1. Introduction
Arabinoxylan and β-glucan naturally occur in the endosperm and aleurone cell walls of cereal grains with varying degrees of solubility. The content of rye and wheat arabinoxylan varies in the range of 6%–12% in the whole grain, and 21%–25% in the bran [
1–
3]. The proportion of β-glucan in whole grain oat is approximately 3% and 8% in the oat bran [
4], whereas in the content of whole grain rye is reported to be ~1.0%–2.5% with double the amount in bran fractions [
5]. Arabinoxylan is a random coil with varying degree of flexibility consisting of linked
d-xylopyranosyl units with α-
l-arabinofuranoside residues substituted at the 2- and 3-carbon position [
6]. (1–3)(1–4)-β-
d-glucan known as β-glucan is a linear glucosidic polysaccharides consisting of 60% 3-
O-β-
d-cellobiosyl-
d-glucose, 30% 3-
O-β-
d-cellotriosyl-
d-glucose, and 10% higher 3-
O-β-
d-cellooligosyl-
d-glucose [
7].
Due to the capability of soluble arabinoxylan and β-glucan to increase viscosity, they have physiologically gained a lot of interest as polysaccharides that can attenuate blood glucose and insulin responses and lower blood cholesterol [
2,
8,
9]. Furthermore, the viscosity-elevating properties of arabinoxylan and β-glucan have been shown to slow the rate of gastric emptying and reduce the motility of human small intestine, which may reflect a prolongation of satiety [
10]. Raised gastrointestinal viscosity can also reduce the rate of digestion and absorption of macronutrients, including fat, protein and carbohydrates [
8,
10] which may have a different effect on weight management [
11,
12].
Several physical, chemical and enzymatic methods have been reported in the literature to extract soluble arabinoxylan and β-glucan from foodstuffs such as grains, flour, rice, breads or pasta. The most common procedures are enzymatic, alkaline or water extraction with varying temperature and incubation time [
4]. Boiling the sample for 15 min in 50% ethanol stabilizes the extract and inactivates endogenous or microbial enzymes [
4]. Moreover, it allows the removal of lipid and enhances the efficiency of DF extraction. Water extraction at low temperature, however, brings a risk of insufficient extraction and solubilization of DF from the complex DF matrix, and increases the risk of aggregation in the water solution [
8]. On the contrary, alkaline extraction enhances the DF solubilization and prevents aggregation [
4] resulting in higher recovery than water- and enzymatic extraction procedures. Enzymatic and alkali treatments are used to break the ester and other covalent and/or non-covalent linkages between components and release the initially unextractable polysaccharides from the complex network of the cell walls grains [
13]. Despite differences in the type of method of DF extraction, there are some general structural features that affect the extractability and solubility of DF: branching, glycosidic bonds, ionizing groups, and non-uniformity in repeating structure [
13].
The molecular weight of DF components, their structure, solubility and concentration strongly influence the viscosity of aqueous solutions [
14]. The molecular weight varies from 0.04 to 9 × 10
6 g/mol of arabinoxylan in wheat and rye, with a polydispersity index (PDI) of 1.7–2 in wheat, indicating a broad distribution [
1,
3]. For comparison, the molecular weight distribution of β-glucan ranges from 10
4 to 5 × 10
6 g/mol in rye [
1] and 0.04 to 2.5 × 10
6 g/mol in oat [
15], with a polydispersity index among cereals of 1.2–3.1 [
15]. The polydispersity index reflects heterogenity of sizes of molecules in a polymer population [
3]. However, mixtures of different polymers with uneven distribution will also result in a higher PDI.
