111, 118–122. [Google Scholar]
- Barton, C.C.; Ganey, P.E.; Roth, R.A. Lipopolysaccharide augments aflatoxin B(1)-induced liver injury through neutrophil-dependent and -independent mechanisms. Toxicol. Sci 2000, 58, 208–215. [Google Scholar]
- Wellings, R.P.; Corder, R.; Vane, J.R. Lack of effect of ET antibody or SB 209670 on endotoxin-induced renal failure. J. Cardiovasc. Pharmacol 1995, 26, S476–S478. [Google Scholar]
- Memon, R.A.; Grunfeld, C.; Moser, A.H.; Feingold, K.R. Tumor necrosis factor mediates the effects of endotoxin on cholesterol and triglyceride metabolism in mice. Endocrinology 1993, 132, 2246–2253. [Google Scholar]
- Kassim, M.; Mansor, M.; Achoui, M.; Ong, G.S.Y.; Sekaran, S.D.; Yusoff, K.M. Honey as an immunomodulator during sepsis in animal models. Crit. Care 2009, 13. [Google Scholar] [CrossRef]
- Salah, N.; Miller, N.J.; Paganga, G.; Tijburg, L.; Bolwell, G.P.; Rice-Evans, C. Polyphenolic flavanols as scavengers of aqueous phase radicals and as chain-breaking antioxidants. Arch. Biochem. Biophys 1995, 322, 339–346. [Google Scholar]
- Unno, T.; Sakane, I.; Masumizu, T.; Kohno, M.; Kakuda, T. Antioxidant activity of water extracts of lagerstroemia speciosa leaves. Biosci. Biotechnol. Biochem 1997, 61, 1772–1774. [Google Scholar]
- Virgili, F.; Kim, D.; Packer, L. Procyanidins extracted from pine bark protect alpha-tocopherol in ECV 304 endothelial cells challenged by activated raw 264.7 macrophages: Role of nitric oxide and peroxynitrite. Fed. Eur. Biochem. Soc. Lett 1998, 431, 315–318. [Google Scholar]
- Yang, F.; de Villiers, W.J.; McClain, C.J.; Varilek, G.W. Green tea polyphenols block endotoxin-induced tumor necrosis factor-production and lethality in a murine model. J. Nutr 1998, 128, 2334–2340. [Google Scholar]
- Wakabayashi, I. Inhibitory effects of baicalein and wogonin on lipopolysaccharide-induced nitric oxide production in macrophages. Basic Clin. Pharmacol. Toxicol 1999, 84, 288–291. [Google Scholar]
- Bauer, L.; Kohlich, A.; Hirschwehr, R.; Siemann, U.; Ebner, H.; Scheiner, O.; Kraft, D.; Ebner, C. Food allergy to honey: Pollen or bee products? Characterization of allergenic proteins in honey by means of immunoblotting. J. Allergy Clin. Immunol 1996, 97, 65–73. [Google Scholar]
- Bradley, P.P.; Christensen, R.D.; Rothstein, G. Cellular and extracellular myeloperoxidase in pyogenic inflammation. Blood 1982, 60, 618–622. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
| Parameter | Normal Rabbits (N = 6) | Untreated (N = 6) | Honey-treated (N = 6) |
|---|---|---|---|
| Urea (mmol/L) | 5.85 ± 0.20 | 55.85 ± 2.5 | 10.5 ± 0.23 a |
| Creatinine (mmol/L) | 83.71 ± 2.5 | 154 ± 6.16 | 72 ± 2.87 a |
| ALT (IU/L) | 54.125 ± 1.8 | 108.75 ± 3.6 | 78.75 ± 1.98 a |
| AST (IU/L) | 27.75 ± 0.9 | 577.33 ± 19.2 | 231.5 ± 7.6 a |
| ALP (IU/L) | 131.5 ± 4.1 | 542.75 ± 15.5 | 308.75 ± 11.4 a |
| GGT (IU/L) | 10 ± 0.32 | 38.4 ± 0.6 | 25 ± 1.7 b |
| Triglyceride (mmol/L) | 0.885 ± 0.03 | 10.434 ± 0.4 | 2.47 ± 0.13 a |
| Total cholesterol (mmol/L) | 1.1125 ± 0.04 | 3.16 ± 0.1 | 1.8 ± 0.08 c |
| HDL (mmol/L) | 0.65 ± 0.03 | 0.25 ± 0.006 | 0.545 ± 0.02 b |
| LDL (mmol/L) | 0.202 ± 0.005 | 0.45 ± 0.01 | 0.32 ± 0.012 |
| Creatine kinase (IU/L) | 1327.4 ± 5.3 | 2168.3 ± 34 | 998.6 ± 26.8 a |
| pH (KPa) | 7.38 ± 0.3 | 7.36 ± 0.3 | 7.5 ± 0.27 b |
| pCO2 (KPa) | 4.3 ± 0.17 | 3 ± 0.12 | 3 ± 0.12 |
| pO2 (KPa) | 16.21 ± 0.53 | 7.65 ± 0.23 | 19.3 ± 0 .77 b |
| HCO3 (mmol/L) | 19 ± 0.71 | 15 ± 0.51 | 19 ± 0.54 a |
| Platelet (10e9/L) | 194.3 ± 6.4 | 144.5 ± 4.3 | 183.4 ± 7.6 b |
| Amylase (IU/L) | 181.3 ± 7.2 | 215.5 ± 6.2 | 180 ± 5.2 b |
| RBC (10e12/L | 9.27 ± 0.31 | 4.83 ± 0.15 | 8.3475 ± 0.32 a |
| WBC (10e9/L) | 15 ± 0.6 | 6.05 ± 0.25 | 11.65 ± 0.36 a |
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Kassim, M.; Mansor, M.; Al-Abd, N.; Yusoff, K.M. Gelam Honey Has a Protective Effect against Lipopolysaccharide (LPS)-Induced Organ Failure. Int. J. Mol. Sci. 2012, 13, 6370-6381. https://doi.org/10.3390/ijms13056370
Kassim M, Mansor M, Al-Abd N, Yusoff KM. Gelam Honey Has a Protective Effect against Lipopolysaccharide (LPS)-Induced Organ Failure. International Journal of Molecular Sciences. 2012; 13(5):6370-6381. https://doi.org/10.3390/ijms13056370
Chicago/Turabian StyleKassim, Mustafa, Marzida Mansor, Nazeh Al-Abd, and Kamaruddin Mohd Yusoff. 2012. "Gelam Honey Has a Protective Effect against Lipopolysaccharide (LPS)-Induced Organ Failure" International Journal of Molecular Sciences 13, no. 5: 6370-6381. https://doi.org/10.3390/ijms13056370