In Vitro Antioxidant and Anti-Proliferation Activities of Polysaccharides from Various Extracts of Different Mushrooms



Abstract
:1. Introduction
2. Results and Discussion
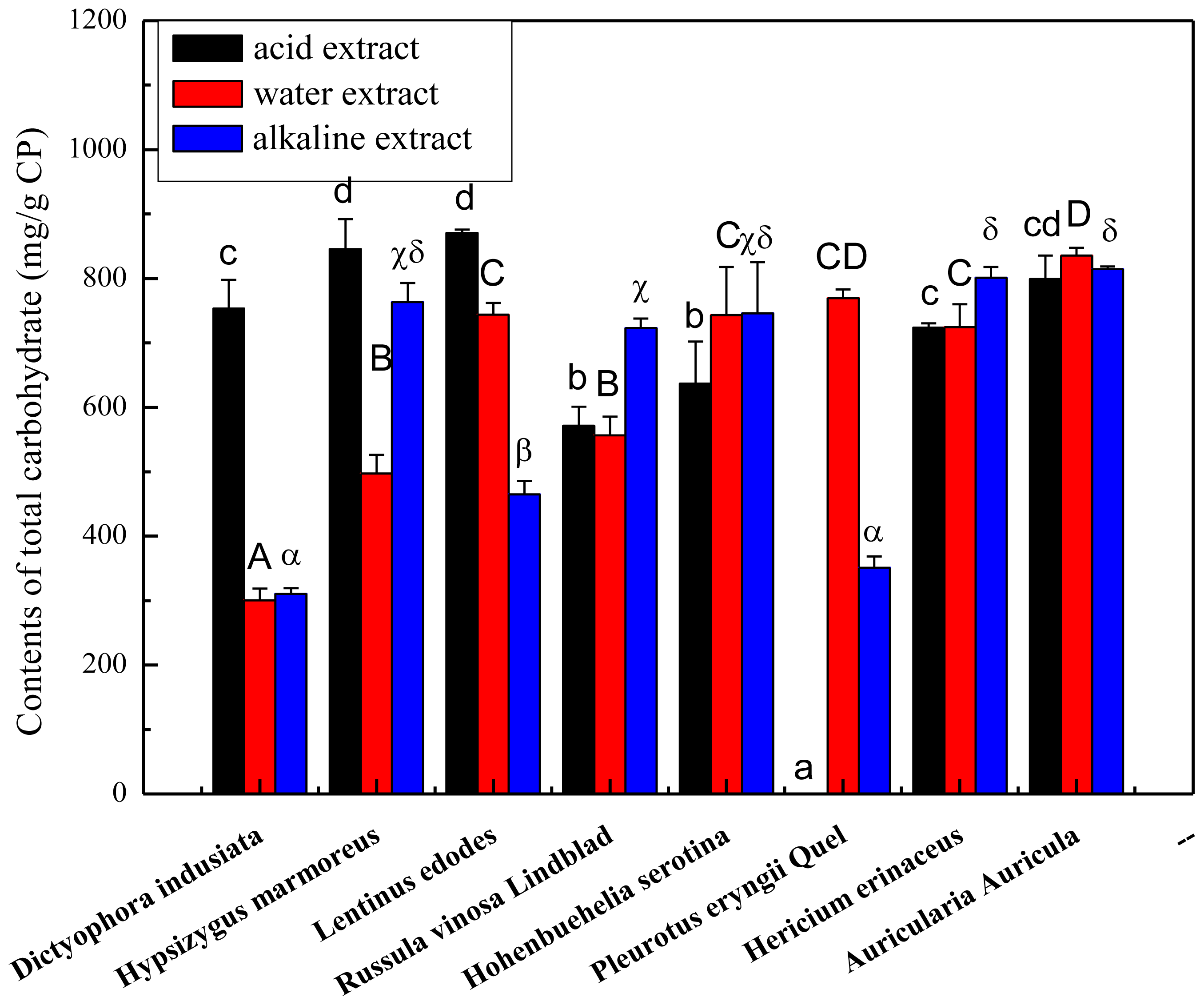
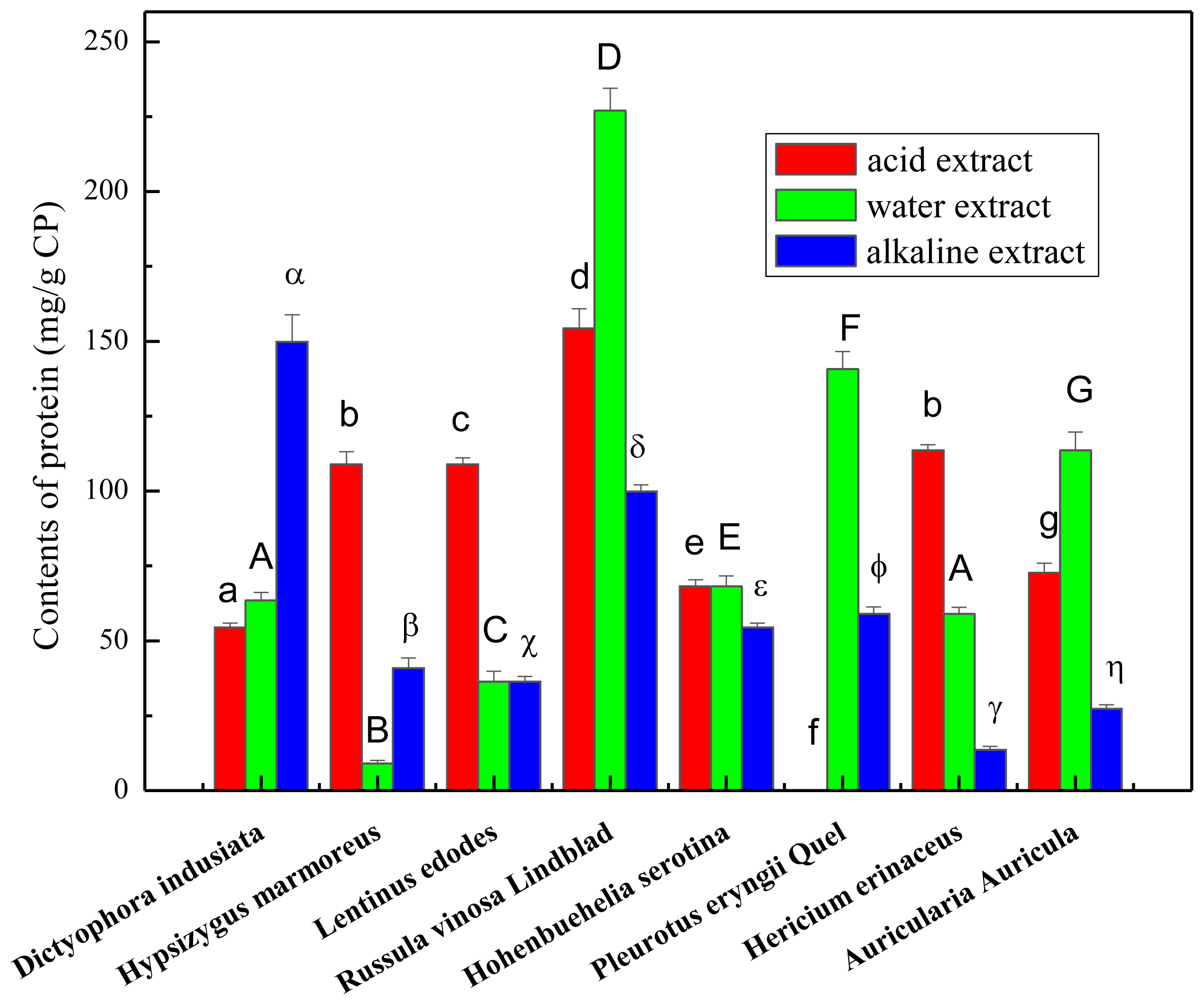
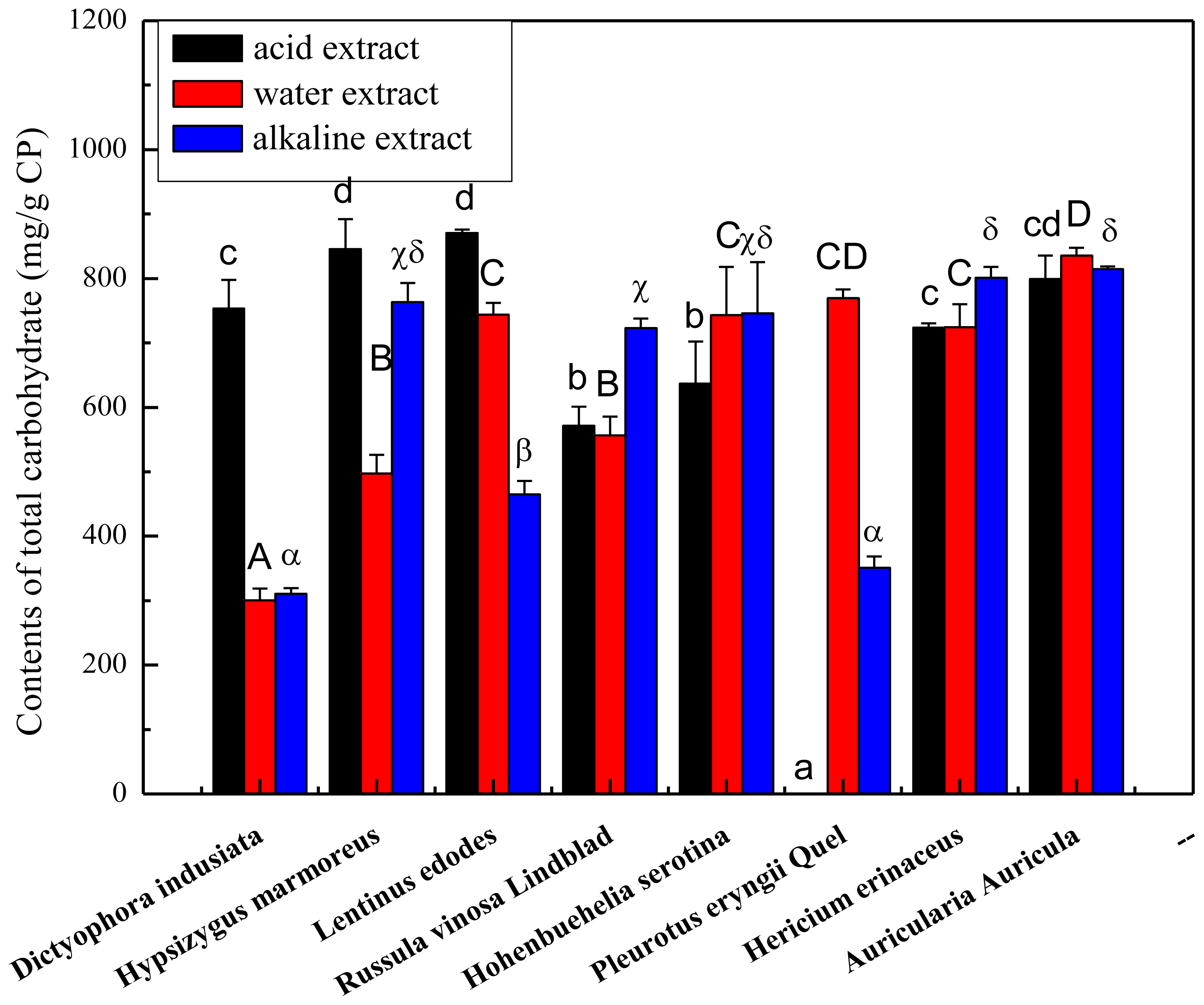
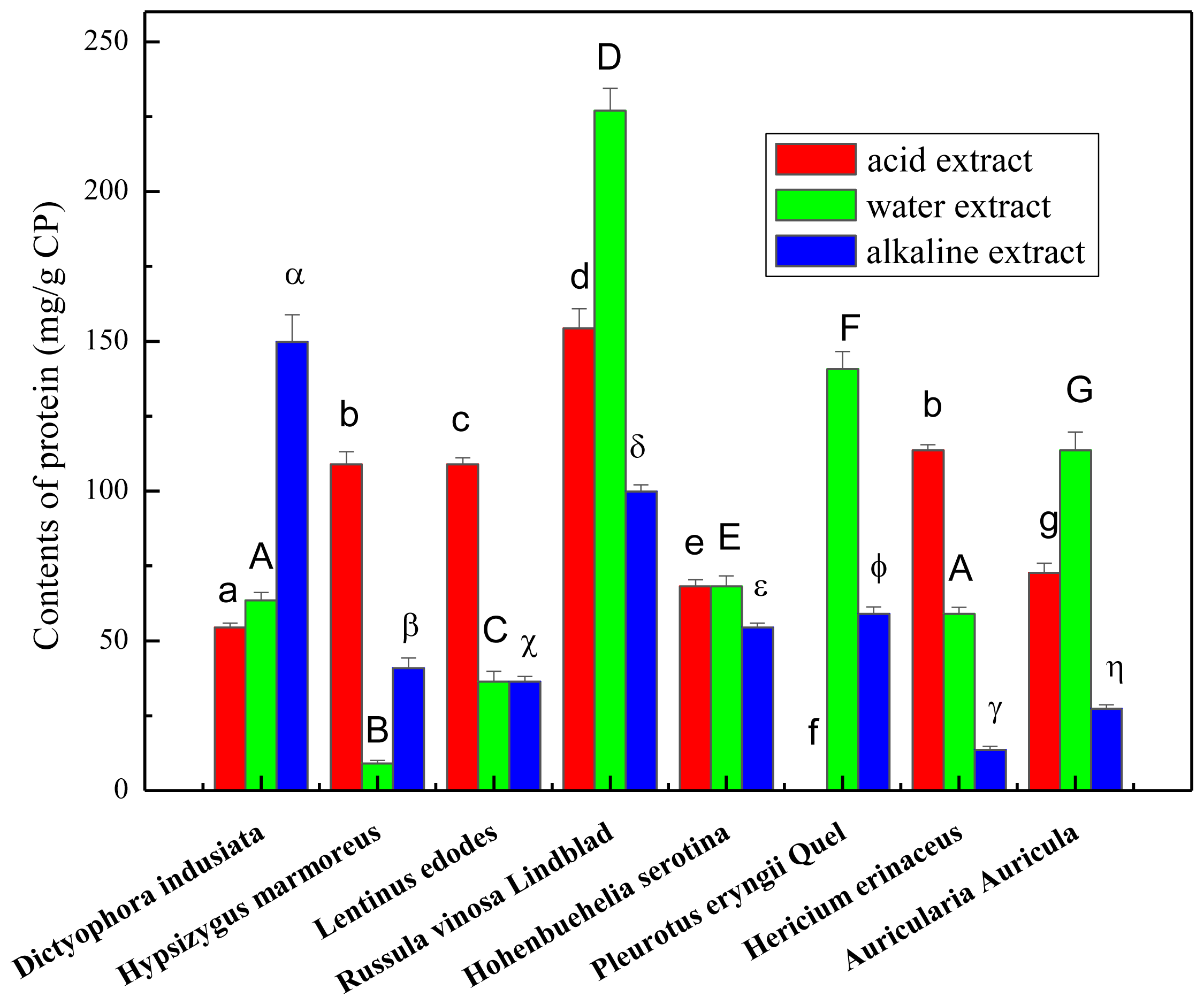
2.1. Total Carbohydrate, Phenolics and Protein Content of the Crude Polysaccharides
2.2. Antioxidant Activity
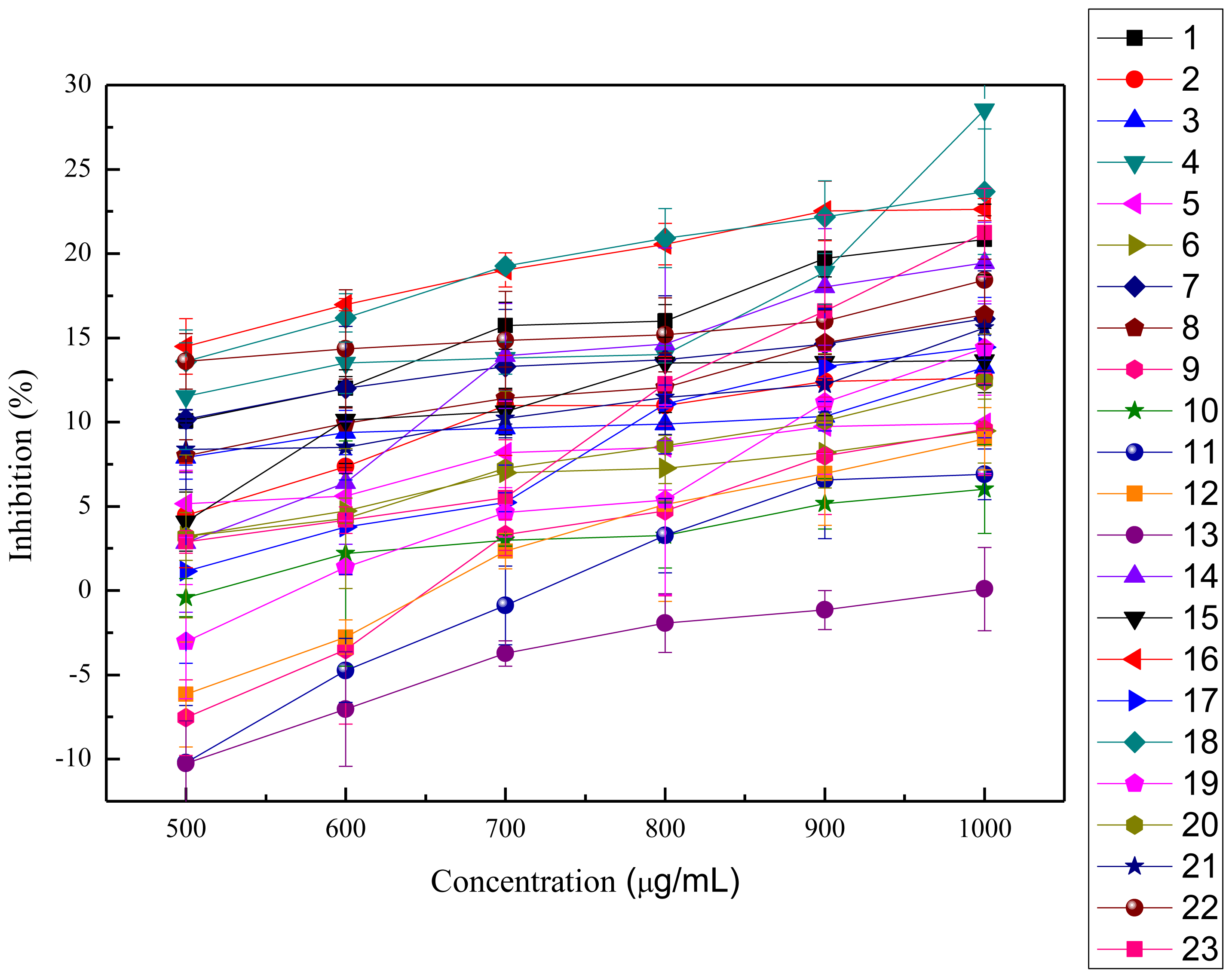
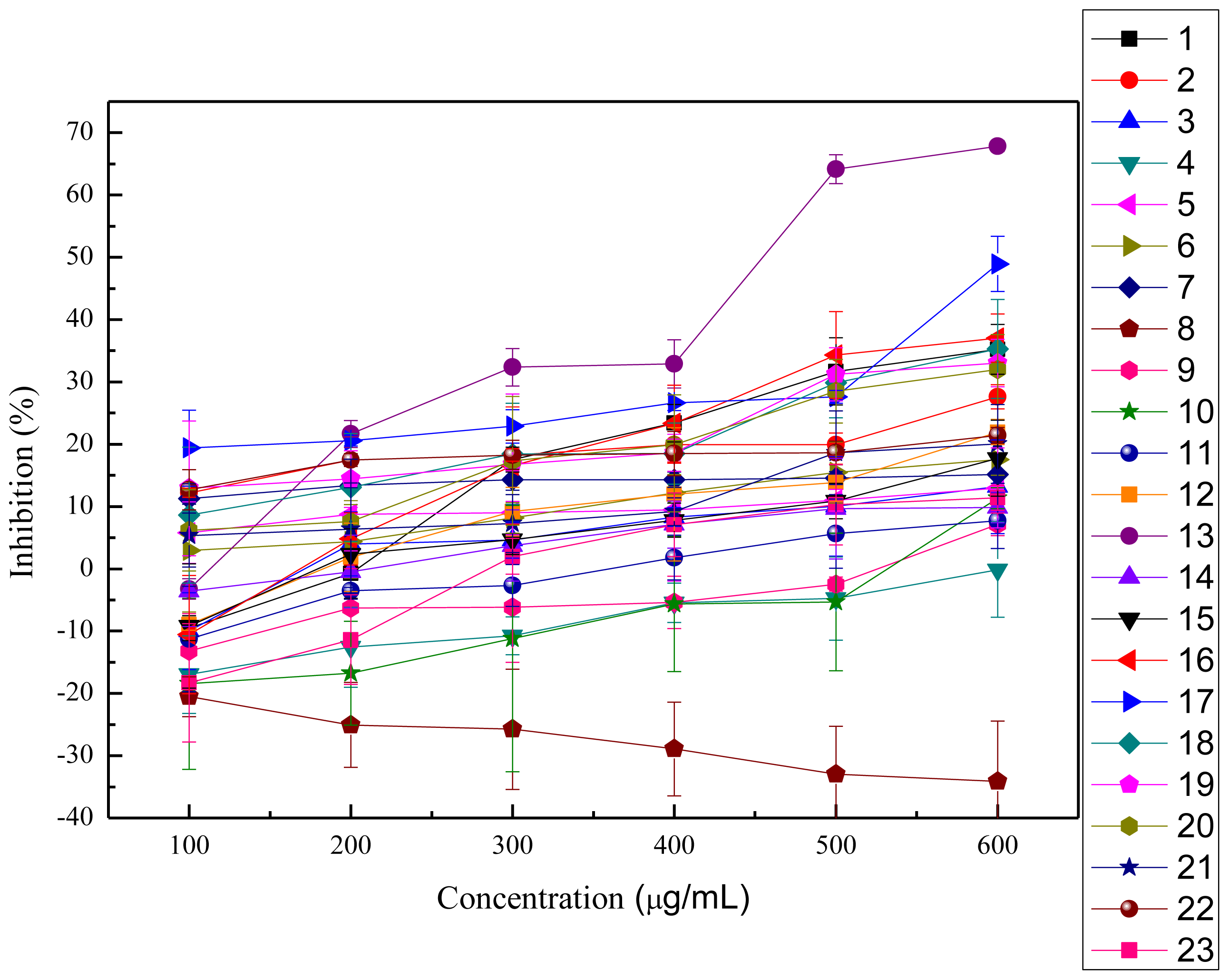
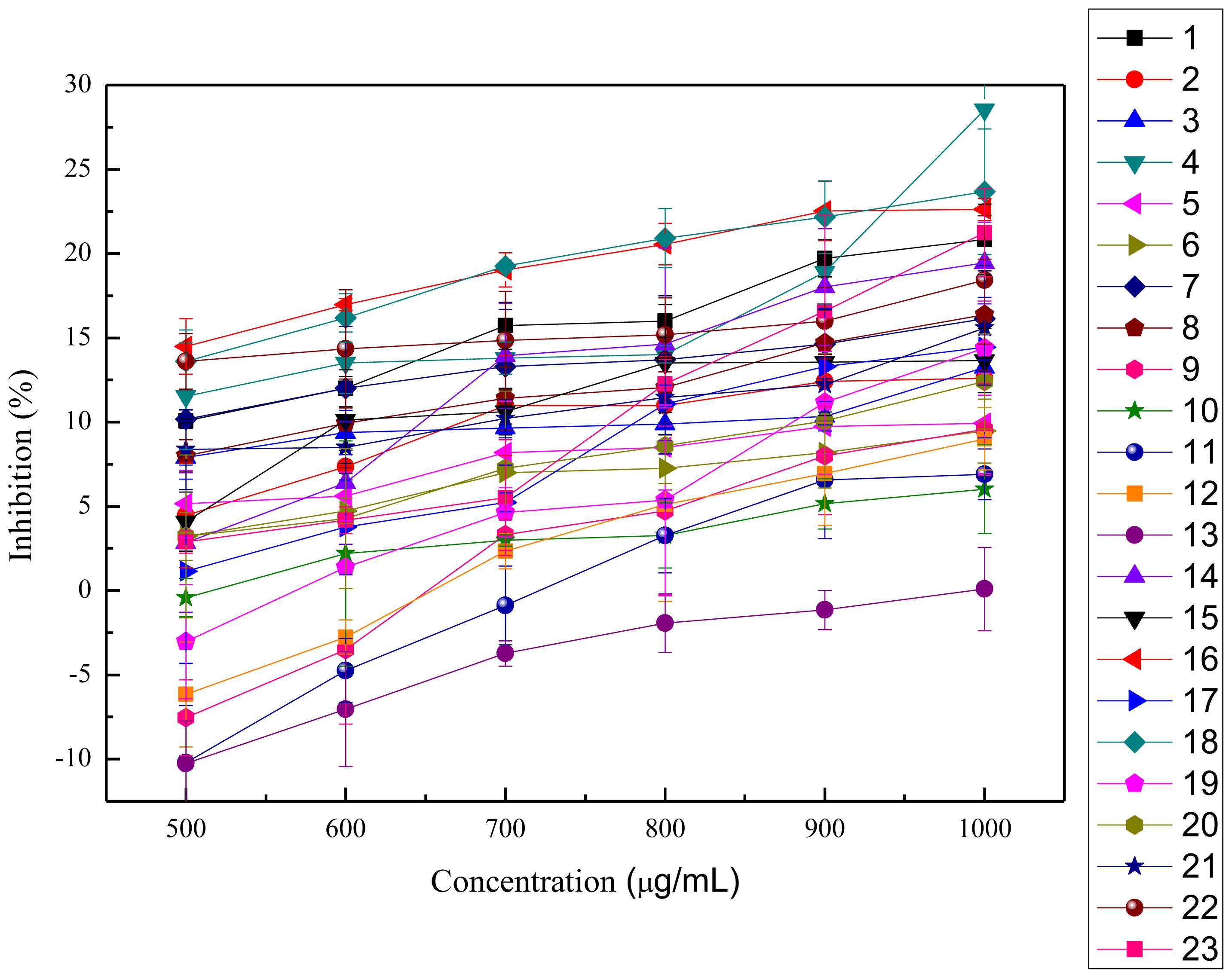
2.2.1. ABTS+ Radical-scavenging Activity
2.2.2. OH· Scavenging Assay
2.2.3. Inhibition of Lipid Peroxidation in Rat Liver Homogenate
2.3. Correlations of Polysaccharides with Antioxidant Activity
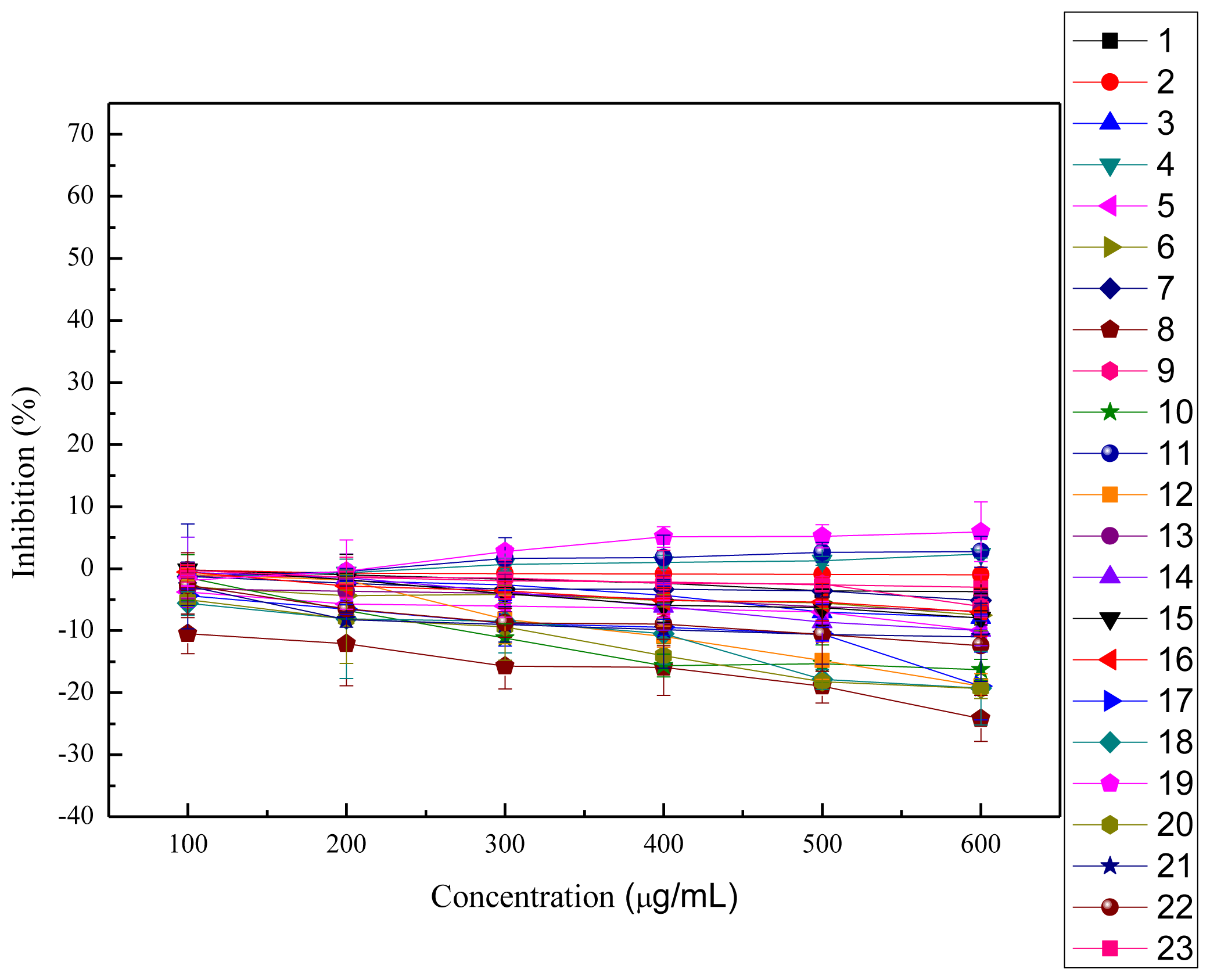
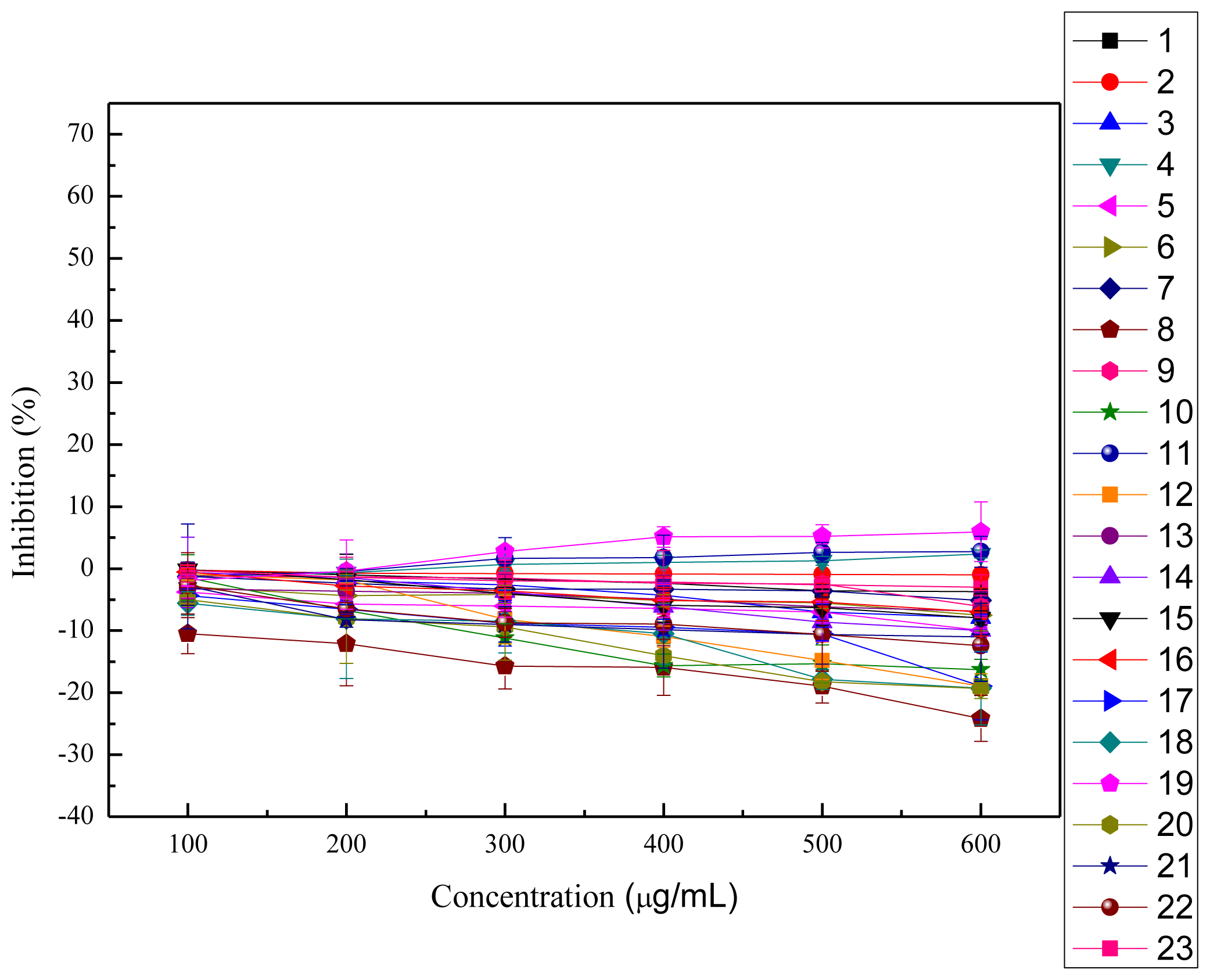
2.4. Cytotoxicity and Anti-proliferation Assay
3. Experimental Section
3.1. Chemicals
3.2. Mushrooms
3.3. Crude Polysaccharides Extraction
3.4. The Properties of Crude Polysaccharides
3.4.1. Measurement of Total Carbohydrate Contents
3.4.2. Measurement of Protein Contents
3.4.3. Determination of Total Phenolic Contents
3.5. Antioxidant Activity Assay
3.5.1. ABTS+ Radical-scavenging Activity
3.5.2. OH· Scavenging Assay
3.5.3. Inhibition of Lipid Peroxidation in Rat Liver Homogenate
3.6. Cytotoxicity and Anti-proliferation Assay
3.7. Statistical Analysis
4. Conclusions
References
- Xu, X.X.; Wu, Y.D.; Chen, H. Comparative antioxidative characteristics of polysaccharide-enriched extracts from sclerotia and cultured mycelia in submerged fermentation of Inonotus obliquus. Food Chem 2011, 127, 74–79. [Google Scholar]