During mixing, yeast fermentation and baking the DF polysaccharides provided as ingredients in the flour may change regarding molecular weight distribution and polydispersity [
1]. Changes in the molecular weight and concentration of soluble DF influence the extract viscosity, which may have an impact on the functional properties of the breads and thereby on the physiological responses when consumed by monogastric mammals. As the gastrointestinal tract and physiological responses of pig are very similar to humans, the pig was used as a monogastric model providing access to digestive material that is difficult to obtain from humans. The aim of this investigation was to study the fate of polysaccharides in breads with variable content and structure of fiber constituents before and after
in vitro and
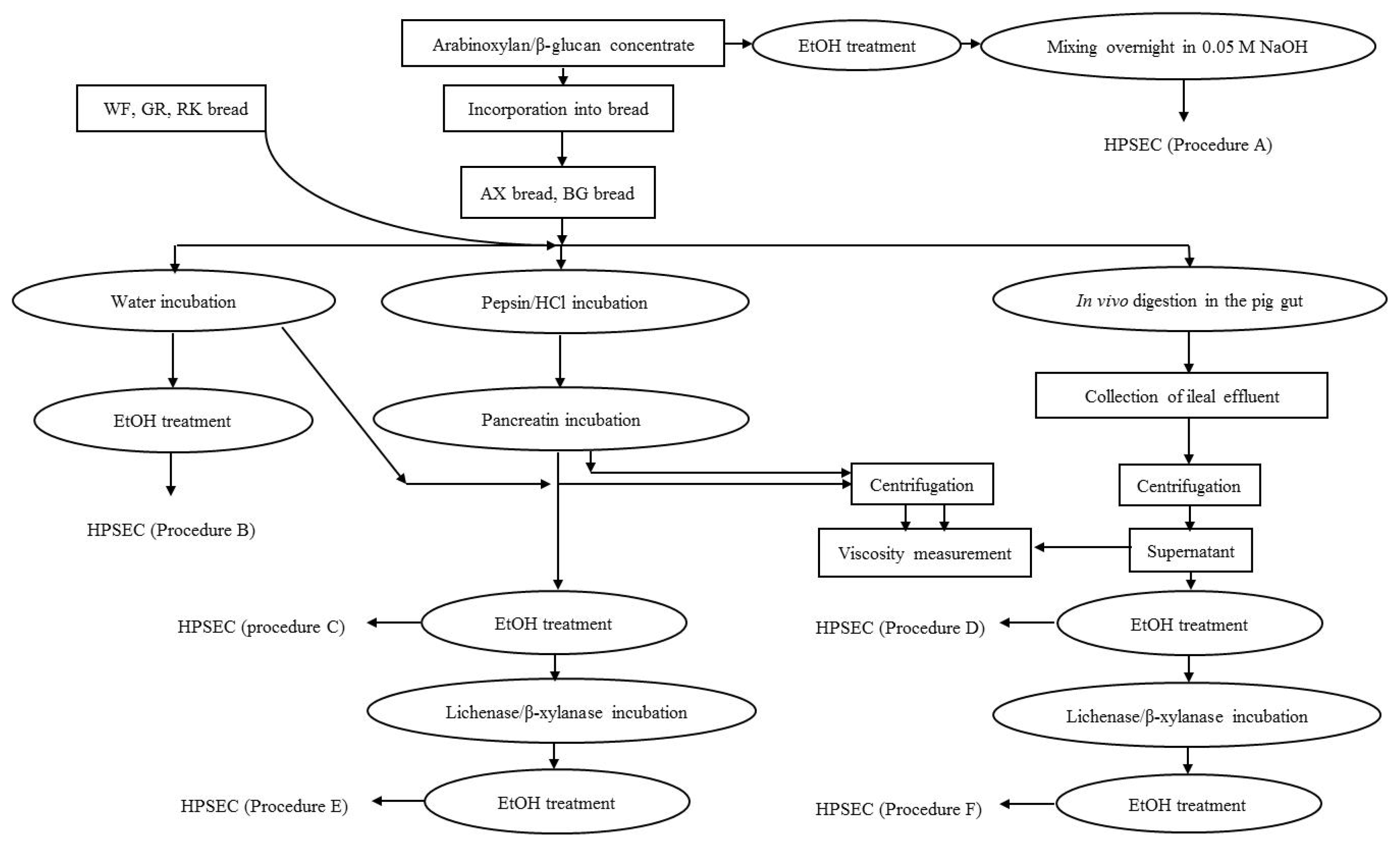
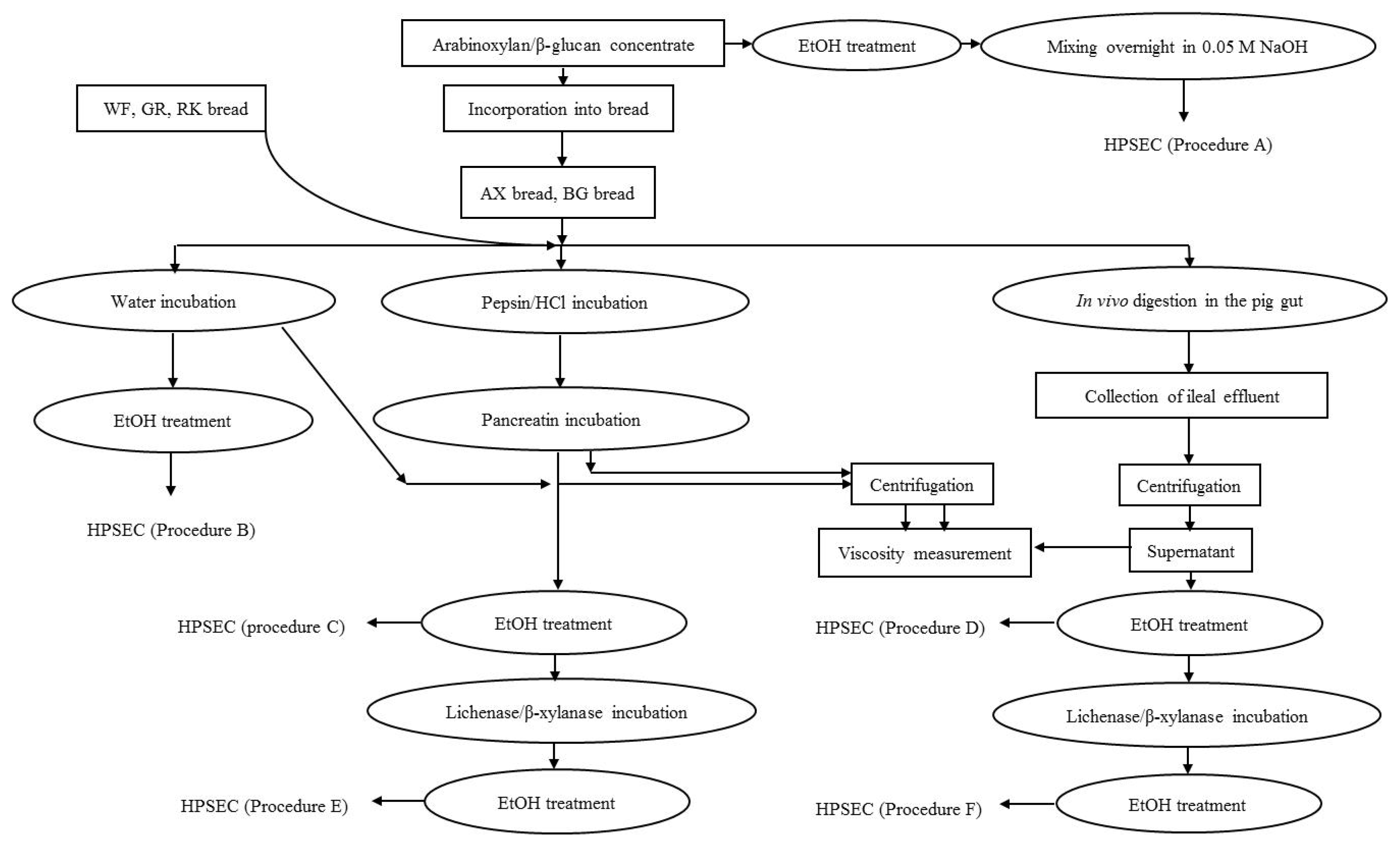
in vivo pig small intestinal digestion using different extraction procedures as illustrated in
Figure 1.
3. Discussion
The potential of a polymer to increase the viscosity, in our case in the gastrointestinal tract, is affected by the molecular weight, concentration, structure of polymer, and possible interaction with other molecules. Hence, not only the properties of the