Pokemon Silencing Leads to Bim-Mediated Anoikis of Human Hepatoma Cell QGY7703

Abstract
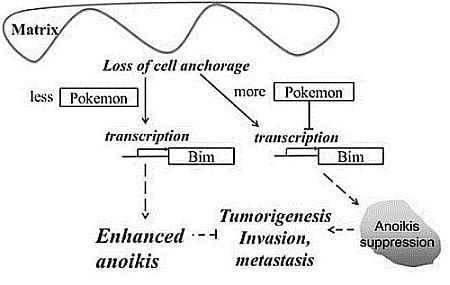
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results and Discussion
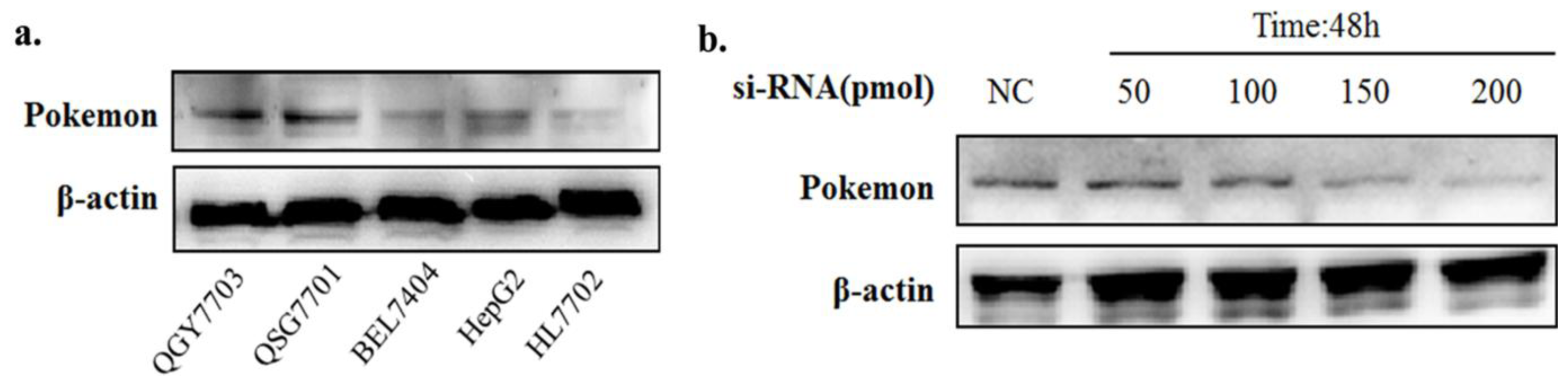
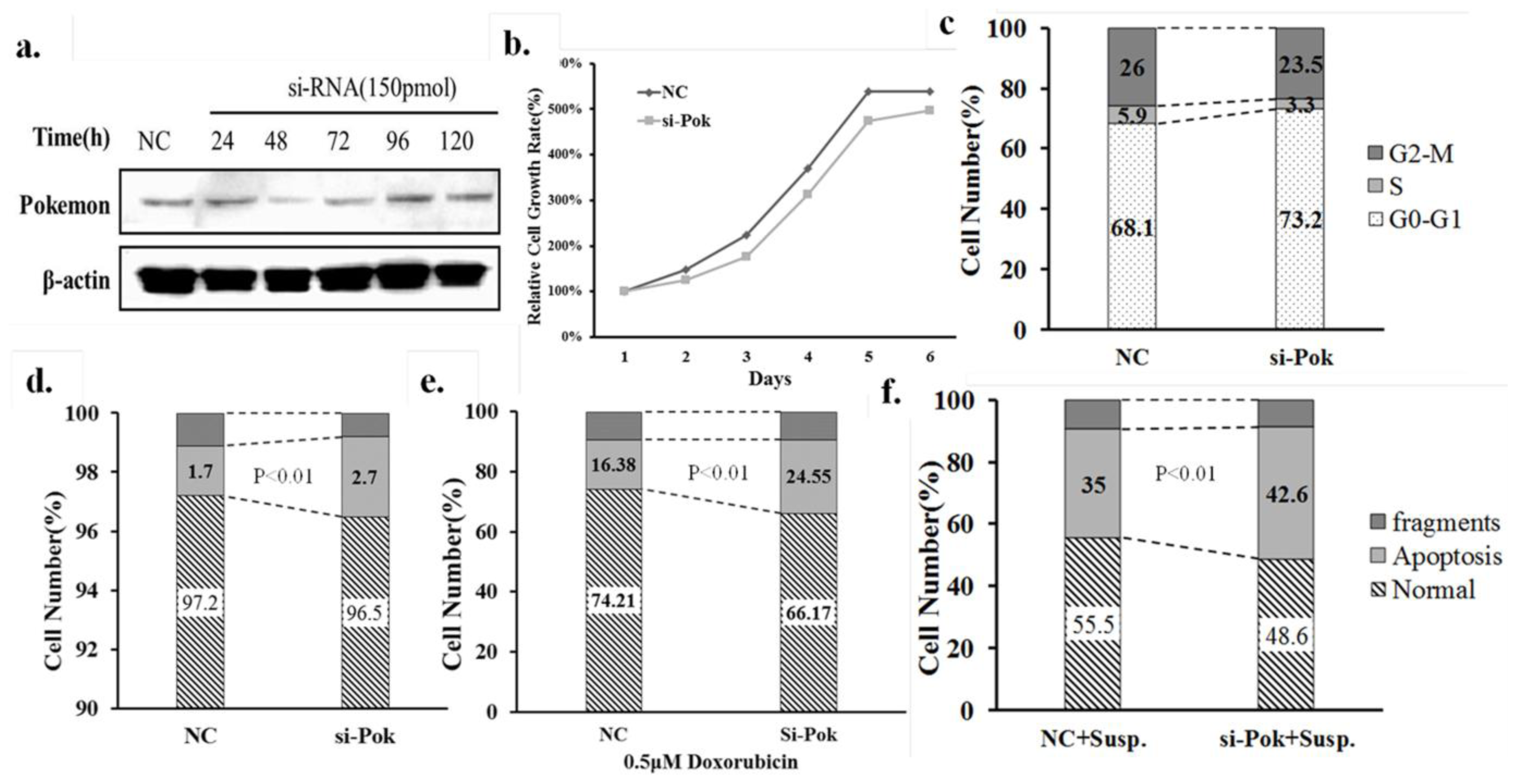
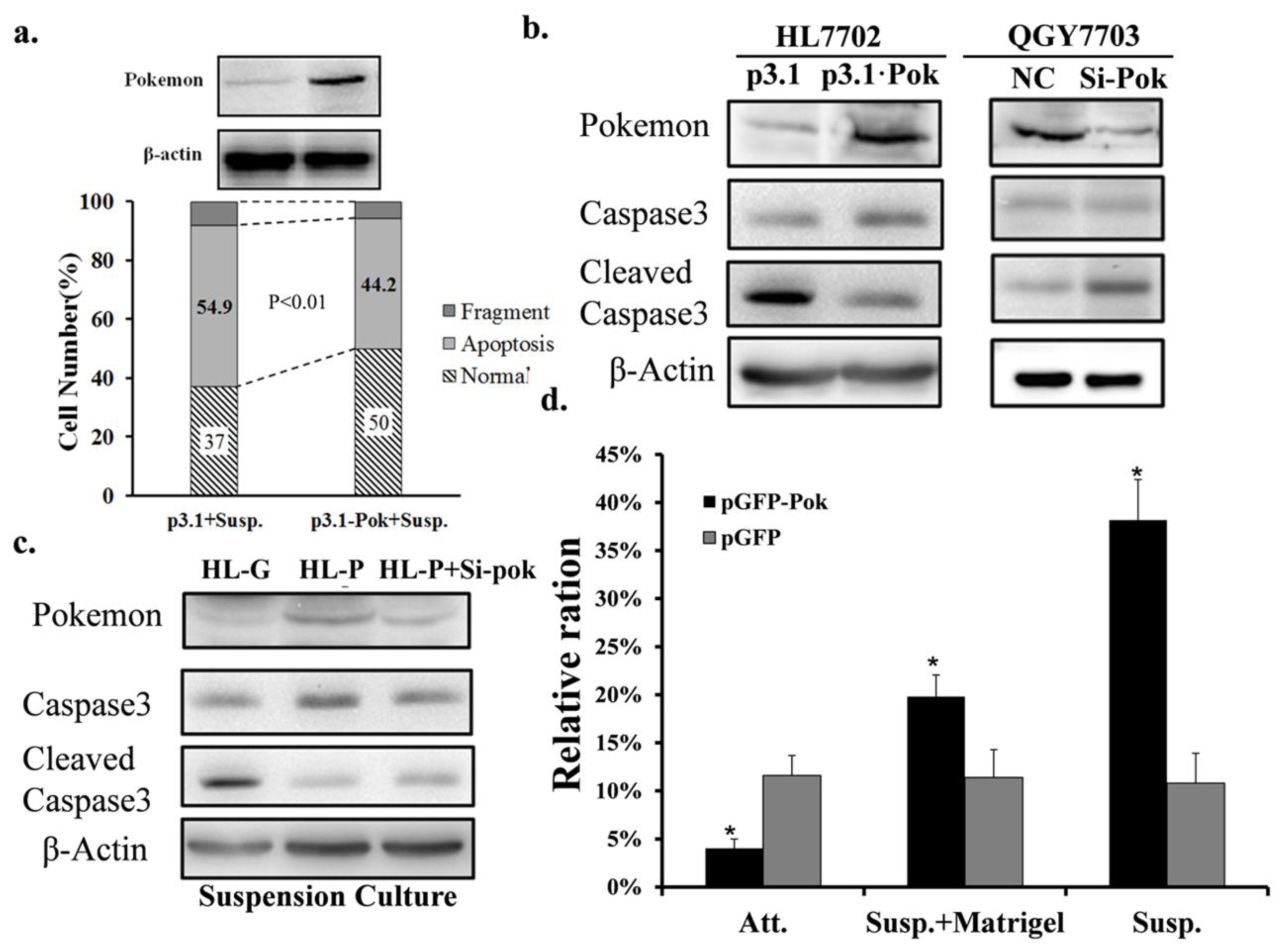
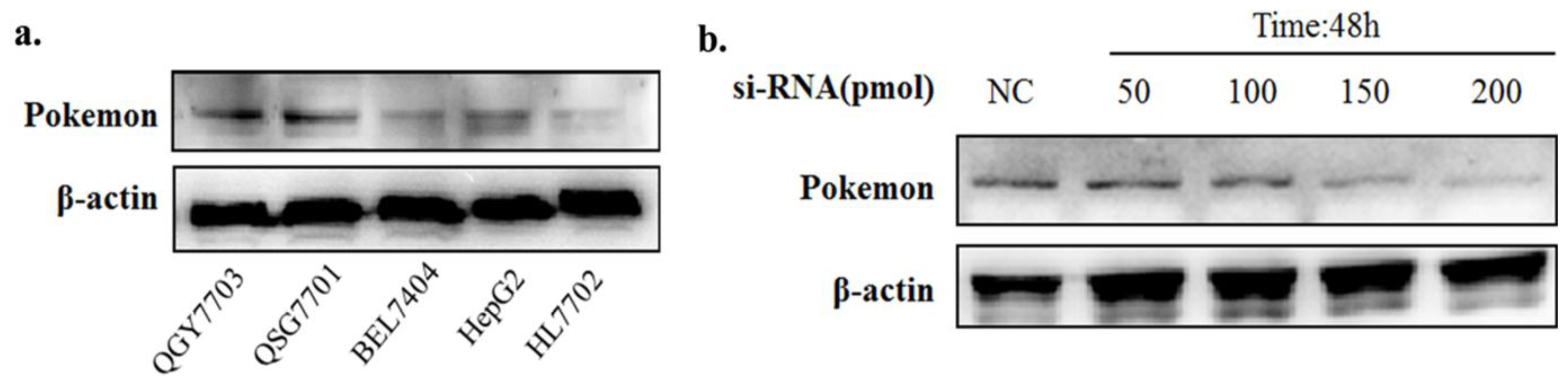
2.1. Pokemon-Silencing Sensitizes Human Hepatoma