Structural and Functional Characterization of Two Alternative Splicing Variants of Mouse Endothelial Cell-Specific Chemotaxis Regulator (ECSCR)

Abstract
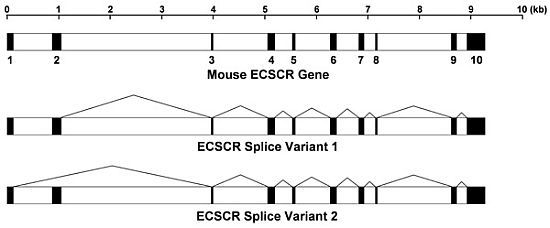
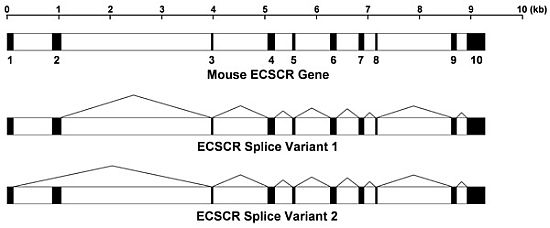
:
{kind=link}
{kind=link}
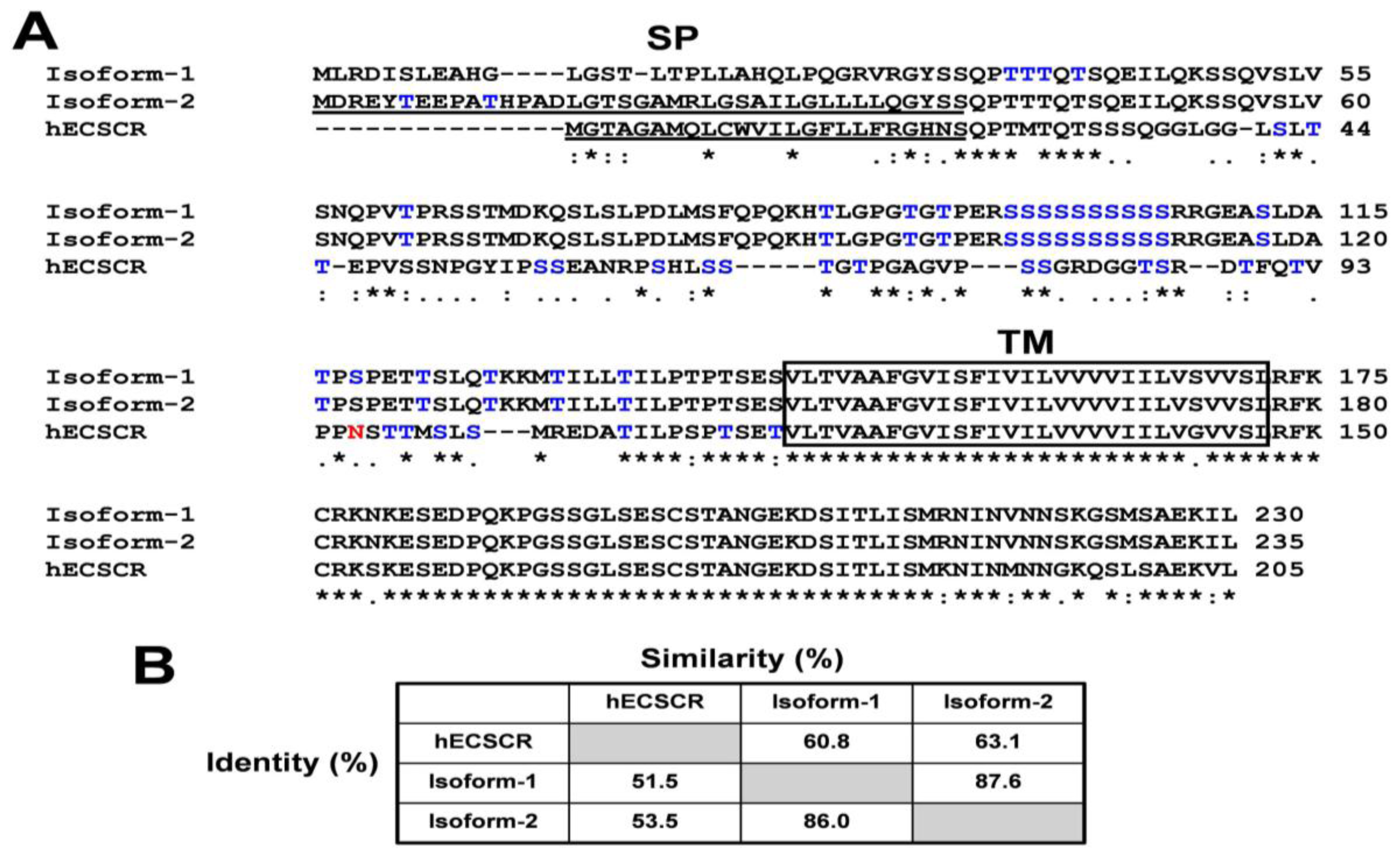{kind=link}
{kind=link}
{kind=link}
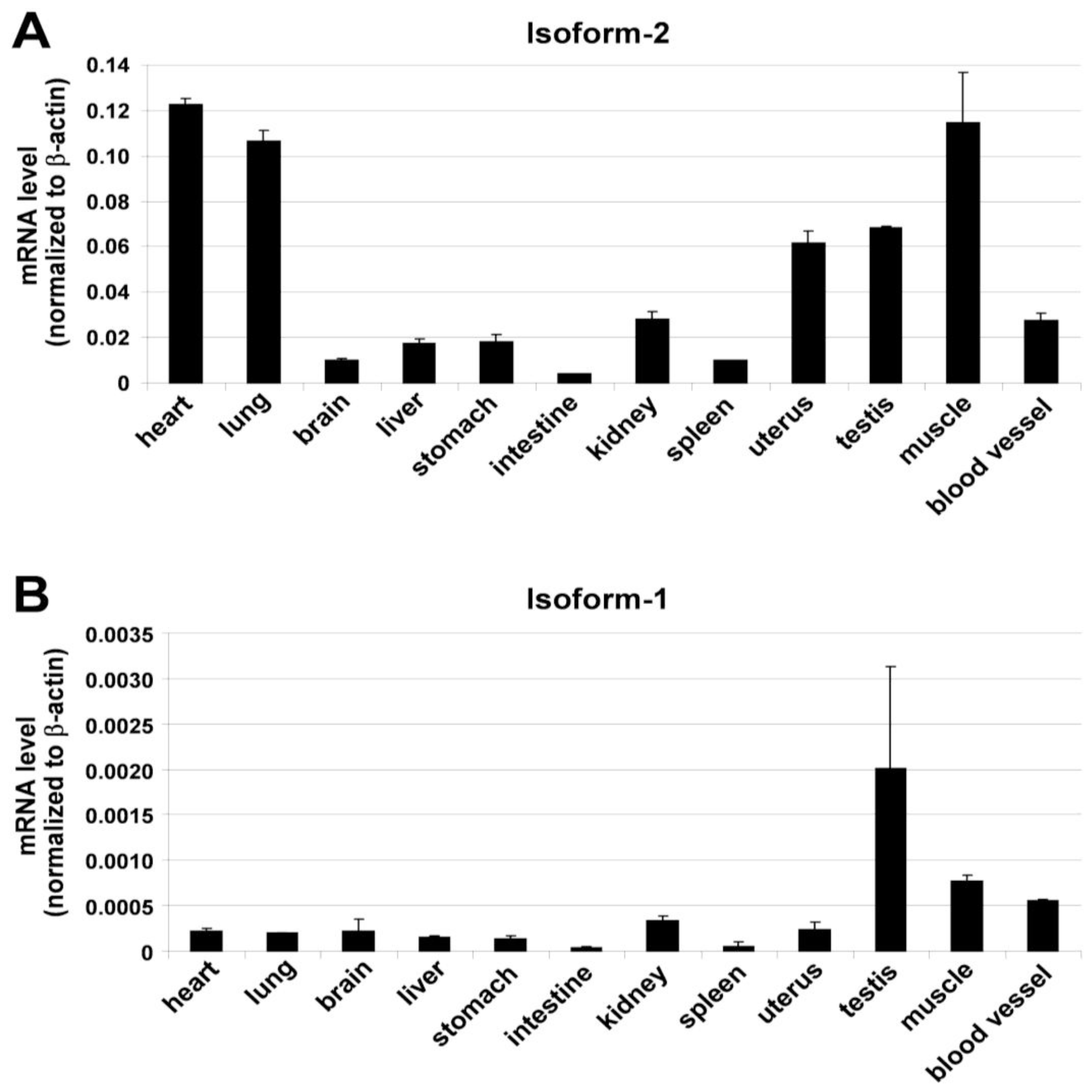{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Experimental Section
2.1. Reagents and Antibodies
2.2. RNA Extraction, RT-PCR, and Quantitative Real-Time PCR
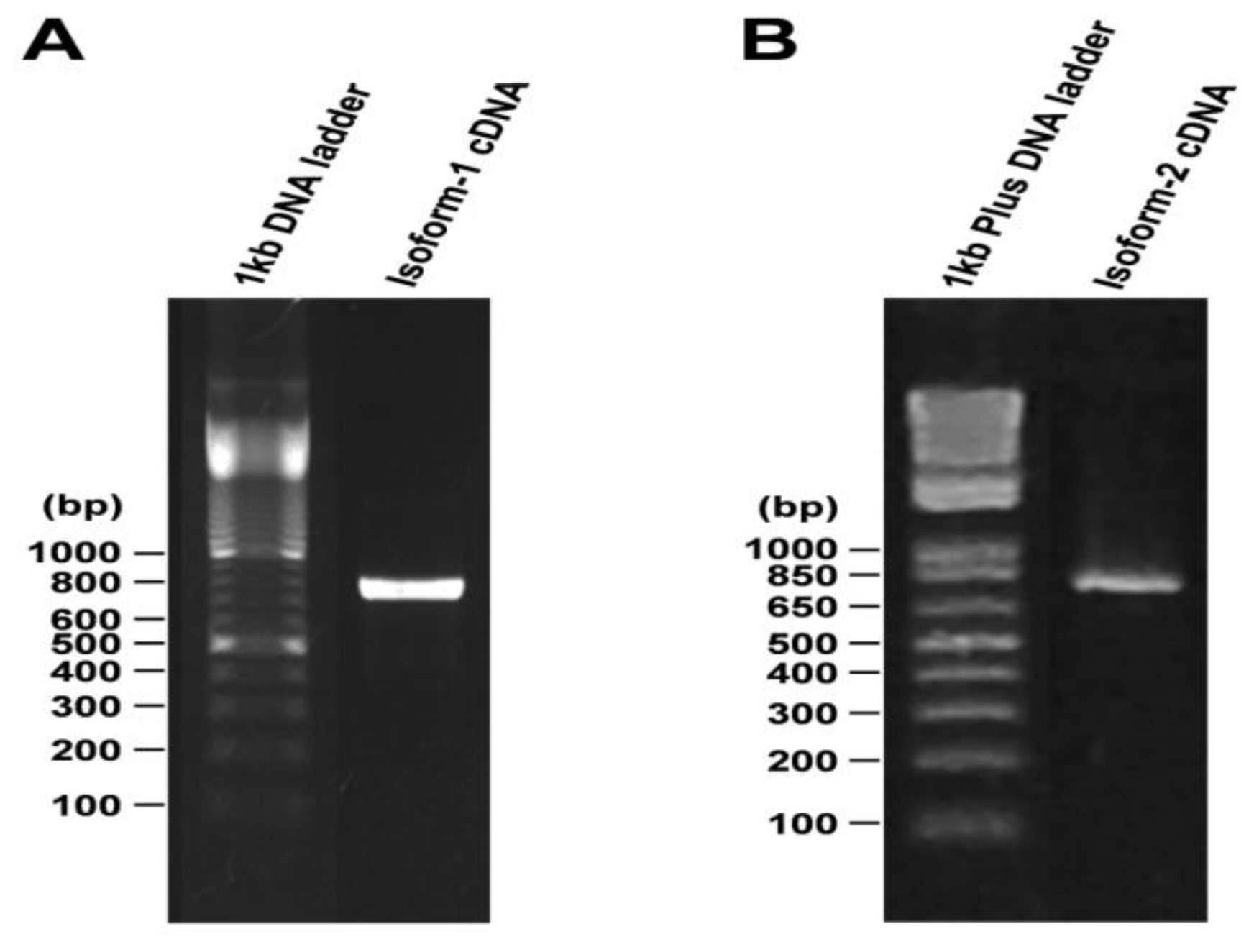
2.3. Plasmid Constructs and Sequencing
2.4. Cell Culture and Transfection
2.5. Protein Extraction and Western Blot Analysis (Immunoblotting)
2.6. Immunofluorescent Staining and Microscopy
2.7. Cell Migration Assay
2.8. Statistical Analysis
2.9. Protein-Protein Basic Local Alignment Search Tool (BLASTP)
2.10. Multiple Sequence Alignment
2.11. Sequence Translation
2.12. Prediction of Signal Peptide and Transmembrane Domain
2.13. Prediction of Glycosylation Sites
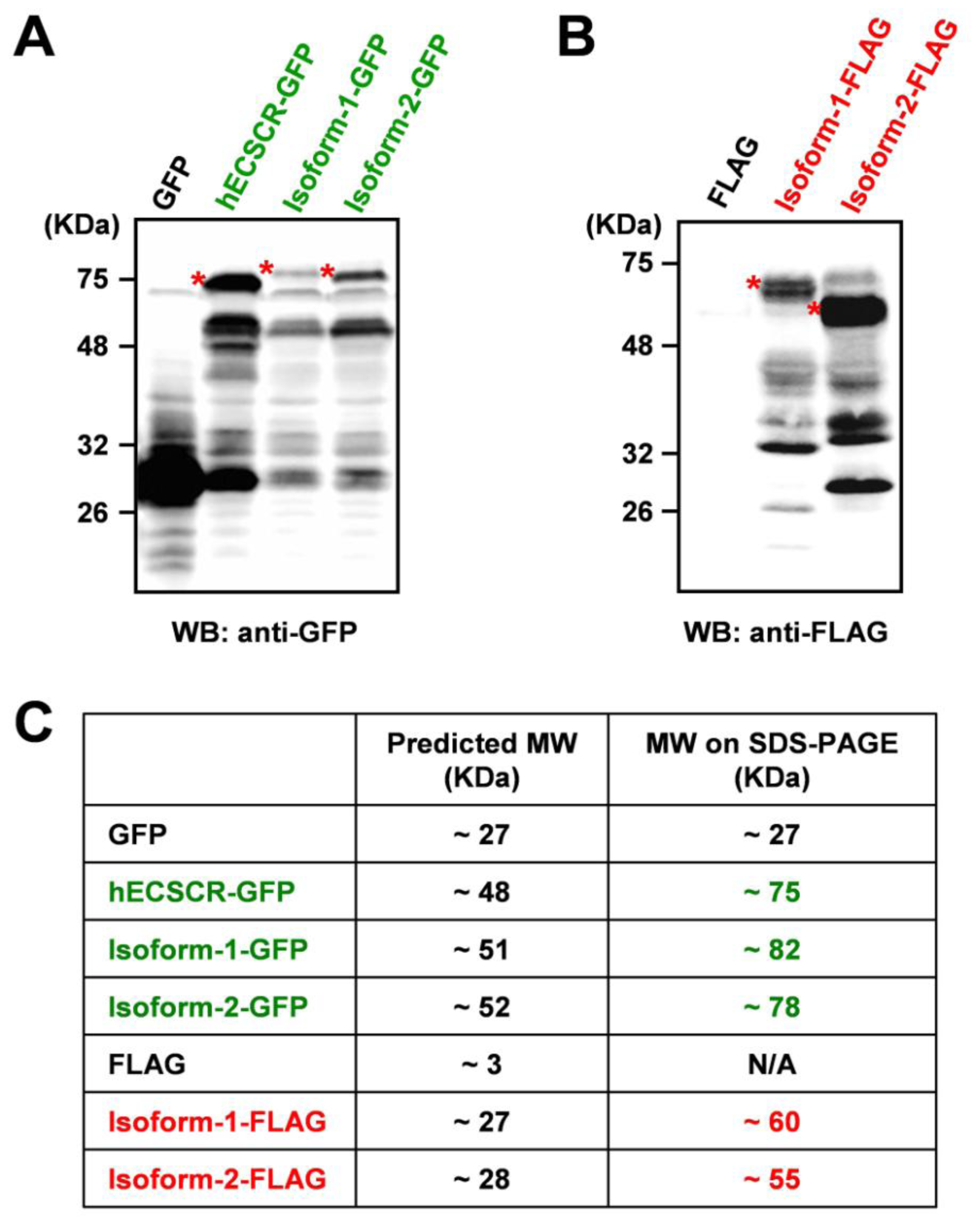
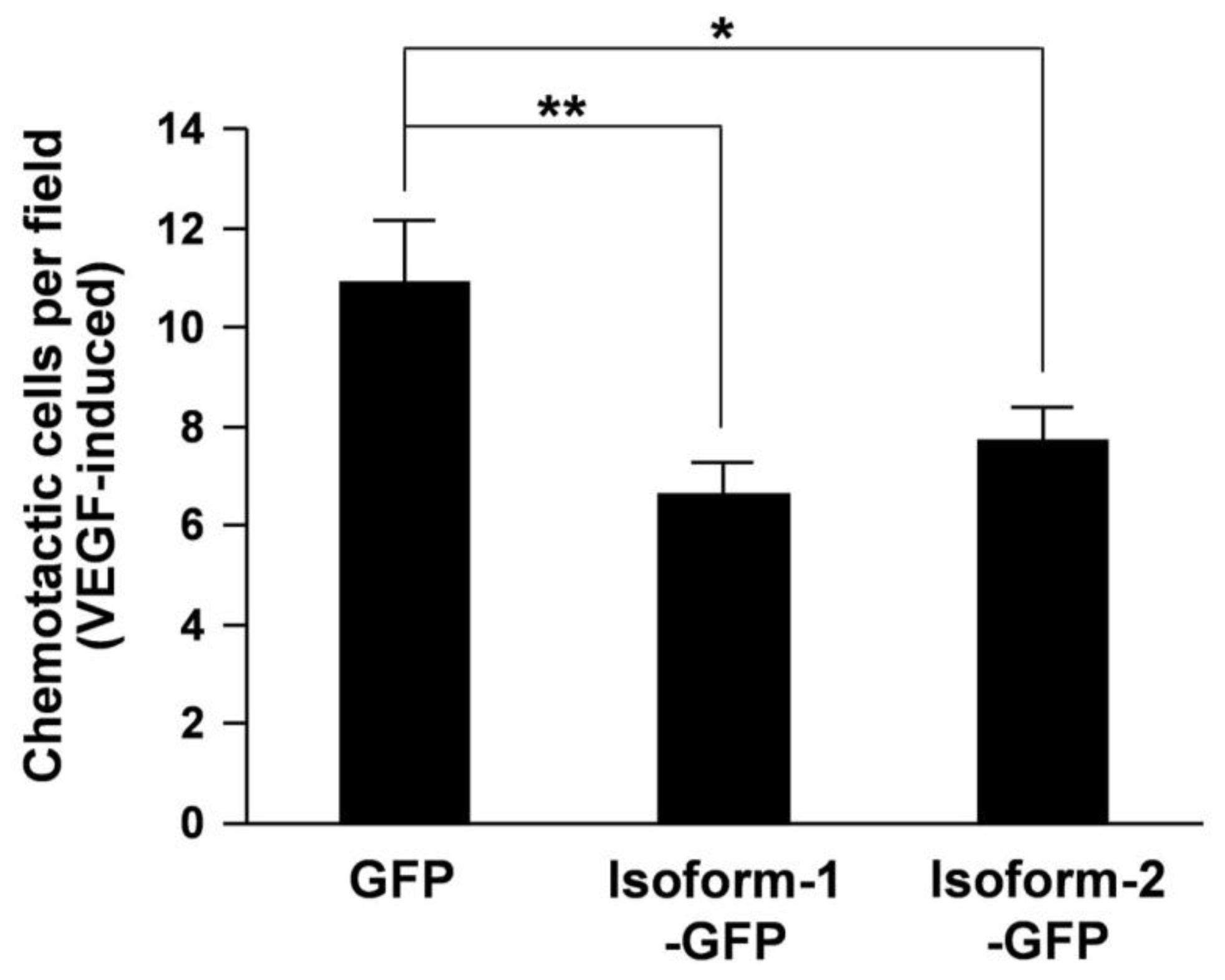
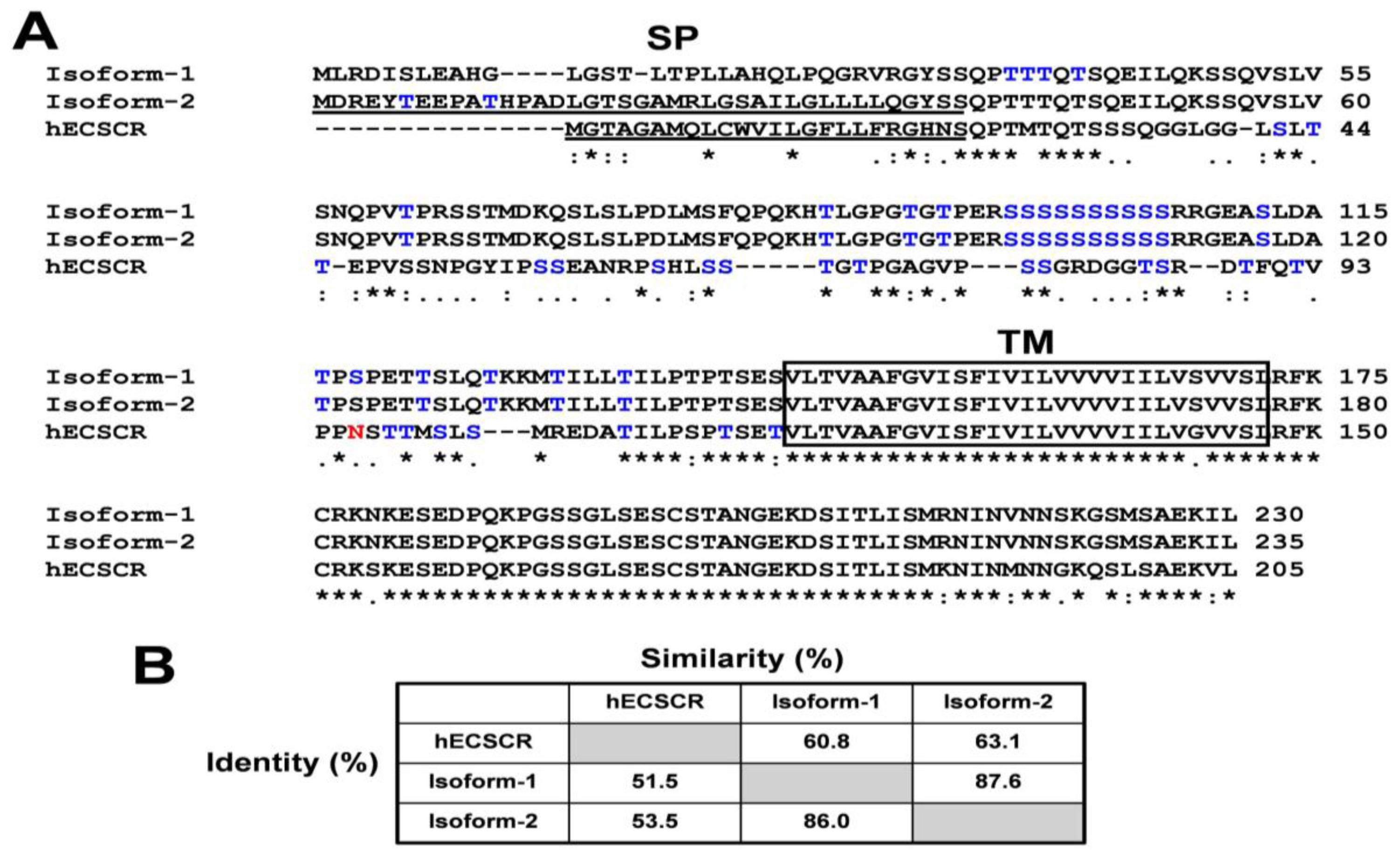
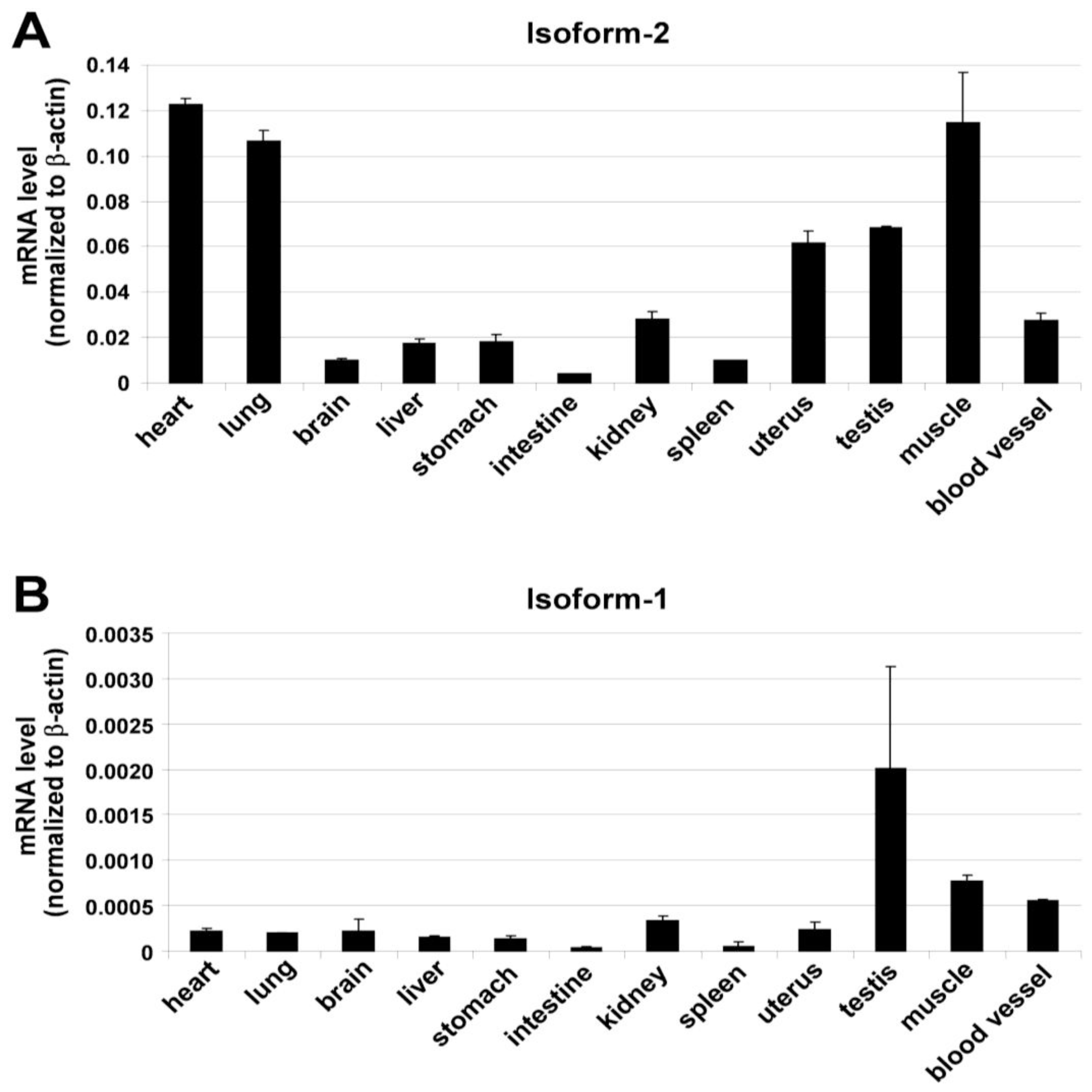
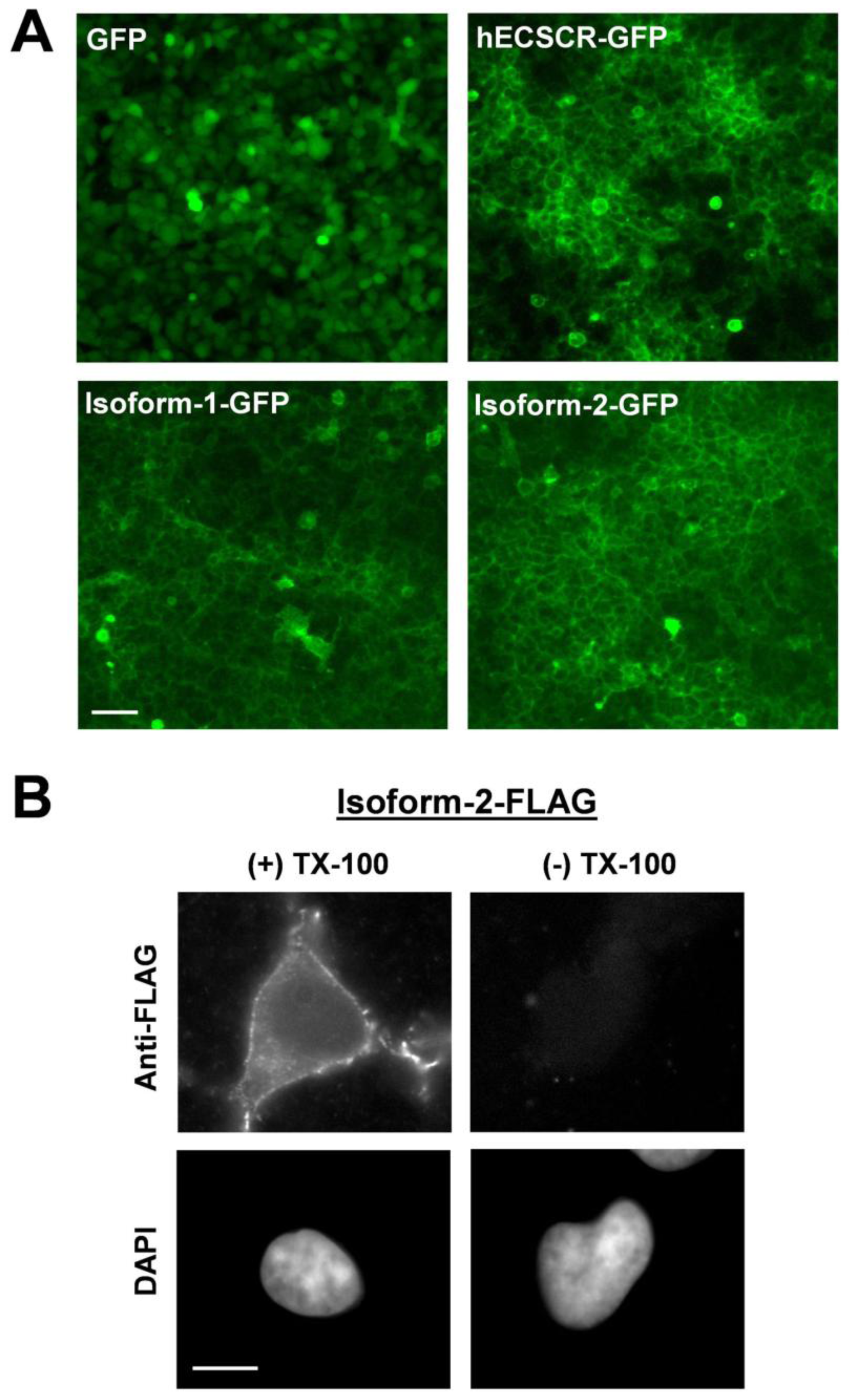
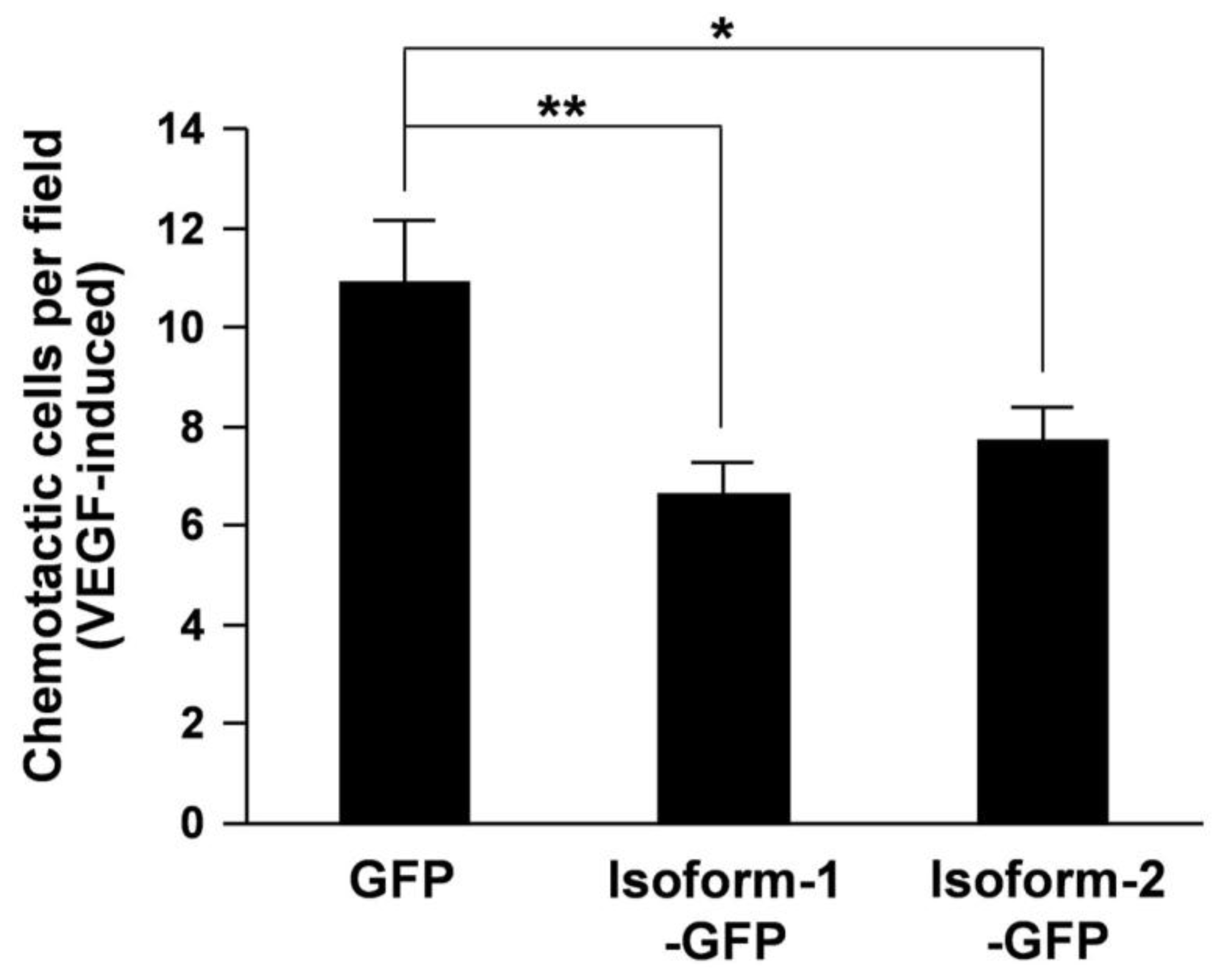
3. Results and Discussion
4. Conclusions
Supplementary Materials
ijms-13-04920-s001.pdfAcknowledgements