fiber source itself, but also changes that occur during processing and in the digestive tract may affect the physicochemical properties of the fiber. To elucidate this, we have used different procedures to extract and purify extracts of bread and studied fiber modification after baking and passage of the small intestine.
The M
w of alkaline extract of fiber concentrates (602 kDa for arabinoxylan, 1978 kDa for β-glucan) were in line with other investigations. Saulnier
et al.[
3] referred to M
w of wheat arabinoxylan varying from 70 kDa to 655 kDa, while Åman
et al.[
14] characterized oat β-glucan indicating Calcofluor average molecular weight 2060–2300 kDa. The studied fiber concentrates were extracted under mild alkaline conditions and showed a high M
w that was caused by the extractable fiber, but possibly also some starch residues. The main reason of using an alkaline procedure was to prevent aggregation of β-glucan and arabinoxylan [
16] as well as β-elimination or peeling reactions (sequential elimination of the reducing end residues from the polysaccharides chain) than may occur at a high pH [
17]. Although the manufacture of both DF concentrates included a starch elimination step using α-amylase and amyloglucosidase [
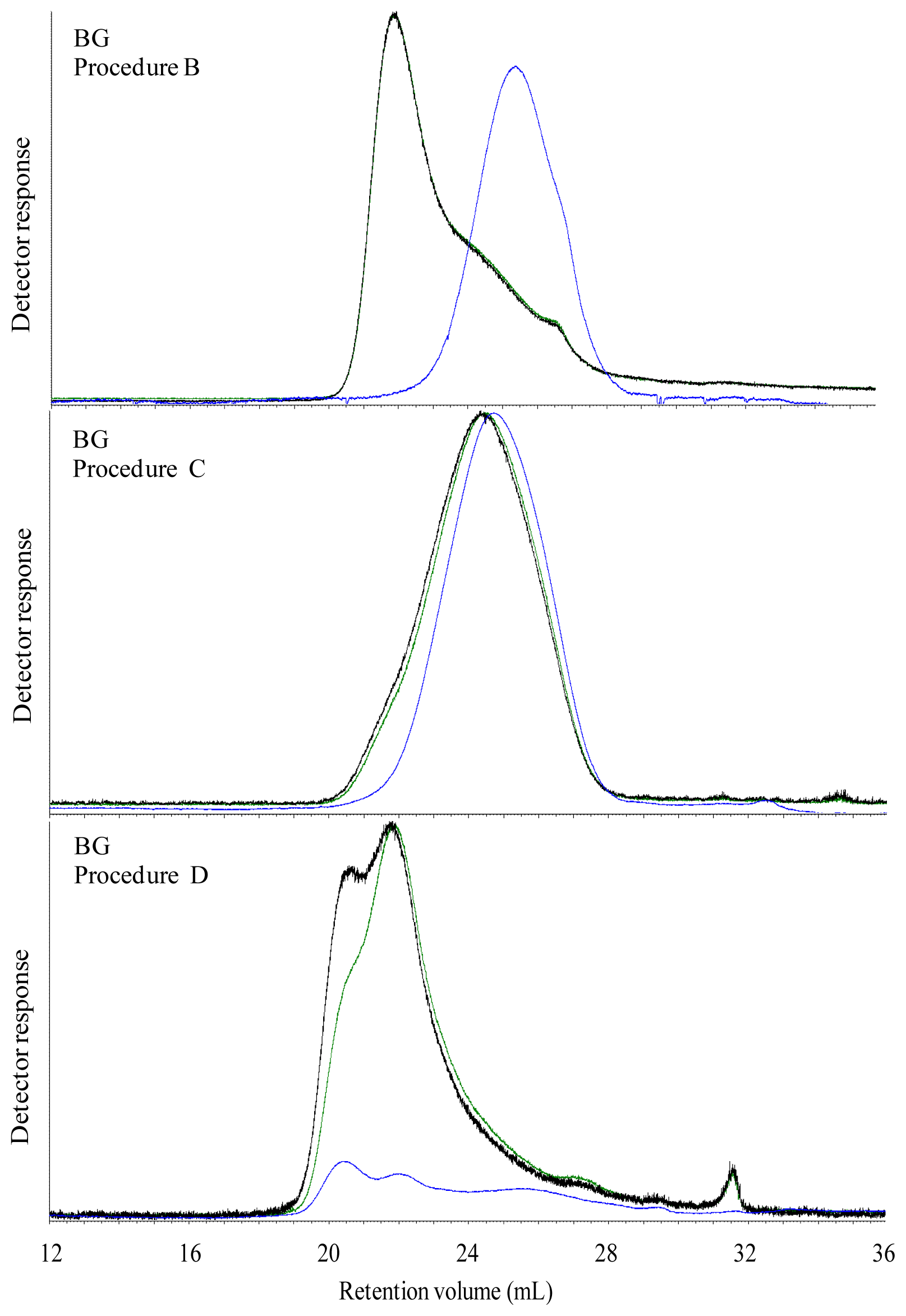
18], particularly the β-glucan concentrate still contained high amounts of starch residues that might have interfered in the M
w determination. On the other hand, the unimodal shape of the BG chromatogram suggests that this was not the case, probably because remaining starch was heavily degraded by the enzymatic treatment during manufacturing.
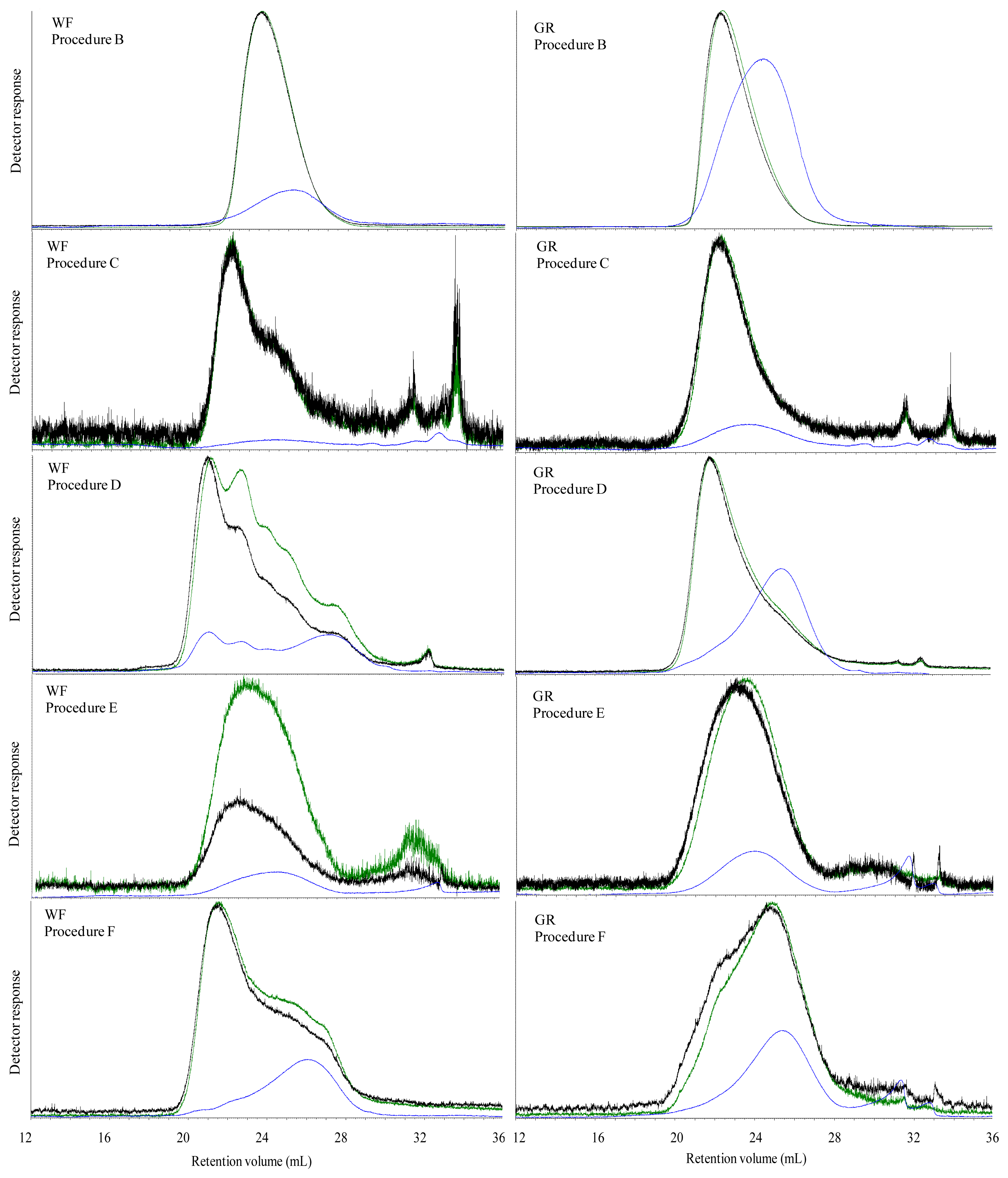
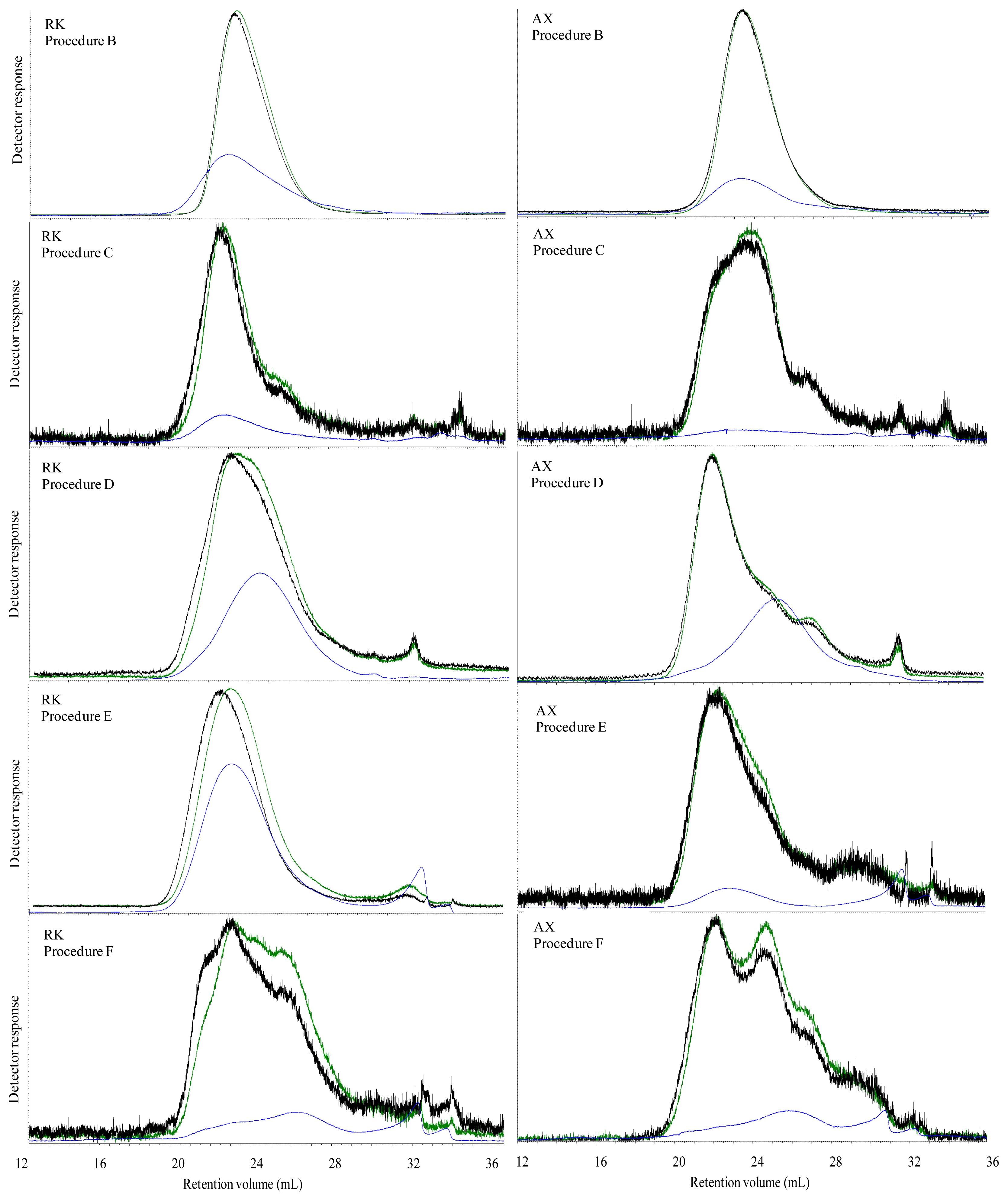
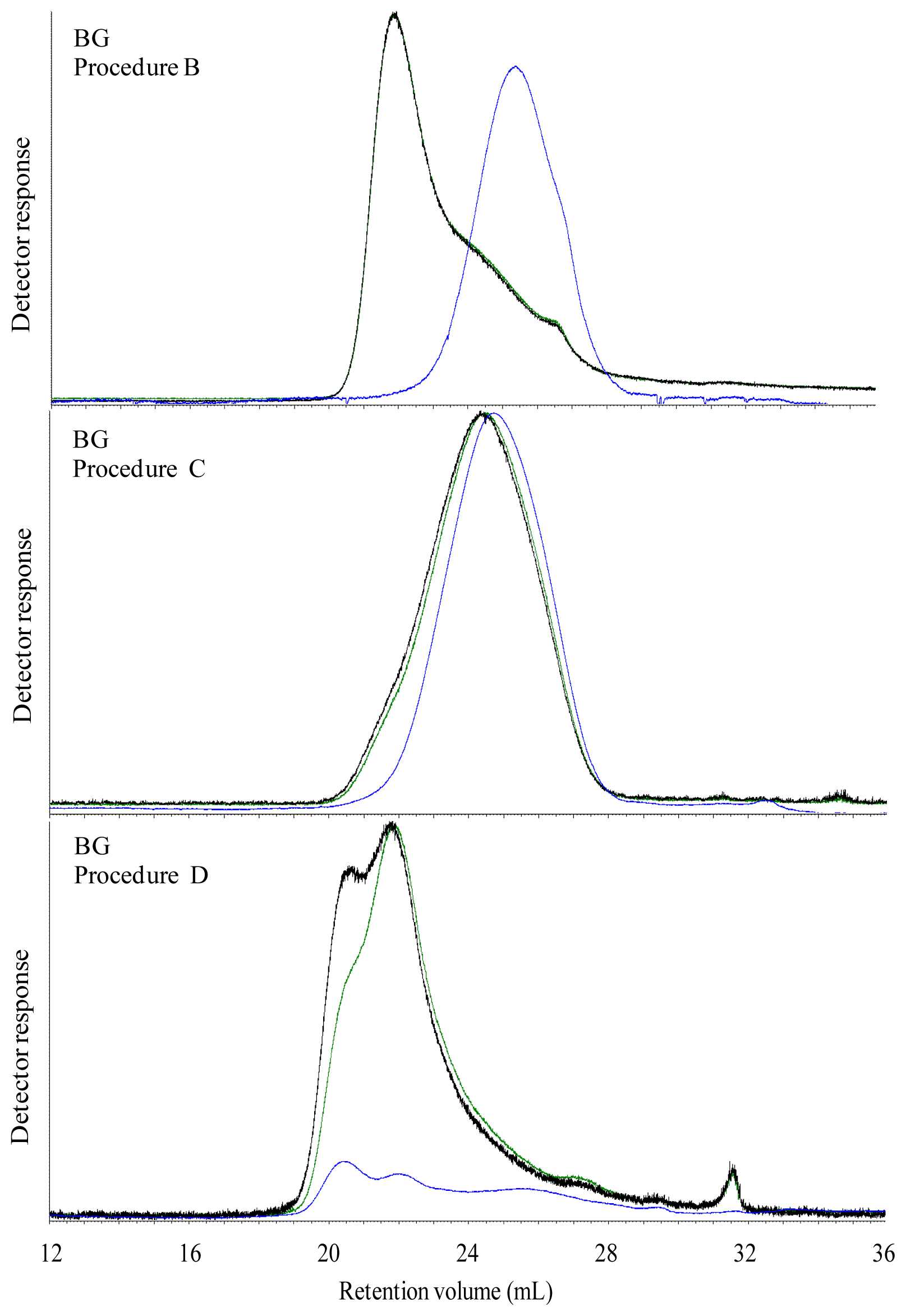
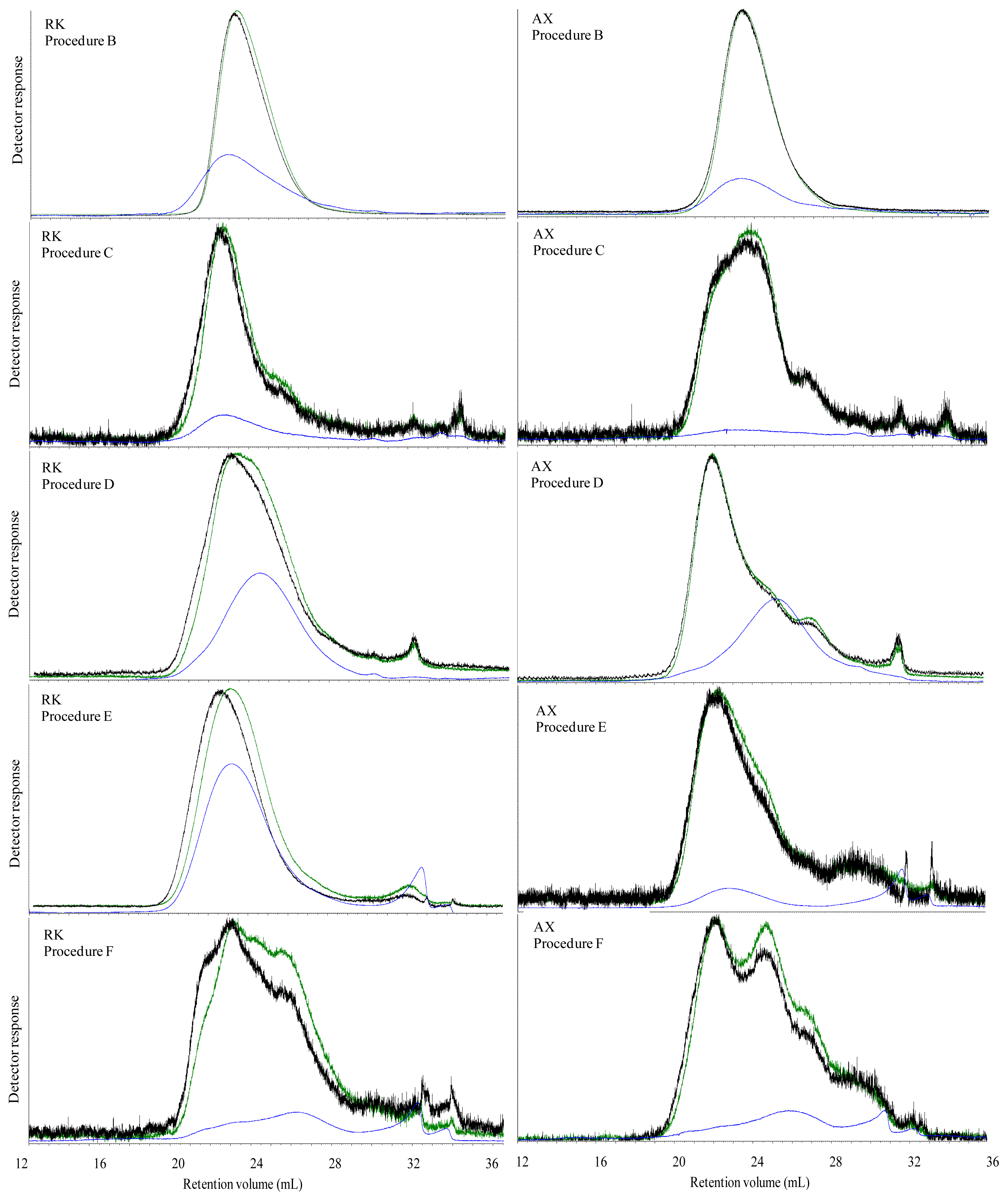
The Mw of carbohydrates was high in all water extracted breads, which most likely was caused by dissolution of both starch and DF. However, in spite of an apparent high Mw of water extractable carbohydrates, the viscosity of the water extract were low in all breads with exception of BG bread. This was possibly due to an insufficient extraction of DF from the bread matrix, indicating that polymers of gelatinized starch and DF present in the grain matrix in the bread were linked with each other or other bread components. The high viscosity of water extract of BG bread was due to the linear structure of β-glucan, which most likely favor its extraction and solubilization in water solution and thereby increasing the viscosity.