to Anoikis and an Apoptosis Inducer
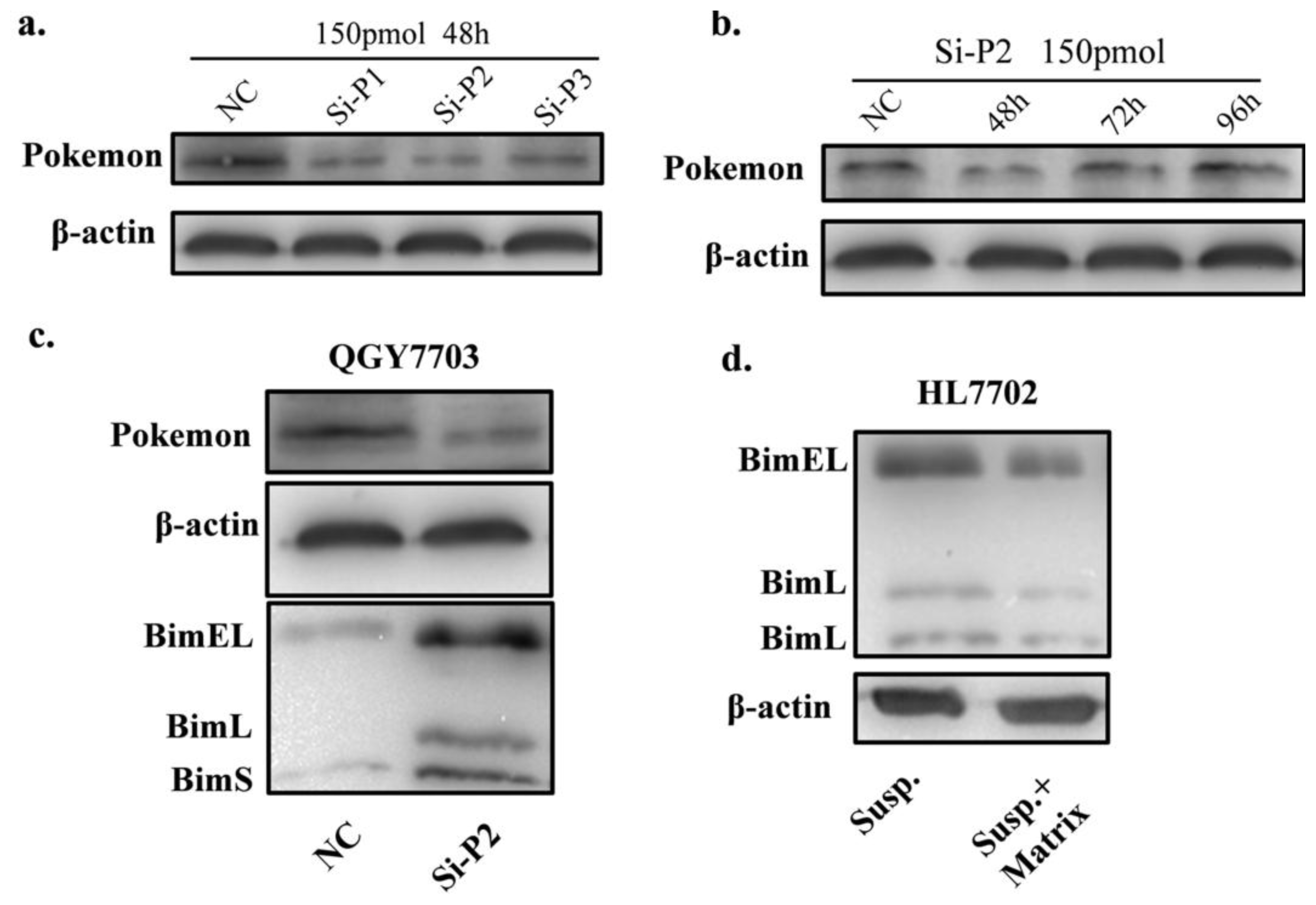
2.2. Pokemon Enhances Resistance of HL7702 Cells to Anoikis
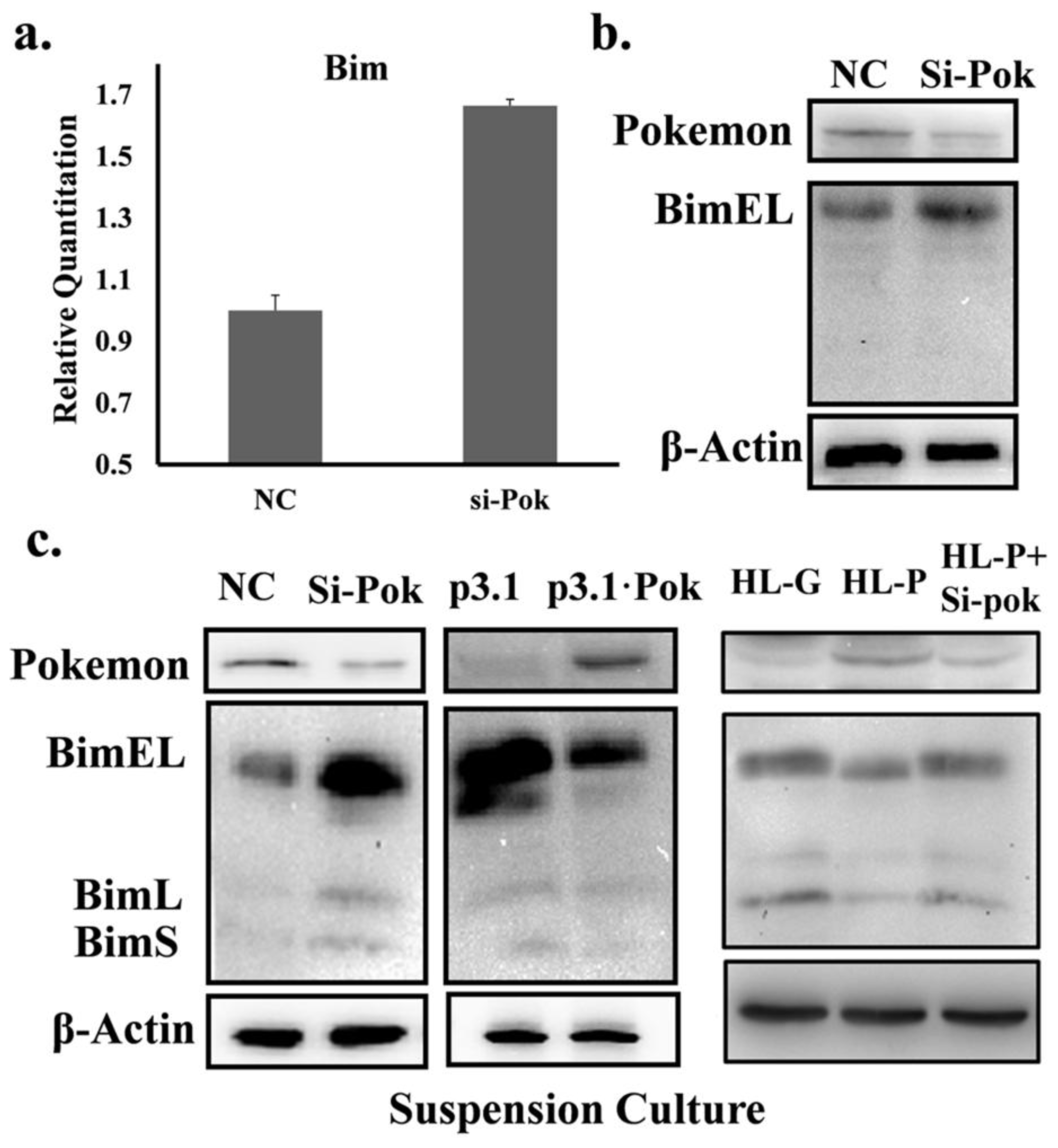
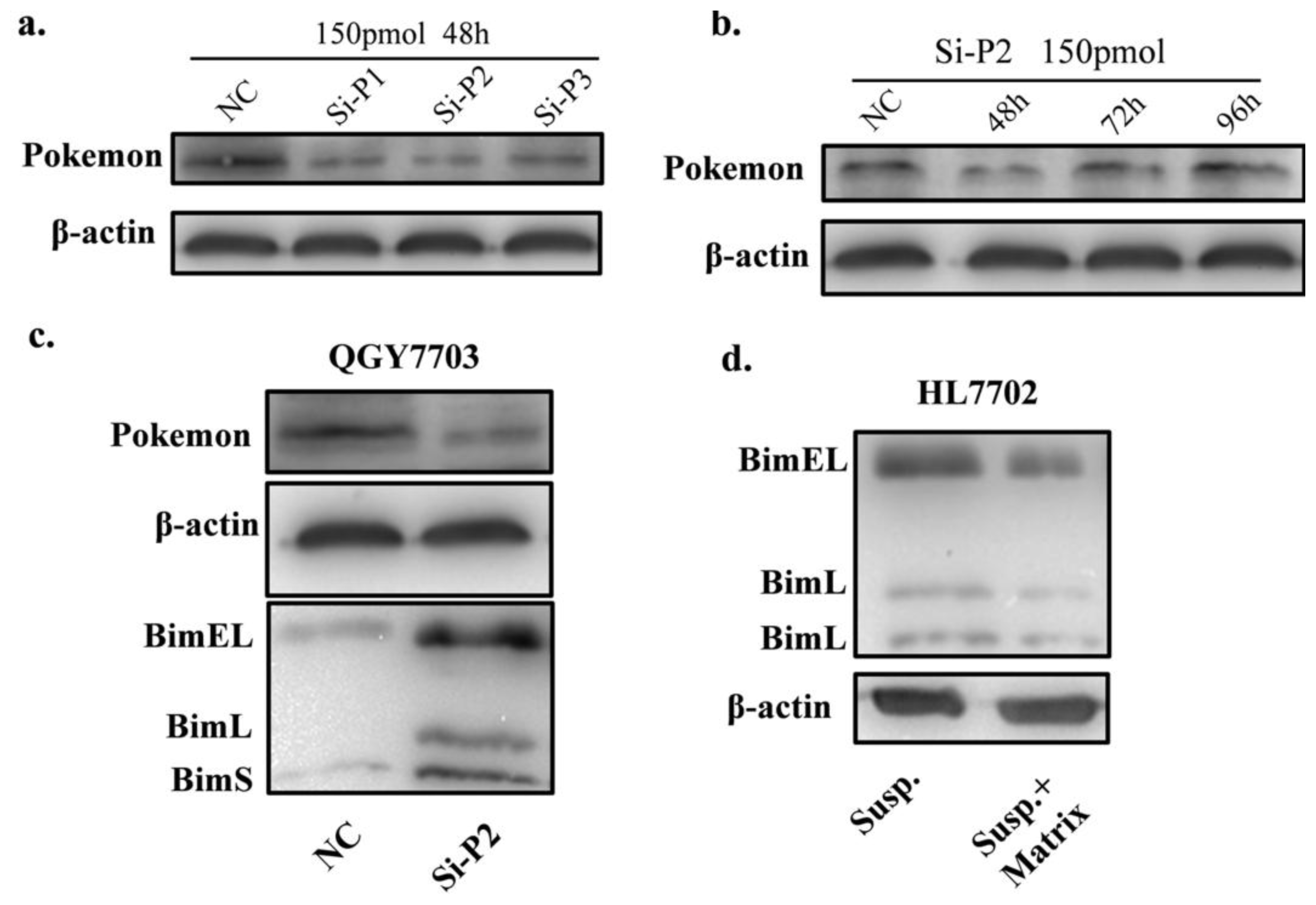