- Chang, S.T.; Miles, P.G. Mushrooms: Cultivation, Nutriational Value, Medicinal Effect, and Enviromental Impact, 2nd ed; CRC Press: Boca Raton, FL, USA, 2004; pp. 1–25. [Google Scholar]
- Chang, S.T.; Buswell, J.A. Mushroom nutriceuticals. World J. Microbiol. Biotechnol 1996, 12, 473–476. [Google Scholar]
- Pan, Y.; Hao, Y.; Chu, T.W.; Li, C.Q.; Zhang, Z.F.; Zhou, Y. Ultrasonic-assisted extraction, chemical characterization of polysaccharides from Yunzhi mushroom and its effect on osteoblast cells. Carbohydr. Polym 2010, 80, 922–926. [Google Scholar]
- Ananthi, S; Raghabendran, SAHRB; Sunil, AG; Gayathri, V; Ramakrishnan, G; Vasanthi, HR. In vitro antioxidant and in vivo anti-inflammatory potential of crude polysaccharide from Turbinaria ornate. Food Chem. Toxicol. 2010, 48, 187–192. [Google Scholar]
- Liu, L.; Yuan, S.F.; Long, Y.; Guo, Z.J.; Sun, Y.; Li, Y.H.; Niu, Y.B.; Li, C.; Mei, Q.B. Immunomodulation of Rheum tanguticum polysaccharide (RTP) on the immunosuppressive effects of dexamethasone (DEX) on the treatment of colitis in rats induced by 2,4,6-trinitrobenzene sulfonic acid. Int. Immunol 2009, 9, 1568–1577. [Google Scholar]
- Godard, M.; Décordé, K.; Vetura, E.; Soteras, G.; Baccou, J.C.; Cristol, J.P.; Rouanet, J.M. Polysaccharides from the green alga Ulva rigida improve the antioxidant status and prevent fatty streak lesions in the high cholesterol fed hamster, an animal model of nutritionally-induced atherosclerosis. Food Chem 2009, 115, 176–180. [Google Scholar]
- Yoshioka, Y.; Emori, M.; Ikekawa, J.; Fukuoka, F. Isolation, purification and structure of components from acidic polysaccharides of Pleurotus ostreatus (Fr.) Quel. Carbohydr. Res 1975, 43, 305–320. [Google Scholar]