References
- Risau, W. Mechanisms of angiogenesis. Nature 1997, 386, 671–674. [Google Scholar]
- Carmeliet, P.; Jain, R.K. Angiogenesis in cancer and other diseases. Nature 2000, 407, 249–257. [Google Scholar]
- Folkman, J.; D’Amore, P.A. Blood vessel formation: What is its molecular basis? Cell 1996, 87, 1153–1155. [Google Scholar]
- Jacobs, J. Combating cardiovascular disease with angiogenic therapy. Drug Discov. Today 2007, 12, 1040–1045. [Google Scholar]
- Rahimi, N. Vascular endothelial growth factor receptors: Molecular mechanisms of activation and therapeutic potentials. Exp. Eye Res 2006, 83, 1005–1016. [Google Scholar]
- Lamalice, L.; Le Boeuf, F.; Huot, J. Endothelial cell migration during angiogenesis. Circ. Res 2007, 100, 782–794. [Google Scholar]
- Newman, P.J.; Berndt, M.C.; Gorski, J.; White, G.C.; Lyman, S.; Paddock, C.; Muller, W.A. PECAM-1 (CD31) cloning and relation to adhesion molecules of the immunoglobulin gene superfamily. Science 1990, 247, 1219–1222. [Google Scholar]
- Osborn, L.; Hession, C.; Tizard, R.; Vassallo, C.; Luhowskyj, S.; Chi-Rosso, G.; Lobb, R. Direct expression cloning of vascular cell adhesion molecule 1, a cytokine-induced endothelial protein that binds to lymphocytes. Cell 1989, 59, 1203–1211. [Google Scholar]
- Hirata, K.; Ishida, T.; Penta, K.; Rezaee, M.; Yang, E.; Wohlgemuth, J.; Quertermous, T. Cloning of an immunoglobulin family adhesion molecule selectively expressed by endothelial cells. J. Biol. Chem 2001, 276, 16223–16231. [Google Scholar]