2.3. Pokemon Prevents Anoikis through Suppression of Bim Expression
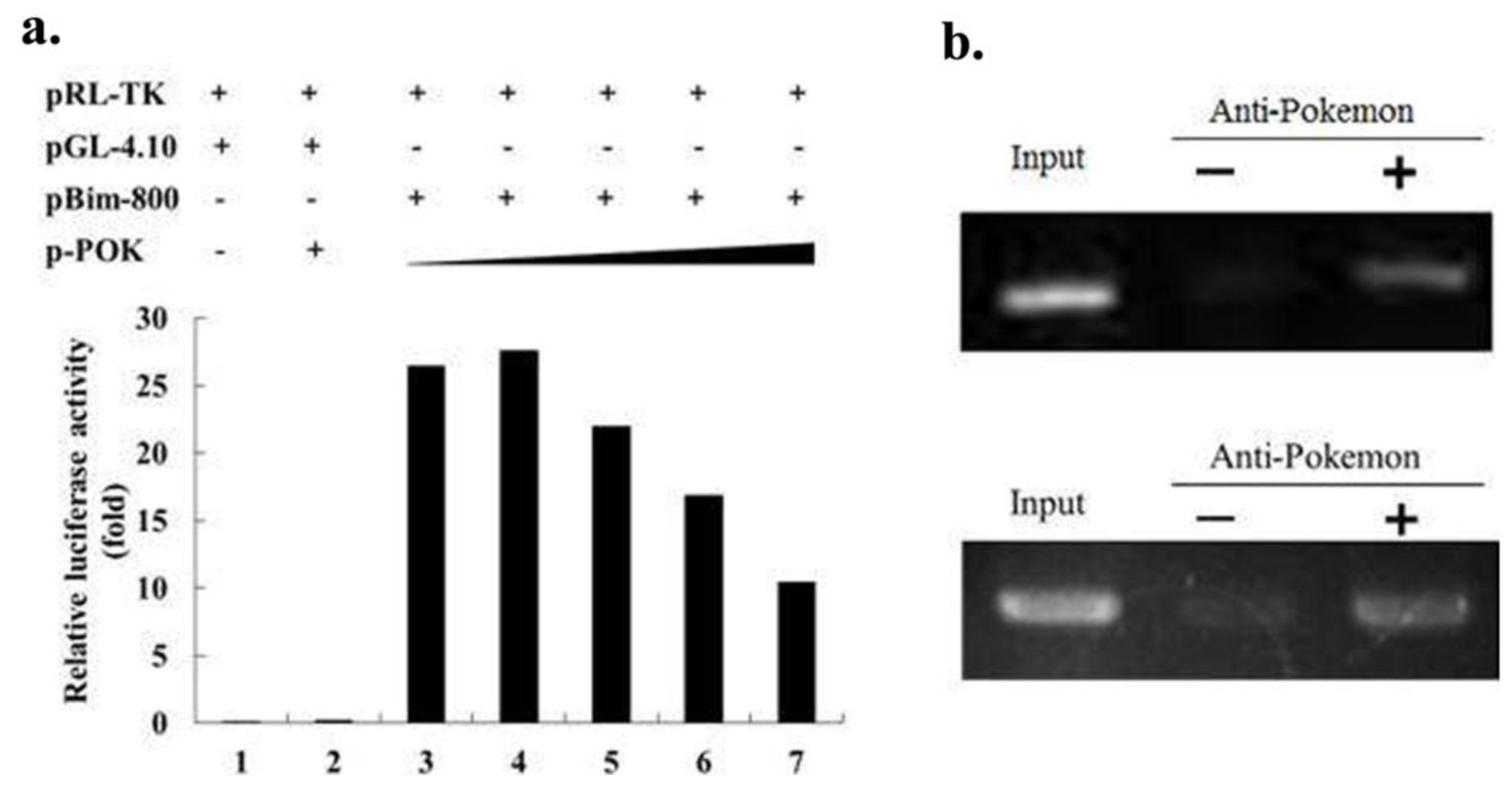
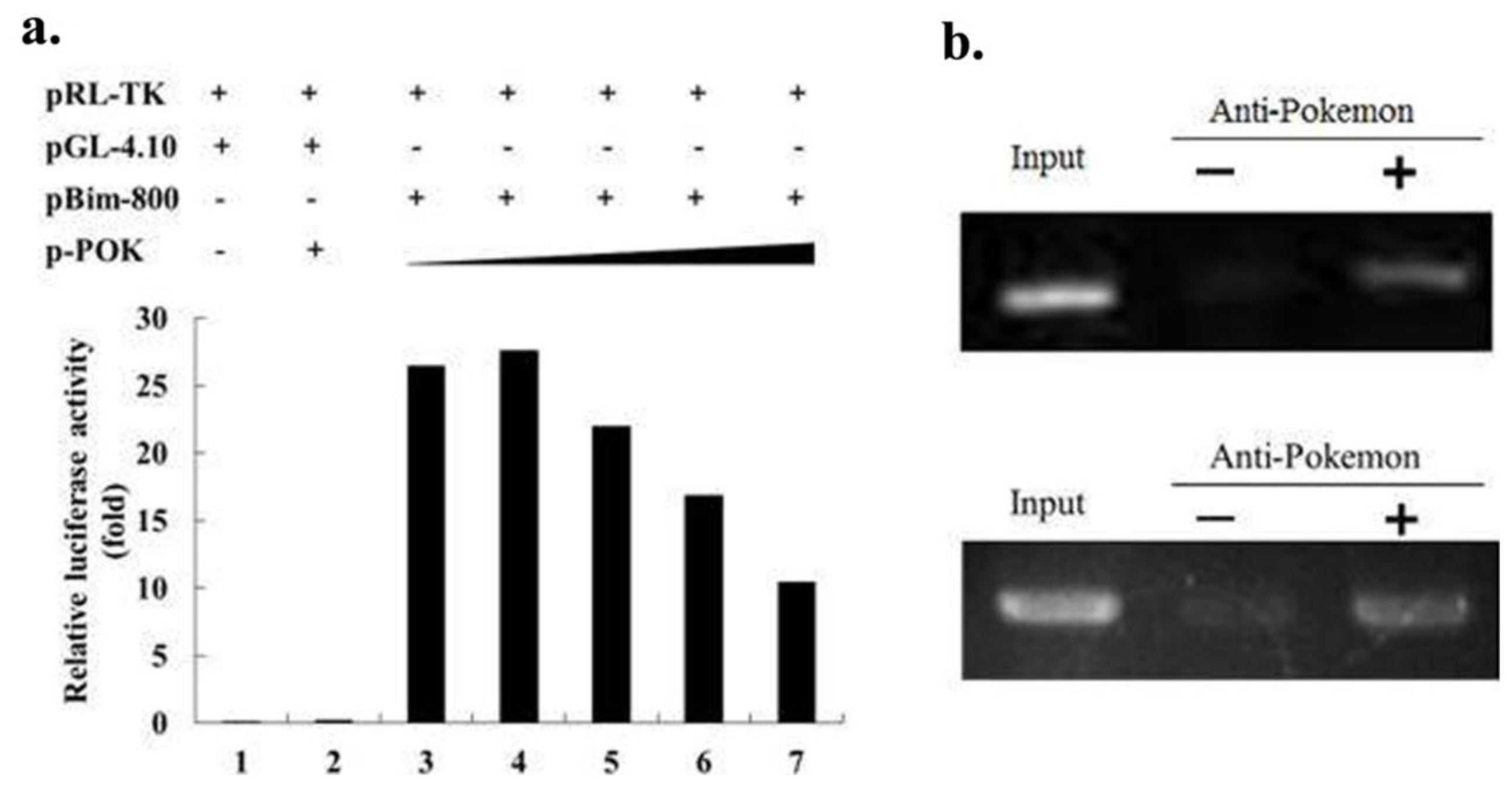
2.4. Pokemon Suppresses Bim Expression via Direct Binding to the Bim Promoter
2.5. Discussion
3. Experimental Section
3.1. Cell Lines and Culture
3.2. Plasmid Construction
3.3. Pokemon Silencing by siRNA
3.4. Flow Cytometry Analysis
3.5. Real-Time PCR
3.6. Western Blot
3.7. Chromatin Immunoprecipitation
3.8. Dual Luciferase Reporter Assay
3.9. Cell Counting Assay
4. Conclusions
Acknowledgments
- Conflict of InterestThe authors declare no conflict of interest.