- Li, J.W.; Liu, Y.F.; Fan, L.P.; Ai, L.Z.; Shan, L. Antioxidant activities of polysaccharides from the fruiting bodies of Zizyphus Jujuba cv. Jinsixiaozao. Carbohydr. Polym 2011, 84, 390–394. [Google Scholar]
- Hu, W.C.; Han, W.; Huang, C.Q.; Wang, M.H. Protective effect of the methanolic extract from Duchesnea indica against oxidative stress in vitro and in vivo. Environ. Toxicol. Pharmacol 2011, 31, 42–50. [Google Scholar]
- Tian, L.M.; Zhao, Y.; Guo, C.; Yang, X.B. A comparative study on the antioxidant activities of an acidic polysaccharide and various solvent extracts derived from herbal Houttuynia cordata. Carbohydr. Polym 2011, 83, 537–544. [Google Scholar]
- Mattila, P.H.; Piironen, V.I.; Uusi-Rauva, E.J.; Koivistoinen, P.E. Vitamin D contents in edible mushrooms. J. Agric. Food Chem 2001, 42, 2449–2453. [Google Scholar]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Evans, C.E. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med 1999, 26, 1231–1237. [Google Scholar]
- Osman, A.M.; Wong, K.K.Y.; Fernyhough, A. ABTS radical-driven oxidation of polyphenols: Isolation and structural elucidation of covalent adducts. Biochem. Biophys. Res. Commun 2006, 346, 321–329. [Google Scholar]
- Liu, F.; Ooi, V.E.; Chang, S.T. Free radical scavenging activities of mushroom polysaccharide extracts. Life Sci 1997, 10, 763–771. [Google Scholar]
- Zhang, Y.L.; Lu, X.Y.; Fu, Z.B.; Wang, Z.B.; Zhang, J.B. Sulphated modification of a polysaccharide obtained from fresh persimmon fruit and antioxidant activities of the sulphated derivatives. Food Chem 2011, 127, 1084–1090. [Google Scholar]