- Suzuki, S.; Sano, K.; Tanihara, H. Diversity of the cadherin family: Evidence for eight new cadherins in nervous tissue. Cell Regul 1991, 2, 261–270. [Google Scholar]
- Ferrara, N.; Houck, K.A.; Jakeman, L.B.; Winer, J.; Leung, D.W. The vascular endothelial growth factor family of polypeptides. J. Cell. Biochem 1991, 47, 211–218. [Google Scholar]
- Huminiecki, L.; Bicknell, R. In silico cloning of novel endothelial-specific genes. Genome Res 2000, 10, 1796–1806. [Google Scholar]
- Ho, M.; Yang, E.; Matcuk, G.; Deng, D.; Sampas, N.; Tsalenko, A.; Tabibiazar, R.; Zhang, Y.; Chen, M.; Talbi, S.; et al. Identification of endothelial cell genes by combined database mining and microarray analysis. Physiol. Genomics 2003, 13, 249–262. [Google Scholar]
- Ma, F.; Zhang, D.; Yang, H.; Sun, H.; Wu, W.; Gan, Y.; Balducci, J.; Wei, Y.; Zhao, X.; Huang, Y. Endothelial cell-specific molecule 2 (ECSM2) modulates actin remodeling and epidermal growth factor receptor signaling. Genes Cells 2009, 14, 281–293. [Google Scholar]
- Armstrong, L.J.; Heath, V.L.; Sanderson, S.; Kaur, S.; Beesley, J.F.; Herbert, J.M.; Legg, J.A.; Poulsom, R.; Bicknell, R. ECSM2, an endothelial specific filamin a binding protein that mediates chemotaxis. Arterioscler. Thromb. Vasc. Biol 2008, 28, 1640–1646. [Google Scholar]