References
- Morrison, D.J.; Pendergrast, P.S.; Stavropoulos, P.; Colmenares, S.U.; Kobayashi, R.; Hernandez, N. FBI-1, a factor that binds to the HIV-1 inducer of short transcripts (IST), is a POZ domain protein. Nucleic Acids Res 1999, 27, 1251–1252. [Google Scholar]
- Maeda, T.; Merghoub, T.; Hobbs, R.M.; Dong, L.; Maeda, M.; Zakrzewski, J.; van den Brink, M.R.M.; Zelent, A.; Shigematsu, H.; Akashi, K.; et al. Regulation of B versus T lymphoid lineage fate decision by the proto-oncogene LRF. Science 2007, 316, 860–866. [Google Scholar]
- Maeda, T.; Ito, K.; Merghoub, T.; Poliseno, L.; Hobbs, R.M.; Wang, G.; Dong, L.; Maeda, M.; Dore, L.C.; Zelent, A.; et al. LRF is an essential downstream target of GATA1 in erythroid development and regulates BIM-dependent apoptosis. Dev. Cell 2009, 17, 527–540. [Google Scholar]
- Ji, Y.; He, X.S.; Ma, Y.; Zhu, X.F.; Zhang, R.; Wu, J. Expression of Pokemon in HCC, HCC with distant place metastasis, hepatocirrhosis and normal crowds. Chin. J. Exp. Surg 2007, 24, 75–79. [Google Scholar]
- Rovin, R.A.; Winn, R. Pokemon expression in malignant glioma: An application of bioinformatics methods. Neurosurg. Focus 2005, 19, 1–2. [Google Scholar]
- Aggarwal, A.; Hunter, W.J.; Aggarwal, H.; Silva, E.D.; Davey, M.S.; Murphy, R.F.; Agrawal, D.K. Expression of leukemia/lymphoma-related factor (LRF/POKEMON) in human breast carcinoma and other cancers. Exp. Mol. Pathol 2010, 89, 140–148. [Google Scholar]
- Maeda, T.; Hobbs, R.M.; Merghoub, T.; Guernah, I.; Arthur, Z.; Cordon-Cardo, C.; Teruya-Feldstein, J.; Pandolfi, P.P. Role of the proto-oncogene Pokemon in cellular transformation and ARF repression. Nature 2005, 433, 278–285. [Google Scholar]
- Maeda, T.; Hobbs, R.M.; Pandolfi, P.P. The transcription factor Pokemon: A new key player in cancer pathogenesis. Cancer Res 2005, 65, 8575. [Google Scholar]
- El-Deiry, W.S.; Tokinom, T.; Velculescu, V.E.; Levy, D.B.; Parsons, R.; Trent, J.M.; Lin, D.; Mercer, W.E.; Kinzler, K.W.; Vogelstein, B. WAF1, a potential mediator of p53 tumor suppression. Cell 1993, 75, 817–825. [Google Scholar]
- El-Deiry, W.S.; Harper, J.W.; O’Connor, P.M.; Velculescu, V.E.; Canman, C.E.; Jackman, J.; Pietenpol, J.A.; Burrell, M.; Hill, D.E.; Wang, Y.; et al. WAF1/CIP1 is induced in p53-mediated G1 arrest and apoptosis. Cancer Res 1994, 54, 1169–1174. [Google Scholar]
- Ezhevsky, S.A.; Ho, A.; Becker-Hapak, M.; Davis, P.K.; Dowdy, S.F. Differential regulation of retinoblastoma tumor suppressor protein by G1 cyclin-dependent kinase complexes in vivo. Mol. Cell. Biol 2001, 21, 4773–4784. [Google Scholar]
- Goodrich, D.W.; Wang, N.P.; Qian, Y.W.; Lee, E.Y.; Lee, W.H. The retinoblastoma gene product regulates progression through the G1 phase of the cell cycle. Cell 1981, 67, 293–302. [Google Scholar]
- Choi, W.I.; Jeon, B.N.; Yun, C.O.; Kim, P.H.; Kim, S.E.; Choi, K.Y.; Kim, S.H.; Hur, M.W. Proto-oncogene FBI-1 represses transcription of p21CIP1 by inhibition of transcription activation by p53 and Sp1. J. Biol. Chem 2009, 284, 12633–12644. [Google Scholar]
- Jeon, B.N.; Yoo, J.Y.; Choi, W.I.; Lee, C.E.; Yoon, H.G.; Hur, M.W. Proto-oncogene FBI-1 (Pokemon/ZBTB7A) represses transcription of the tumor suppressor Rb Gene via binding competition with Sp1 and recruitment of co-repressors. J. Biol. Chem 2008, 283, 33199–33210. [Google Scholar]
- Choi, W.I.; Jeon, B.N.; Park, H.; Yoo, J.Y.; Kim, Y.S.; Koh, D.I.; Kim, M.H.; Kim, Y.R.; Lee, C.E.; Kim, K.S.; et al. Proto-oncogene FBI-1 (Pokemon) and SREBP-1 synergistically activate transcription of fatty-acid synthase gene (FASN). J. Biol. Chem 2008, 283, 29341–29354. [Google Scholar]
- Kuhajda, F.P. Fatty acid synthase and cancer: New application of an old pathway. Cancer Res 2006, 66, 5977–5980. [Google Scholar]