- Liu, C.H.; Wang, C.H.; Xu, Z.L.; Wang, Y. Isolation, chemical characterization and antioxidant activities of two polysaccharides from the gel and the skin of Aloe barbadensis Miller irrigated with sea water. Proc. Biochem 2007, 42, 961–970. [Google Scholar]
- Lichtenstern, C.; Hofer, S.; Ramos, S.S. Lipid peroxidation in acute respiratory distress syndrome and liver failure. J. Surg. Res 2009, 168, 243–252. [Google Scholar]
- Reed, T.T. Lipid peroxidation and neurodegenerative disease. Free Radic. Biol. Med 2011, 51, 1302–1319. [Google Scholar]
- Cheung, L.M.; Cheung, P.C.K. Mushroom extracts with antioxidant activity against lipid peroxidation. Food Chem 2005, 89, 403–409. [Google Scholar]
- Floegel, A.; Kim, D.O.; Chung, S.J.; Koo, S.I.; Chun, O.K. Comparison of ABTS/DPPH assays to measure antioxidant capacity in popular antioxidant-rich US foods. J. Food. Compos. Anal 2011, 24, 1043–1048. [Google Scholar]
- Ramful, D.; Tarnus, E.; Aruoma, O.I.; Bourdon, E.; Bahorun, T. Polyphenol composition, vitamin C content and antioxidant capacity of Mauritiafn citrus fruit pulps. Food Res. Int 2011, 44, 2088–2099. [Google Scholar]
- Wang, Y.F.; Mao, F.F.; Wei, X.L. Characterization and antioxidant activities of polysaccharide from leaves, flowers and seeds of green tea. Carbohydr. Polym 2012, 88, 146–153. [Google Scholar]
- Zhao, Q.S.; Xie, B.X.; Yan, J.; Zhao, F.C.; Xiao, J.; Yao, L.Y.; Zhao, B.; Huang, Y.X. In vitro antioxidant and antitumor activities of polysaccharides extracted from Asparagus officinalis. Carbohydr. Polym 2012, 87, 392–396. [Google Scholar]