- Ikeda, K.; Nakano, R.; Uraoka, M.; Nakagawa, Y.; Koide, M.; Katsume, A.; Minamino, K.; Yamada, E.; Yamada, H.; Quertermous, T.; et al. Identification of ARIA regulating endothelial apoptosis and angiogenesis by modulating proteasomal degradation of cIAP-1 and cIAP-2. Proc. Natl. Acad. Sci. USA 2009, 106, 8227–8232. [Google Scholar]
- Koide, M.; Ikeda, K.; Akakabe, Y.; Kitamura, Y.; Ueyama, T.; Matoba, S.; Yamada, H.; Okigaki, M.; Matsubara, H. Apoptosis regulator through modulating IAP expression (ARIA) controls the PI3K/Akt pathway in endothelial and endothelial progenitor cells. Proc. Natl. Acad. Sci. USA 2011, 108, 9472–9477. [Google Scholar]
- Verma, A.; Bhattacharya, R.; Remadevi, I.; Li, K.; Pramanik, K.; Samant, G.V.; Horswill, M.; Chun, C.Z.; Zhao, B.; Wang, E.; et al. Endothelial cell-specific chemotaxis receptor (ECSCR) promotes angioblast migration during vasculogenesis and enhances VEGF receptor sensitivity. Blood 2010, 115, 4614–4622. [Google Scholar]
- Verissimo, A.R.; Herbert, J.M.; Heath, V.L.; Legg, J.A.; Sheldon, H.; Andre, M.; Swain, R.K.; Bicknell, R. Functionally defining the endothelial transcriptome, from Robo4 to ECSCR. Biochem. Soc. Trans 2009, 37, 1214–1217. [Google Scholar]
- Shi, C.; Lu, J.; Wu, W.; Ma, F.; Georges, J.; Huang, H.; Balducci, J.; Chang, Y.; Huang, Y. Endothelial cell-specific molecule 2 (ECSM2) localizes to cell-cell junctions and modulates bFGF-directed cell migration via the ERK-FAK pathway. PLoS One 2011, 6. [Google Scholar] [CrossRef]