- Bouillet, P.; Zhang, L.C.; Huang, D.C.S.; Webb, G.C.; Bottema, C.D.K.; Shore, P.; Eyre, H.J.; Sutherland, G.R.; Adams, J.M. Gene structure, alternative splicing, and chromosomal localization of pro-apoptotic Bcl-2 relative Bim. Mamm. Genome 2001, 12, 163–168. [Google Scholar]
- Adams, J.M.; Cory, S. The Bcl-2 protein family: Arbiters of cell survival. Science 1998, 281, 1322–1326. [Google Scholar]
- Gilmore, A.P.; Metcalfe, A.D.; Romer, L.H.; Streuli, C.H. Integrin-mediated survival signals regulate the apoptotic function of Bax through its conformation and subcellular localization. J. Cell. Biochem 2000, 149, 431–446. [Google Scholar]
- Lindsten, T.; Ross, A.J.; King, A.; Zong, W.X.; Rathmell, J.C.; Shiels, H.A.; Ulrich, E.; Waymire, K.G.; Mahar, P.; Frauwirth, K. The combined functions of proapoptotic Bcl-2 family members bak and bax are essential for normal development of multiple tissues. Mol. Cell 2000, 6, 1389–1399. [Google Scholar]
- Schwartz, M.A. Integrins, oncogenes, and anchorage independence. J. Cell. Biochem 1997, 139, 575–578. [Google Scholar]
- Chiarugi, P.; Giannoni, E. Anoikis: A necessary death program for anchorage-dependent cells. Biochem. Pharmacol 2008, 76, 1352–1364. [Google Scholar]
- Frisch, S.M.; Ruoslahti, E. Integrins and anoikis. Curr. Opin. Cell Biol 1997, 9, 701–706. [Google Scholar]
- Li, Z.; Zhao, J.; Du, Y.; Park, H.R.; Sun, S.Y.; Bernal-Mizrachi, L.; Aitken, A.; Khuri, F.R.; Fu, H. Down-regulation of 14-3-3ζ suppresses anchorage-independent growth of lung cancer cells through anoikis activation. Proc. Natl. Acad. Sci. USA 2008, 105, 162–167. [Google Scholar]
- Gan, L.; Liu, P.; Lu, H.; Chen, S.; Yang, J.; McCarthy, J.; Knudsen, K.; Huang, H. Cyclin D1 promotes anchorage-independent cell survival by inhibiting FOXO-mediated anoikis. Cell Death Differ 2009, 16, 1408–1417. [Google Scholar]
- Woods, N.T.; Yamaguchi, H.; Lee, F.Y.; Bhalla, K.N.; Wang, H.G. Anoikis, initiated by Mcl-1 degradation and Bim induction, is deregulated during oncogenesis. Cancer Res 2007, 67, 10744–10752. [Google Scholar]
- Frisch, S.M.; Screaton, R.A. Anoikis mechanisms. Curr. Opin. Cell Biol 2001, 13, 555–562. [Google Scholar]
- Hanahan, D. The hallmarks of cancer. Cell 2000, 100, 57–70. [Google Scholar]
- Zu, X.; Ma, J.; Liu, H.; Liu, F.; Tan, C.; Yu, L.; Wang, J.; Xie, Z.; Cao, D.; Jiang, Y. Pro-oncogene Pokemon promotes breast cancer progression by upregulating survivin expression. Breast Cancer Res 2011, 13. [Google Scholar] [CrossRef]
- Parkin, D.M.; Pisani, P.; Ferlay, J. Global cancer statistics. CA Cancer J. Clin 1999, 49, 33–64. [Google Scholar]
- Ku, S.C.Y.; Ng, D.T.W.; Yap, M.G.S.; Chao, S.H. Effects of overexpression of X-box binding protein 1 on recombinant protein production in Chinese hamster ovary and NS0 myeloma cells. Biotechnol. Bioeng 2008, 99, 155–164. [Google Scholar]
- He, S.; Liu, F.; Xie, Z.; Zu, X.; Xu, W.; Jiang, Y. P-Glycoprotein/MDR1 regulates Pokemon gene transcription through p53 expression in human breast cancer cells. Int. J. Mol. Sci 2010, 11, 3039–3051. [Google Scholar]



© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Liu, K.; Liu, F.; Zhang, N.; Liu, S.; Jiang, Y. Pokemon Silencing Leads to Bim-Mediated Anoikis of Human Hepatoma Cell QGY7703. Int. J. Mol. Sci. 2012, 13, 5818-5831. https://doi.org/10.3390/ijms13055818
Liu K, Liu F, Zhang N, Liu S, Jiang Y. Pokemon Silencing Leads to Bim-Mediated Anoikis of Human Hepatoma Cell QGY7703. International Journal of Molecular Sciences. 2012; 13(5):5818-5831. https://doi.org/10.3390/ijms13055818
Chicago/Turabian StyleLiu, Kun, Feng Liu, Nannan Zhang, Shiying Liu, and Yuyang Jiang. 2012. "Pokemon Silencing Leads to Bim-Mediated Anoikis of Human Hepatoma Cell QGY7703" International Journal of Molecular Sciences 13, no. 5: 5818-5831. https://doi.org/10.3390/ijms13055818