- Chen, X.; Xu, X.; Zhang, L.; Zeng, F. Chain conformation and anti-tumor activities of phosphorylated (1→3)-[β]-D-glucan from Poria cocos. Carbohydr. Polym 2009, 78, 581–587. [Google Scholar]
- Hung, C.F.; Hsu, B.Y.; Chang, S.C.; Chen, B.H. Antiproliferation of melanomacells by polysaccharide isolated from Zizyphus Jujuba. Nutrition 2012, 28, 98–105. [Google Scholar]
- Fujimoto, T.; Omote, K.; Mai, M.; Natsuume-Sakai, S. Evaluation of basic procedures for adoptive immunotherapy for gastric cancer. Biotherapy 1992, 5, 153–163. [Google Scholar]
- Ying, Z.; Han, X.X.; Li, J.R. Ultrasound-assisted extraction of polysaccharides from mulberry leaves. Food Chem 2011, 127, 1273–1279. [Google Scholar]
- Zou, Y.F.; Chen, X.F.; Yang, W.Y.; Liu, S. Response surface methodology for optimization of the ultrasonic extraction of polysaccharides from Codonopsis pilosula Nannf. Var. modesta L. T. Shen. Carbohydr. Polym 2011, 84, 503–508. [Google Scholar]
- Thestrimuang, C.; Khammuang, S.; Chiablaem, K.; Srisomsap, C.; Sarnthima, R. Antioxidant properties and cytotoxicity of crude polysaccharides from Lentinus polychrous Lév. Food Chem 2011, 128, 634–639. [Google Scholar]
- Ma, X.W.; Wu, H.X.; Liu, L.Q.; Yao, Q.S.; Wang, S.B.; Zhan, R.L.; Xing, S.S.; Zhou, Y.G. Polyphenolic compounds and antioxidant properties in mango fruits. Sci. Houtic 2011, 129, 102–107. [Google Scholar]
- Aparicio, I.M.; Peinado, C.M.; Escrig, A.J.; Rupérez, P. Multifunctional antioxidant activity of polysaccharide fractions from the soybean byproduct okara. Carbohydr. Polym 2010, 82, 245–250. [Google Scholar]
- Barahona, T.; Chandía, N.P.; Encinas, M.V.; Matsuhiro, B. Antioxidant capacity of sulfated polysaccharides from seaweeds. A kinetic approach. Food Hydrocoll 2011, 25, 529–535. [Google Scholar]