- Black, D.L. Mechanisms of alternative pre-messenger RNA splicing. Annu. Rev. Biochem 2003, 72, 291–336. [Google Scholar]
- Hallegger, M.; Llorian, M.; Smith, C.W. Alternative splicing: Global insights. FEBS J 2010, 277, 856–866. [Google Scholar]
- Kalsotra, A.; Cooper, T.A. Functional consequences of developmentally regulated alternative splicing. Nat. Rev. Genet 2011, 12, 715–729. [Google Scholar]
- Graveley, B.R. Alternative splicing: Increasing diversity in the proteomic world. Trends Genet 2001, 17, 100–107. [Google Scholar]
- Tu, Y.; Huang, Y.; Zhang, Y.; Hua, Y.; Wu, C. A new focal adhesion protein that interacts with integrin-linked kinase and regulates cell adhesion and spreading. J. Cell Biol 2001, 153, 585–598. [Google Scholar]
- Huang, Y.; Li, J.; Zhang, Y.; Wu, C. The roles of integrin-linked kinase in the regulation of myogenic differentiation. J. Cell Biol 2000, 150, 861–872. [Google Scholar]
- Huang, Y.; Li, X.; Jiang, J.; Frank, S.J. Prolactin modulates phosphorylation, signaling and trafficking of epidermal growth factor receptor in human T47D breast cancer cells. Oncogene 2006, 25, 7565–7576. [Google Scholar]
- Gan, Y.; Shi, C.; Inge, L.; Hibner, M.; Balducci, J.; Huang, Y. Differential roles of ERK and Akt pathways in regulation of EGFR-mediated signaling and motility in prostate cancer cells. Oncogene 2010, 29, 4947–4958. [Google Scholar]
- Huang, Y.; Kim, S.O.; Jiang, J.; Frank, S.J. Growth hormone-induced phosphorylation of epidermal growth factor (EGF) receptor in 3T3-F442A cells. Modulation of EGF-induced trafficking and signaling. J. Biol. Chem 2003, 278, 18902–18913. [Google Scholar]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res 1997, 25, 3389–3402. [Google Scholar]
- Altschul, S.F.; Wootton, J.C.; Gertz, E.M.; Agarwala, R.; Morgulis, A.; Schäffer, A.A.; Yu, Y.K. Protein database searches using compositionally adjusted substitution matrices. FEBS J 2005, 272, 5101–5109. [Google Scholar]
- EMBL-EBI ClustalW2—Multiple Sequence Alignment. Available online: http://www.ebi.ac.uk/Tools/msa/clustalw2 accessed on 18 March 2012.
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar]
- ExPASy Translate Tool. Available online: http://web.expasy.org/translate accessed on 18 March 2012.
- SIG-Pred: Signal Peptide Prediction. Available online: http://bmbpcu36.leeds.ac.uk/prot_analysis/Signal.html accessed on 18 March 2012.
- PrediSi: Prediction of Signal Peptides. Available online: http://www.predisi.de/index.html accessed on 18 March 2012.
- SignalP 4.0 Server. Available online: http://www.cbs.dtu.dk/services/SignalP accessed on 18 March 2012.