- Su, X.Y.; Wang, Z.Y.; Liu, J.R. In vitro and in vivo antioxidant activity of Pinus Koraiensis seed extract containing phenolic compounds. Food Chem 2009, 117, 681–686. [Google Scholar]
- Fan, Z.L.; Wang, Z.Y.; Liu, J.R. Cold-field extracts exert different antioxidant and antiproliferative activities in vitro. Food Chem 2011, 129, 402–407. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mushrooms | ABTS+ (μM TE/g) | OH· (mg VCE/g) | LPO (mg VCE/g) | |
|---|---|---|---|---|
| Dictyophora indusiata | Acid extract | 24.28 ± 0.54 a | 60.72 ± 0.83 a | 5.61 ± 0.11 a |
| Water extract | 16.68 ± 0.41 b | 127.38 ± 1.58 b | 19.46 ± 0.27 b | |
| Alkaline extract | 22.35 ± 0.35 c | 46.01 ± 0.81 c | 14.28 ± 0.28 c | |
| Hypsizygus marmoreus | Acid extract | 59.17 ± 0.18 d | 86.91 ± 1.02 d | 21.74 ± 0.78 d |
| Water extract | 26.93 ± 0.21 e | 109.42 ± 1.55 e | 4.25 ± 0.23 e | |
| Alkaline extract | 45.98 ± 1.77 f,l | 46.61 ± 0.94 c | 1.51 ± 0.24 f | |
| Lentinus edodes | Acid extract | 47.23 ± 0.81 f | 68.58 ± 1.44 f | 6.10 ± 0.34 a,g,h |
| Water extract | 35.38 ± 0.52 g | 84.54 ± 0.27 g | 6.17 ± 0.16 g,h | |
| Alkaline extract | 10.97 ± 0.12 h | 46.48 ± 0.51 c | 13.98 ±0.23 c | |
| Russula vinosa Lindblad | Acid extract | 65.98 ± 1.74 i | 83.93 ± 0.27 g | 6.62 ± 0.33 g |
| Water extract | 9.65 ± 0.11 j | 96.44 ± 0.39 h | 0.70 ± 0.04 i | |
| Alkaline extract | 49.88 ± 2.50 k | 50.58 ± 0.58 i | 1.57 ± 0.06 f | |
| Hohenbuehelia serotina | Acid extract | 50.01 ± 1.25 k | 76.41 ± 0.45 j | 18.58 ± 0.41 j |
| Water extract | 45.33 ± 0.44 l | 110.64 ± 0.64 e | 33.54 ± 0.49 k | |