- DAS—Transmembrane Prediction Server. Available online: http://www.sbc.su.se/~miklos/DAS accessed on 18 March 2012.
- Cserzö, M.; Wallin, E.; Simon, I.; von Heijne, G.; Elofsson, A. Prediction of transmembrane alpha-helices in prokaryotic membrane proteins: The dense alignment surface method. Protein Eng 1997, 10, 673–676. [Google Scholar]
- NetNGlyc 1.0 Server. Available online: http://www.cbs.dtu.dk/services/NetNGlyc accessed on 18 March 2012.
- NetOGlyc 3.1 Server. Available online: http://www.cbs.dtu.dk/services/NetOGlyc accessed on 18 March 2012.
- Julenius, K.; Mølgaard, A.; Gupta, R.; Brunak, S. Prediction, conservation analysis, and structural characterization of mammalian mucin-type O-glycosylation sites. Glycobiology 2005, 15, 153–164. [Google Scholar]
- Mouse ECSCR sequence with NCBI Reference Sequence: NP_001028313.1. Available online: http://www.ncbi.nlm.nih.gov/protein/NP_001028313.1 accessed on 18 March 2012.
- Skarnes, W.C.; Rosen, B.; West, A.P.; Koutsourakis, M.; Bushell, W.; Iyer, V.; Mujica, A.O.; Thomas, M.; Harrow, J.; Cox, T.; et al. A conditional knockout resource for the genome-wide study of mouse gene function. Nature 2011, 474, 337–342. [Google Scholar]
- Hansen, G.M.; Markesich, D.C.; Burnett, M.B.; Zhu, Q.; Dionne, K.M.; Richter, L.J.; Finnell, R.H.; Sands, A.T.; Zambrowicz, B.P.; Abuin, A. Large-scale gene trapping in C57BL/6N mouse embryonic stem cells. Genome Res 2008, 18, 1670–1679. [Google Scholar]
- ExPASy Proteomic Tools. Available online: http://www.expasy.org/proteomics accessed on 18 March 2012.
- Mus musculus 181000113601716 genomic scaffold, whole genome shotgun sequence. Available online: http://www.ncbi.nlm.nih.gov/nuccore/70979735?from=19601084&to=19610340&report=gbwithparts accessed on 18 March 2012.



© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Wu, W.; Shi, C.; Ma, F.; Balducci, J.; Huang, H.; Ji, H.-L.; Chang, Y.; Huang, Y. Structural and Functional Characterization of Two Alternative Splicing Variants of Mouse Endothelial Cell-Specific Chemotaxis Regulator (ECSCR). Int. J. Mol. Sci. 2012, 13, 4920-4936. https://doi.org/10.3390/ijms13044920
Wu W, Shi C, Ma F, Balducci J, Huang H, Ji H-L, Chang Y, Huang Y. Structural and Functional Characterization of Two Alternative Splicing Variants of Mouse Endothelial Cell-Specific Chemotaxis Regulator (ECSCR). International Journal of Molecular Sciences. 2012; 13(4):4920-4936. https://doi.org/10.3390/ijms13044920
Chicago/Turabian StyleWu, Wen, Chunwei Shi, Fanxin Ma, James Balducci, Hanju Huang, Hong-Long Ji, Yongchang Chang, and Yao Huang. 2012. "Structural and Functional Characterization of Two Alternative Splicing Variants of Mouse Endothelial Cell-Specific Chemotaxis Regulator (ECSCR)" International Journal of Molecular Sciences 13, no. 4: 4920-4936. https://doi.org/10.3390/ijms13044920