| Alkaline extract | 41.10 ± 1.33 m | 39.82 ± 0.37 k | 9.21 ± 0.12 l | |
| Pleurotus eryngii Quel | Acid extract | - | - | - |
| Water extract | 25.41 ± 0.35 a | 100.90 ± 1.00 l | 10.57 ± 0.32 m | |
| Alkaline extract | 8.88 ± 0.12 j | 26.49 ± 0.40 m | 1.99 ± 0.08 f | |
| Hericium erinaceus | Acid extract | 54.32 ± 0.35 n | 53.50 ± 0.61 n | 19.25 ± 0.34 b |
| Water extract | 16.05 ± 0.19 b | 101.25 ± 1.48 l | 14.44 ± 0.34 c | |
| Alkaline extract | 31.66 ± 0.38 o | 41.26 ± 0.44 o | 5.97 ± 0.09 a,g | |
| Auricularia Auricula | Acid extract | 1.70 ± 0.42 p | 5.06 ± 0.12 p | 8.70 ± 0.51 l |
| Water extract | 14.16 ± 0.29 q | 59.71 ± 0.70 a | 4.00 ± 0.13 e | |
| Alkaline extract | 4.00 ± 0.03 r | 27.93 ± 0.55 q | 2.63 ± 0.15 n |
| Crude polysaccharides | ABTS+· | OH· | LPO | |
|---|---|---|---|---|
| Crude polysaccharides | 1 | 0.401 ** | 0.129 | 0.102 |
| ABTS+· | 0.401 ** | 1 | 0.327 ** | 0.346 ** |
| OH· | 0.129 | 0.327 ** | 1 | 0.464 ** |
| LPO | 0.102 | 0.346 ** | 0.464 ** | 1 |
| Mushrooms | HeLa (% inhibition) | HepG2 (% inhibition) | |
|---|---|---|---|
| Dictyophora indusiata | Acid extract | 35.23 ± 4.00 | 20.83 ± 2.10 |
| Water extract | 34.15 ± 9.70 | 16.37 ± 3.30 | |
| Alkaline extract | 37.03 ± 3.87 | 22.62 ± 0.66 | |
| Hypsizygus marmoreus | Acid extract | −0.15 ± 0.65 | 28.55 ± 4.78 |
| Water extract | 7.74 ± 4.49 | 6.90 ± 1.50 | |
| Alkaline extract | 33.03 ± 3.80 | 14.39 ± 2.79 | |
| Lentinus edodes | Acid extract | 13.18 ± 7.51 | 13.24 ± 4.16 |
| Water extract | 11.33 ± 1.70 | 6.02 ± 2.64 | |
| Alkaline extract | 35.28 ± 7.95 | 23.68 ± 3.72 | |
| Russula vinosa Lindblad | Acid extract | 27.59 ± 1.95 | 12.60 ± 2.96 |
| Water extract | 7.13 ± 1.80 | 9.56 ± 2.73 | |
| Alkaline extract | 48.92 ± 4.43 | 14.44 ± 0.75 | |
| Hohenbuehelia serotina | Acid extract | 12.97 ± 6.42 | 9.93 ± 2.97 |
| Water extract | 21.95 ± 2.01 | 9.01 ± 1.86 | |
| Alkaline extract | 32.00 ± 5.62 | 12.41 ± 2.81 | |
| Pleurotus eryngii Quel | Acid extract | - | - |
| Water extract | 11.74 ± 6.10 | 13.66 ± 1.92 | |
| Alkaline extract | 11.38 ± 2.01 | 21.24 ± 2.62 | |
| Hericium erinaceus | Acid extract | 17.54 ± 2.52 | 9.47 ± 2.97 |
| Water extract | 67.85 ± 0.87 | 0.09 ± 0.47 | |
| Alkaline extract | 20.05 ± 6.30 | 15.59 ± 3.39 | |
| Auricularia Auricula | Acid extract | 15.13 ± 0.45 | 16.14 ± 3.16 |
| Water extract | 9.85 ± 2.46 | 19.45 ± 2.42 | |
| Alkaline extract | 21.44 ± 1.29 | 18.44 ± 3.81 |
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Li, X.; Wang, Z.; Wang, L.; Walid, E.; Zhang, H. In Vitro Antioxidant and Anti-Proliferation Activities of Polysaccharides from Various Extracts of Different Mushrooms. Int. J. Mol. Sci. 2012, 13, 5801-5817. https://doi.org/10.3390/ijms13055801
Li X, Wang Z, Wang L, Walid E, Zhang H. In Vitro Antioxidant and Anti-Proliferation Activities of Polysaccharides from Various Extracts of Different Mushrooms. International Journal of Molecular Sciences. 2012; 13(5):5801-5817. https://doi.org/10.3390/ijms13055801
Chicago/Turabian StyleLi, Xiaoyu, Zhenyu Wang, Lu Wang, Elfalleh Walid, and Hua Zhang. 2012. "In Vitro Antioxidant and Anti-Proliferation Activities of Polysaccharides from Various Extracts of Different Mushrooms" International Journal of Molecular Sciences 13, no. 5: 5801-5817. https://doi.org/10.3390/ijms13055801