The aim of treating the breads with digestive enzymes was to simulate the digestive processes in the gut, but simultaneously this digestion step was expected to deplete DF fraction of the bread for starch and protein. Addition of digestive enzymes did not affect the viscosity of BG bread, whereas the viscosity of WF, GR, RK and AX bread increased after in vitro digestion compared to water extracts. This might suggest that complex structure of arabinoxylan either present in grain matrix or added as concentrate require hydrolysis of starch and protein to enhance its extractability. Moreover, in vitro digestion overall caused a marked decrease in Mw compared to the water extracts in the all of breads. Since none of the enzymes used for in vitro digestion are able to degrade DF polymers, the low Mw of in vitro digested bread compared to the water extracts was possibly due to removal of starch and protein from the matrix. Moreover, the very low Mw of extractable carbohydrates from the AX bread after in vitro digestion (134 kDa) might be due to a low extraction efficiency of branched wheat arabinoxylan polymers (A/X 0.97) in water, resulting also in a low viscosity (2.5 mPa.s) compared to the GR and RK breads. Lichenase purification had a minor impact on the estimated average Mw although it increased in the in vitro digested RK and AX breads. This was consistent with a removal of low Mw components as the proportion being lower than 200 kD decreased for all breads after lichenase treatment but particularly for these two breads.
By inclusion of wheat arabinoxylan concentrate in wheat bread, we observed that the M
w of purified extractable carbohydrates in AX bread was reduced two-fold (307 kDa) compared to the arabinoxylan concentrate (602 kDa). Andersson
et al.[
1] reported that arabinoxylan whole rye flour had a M
w of 4 × 10
4–9 × 10
6 Da with an average of 2 × 10
6 Da, and arabinoxylan whole grain breads showed more narrow M
w of 6 × 10
5–9 × 10
5 Da. This is in the same range as the lichenase purified
in vitro digested RK bread (514 kDa), but much higher than the M
w of in the WF, GR and AX breads. The reduction in M
w of rye and wheat arabinoxylan during bread making is mainly caused by microbial endoxylanases and to minor extent by endogenous endoxylanases present in wheat and rye kernels [
17,
19,
20]. Besides enzymatic degradation, arabinoxylan is sensitive to physical and thermal processing such as mixing and heating, which may lead to degradation of arabinoxylan at 40–70 °C [
17,
21]. Similar to arabinoxylan, β-glucan also depolymerized during bread making. Depolymerization occurs during the mixing and proofing of bread dough with help of added yeast and β-glucanases present in rye flour [
22]. Therefore, along with removal of starch, the M
w of extractable carbohydrates in
in vitro digested BG bread was reduced eight-fold compared to the M
w in the β-glucan concentrate, as previously also demonstrated with barley β-glucan incorporated into wheat bread [
23].
The M
w of extractable carbohydrates was higher in ileal supernatant of GR, RK, AX and BG fed pigs than of the
in vitro digested breads (
Tables 2 and
3). Again, this supports the view that
in vitro digestion did not completely remove starch and protein digestion products from the extract.
In vivo, most starch was digested [
24] leaving a very low concentration in the ileal supernatant except for the BG fed pigs.
In our study, there was a marked change in the elution profile of components after
in vivo digestion compared to
in vitro digestion particularly in the lichenase purified samples. This resulted in the formation of three populations in the RK and AX extracts and two populations in WF and GR extracts. In spite of the change in elution profile, the M
w of the lichenase purified
in vitro and
in vivo digested breads remained almost unchanged (
Tables 4 and
5). The upper gastrointestinal tract is inhabited by protozoa, fungi and cellulotytic and xylanolytic bacteria species, which are able to degrade a mixture of homo- and heteropolymer substrates, including xyloglucan, β-glucan, glucuronomanan, arabinoxylan and xylan [
25,
26]. A range of studies have also shown that both quantitatively and quantitatively β-glucan and arabinoxylan may be degraded in the distal small intestine of pigs and humans [
27–
31], but most studies points toward arabinoxylan being less susceptible towards degradation than β-glucan. This is due to the more complex structure of arabinoxylan, which presumably prevents disintegration by microbial enzymes, since seven different enzymes are required to cleave the arabinoxylan molecule [
32]. Hence, the higher concentration of arabinoxylan in ileal supernatant of the AX fed pigs compared to the GR and RK fed pigs could be brought about by two counteracting mechanisms; higher solubility of the wheat arabinoxylan concentrate added to the wheat bread than of the arabinoxylan located in the cell walls of rye in the GR and RK bread, or lower degree of degradation of the added wheat arabinoxylan compared to the two rye breads. Although we did not find significant differences in the ileal digestibility of arabinoxylan between the breads, the ileal digestibility of the AX bread was numerically lower [
24]. Furthermore, as the arabinoxylan in the AX bread was highly branched (A/X 0.97) compared to the arabinoxylan found in the two rye breads (A/X 0.70–0.74), we may expect that this branching pattern (
i.e., high xylose substitution by arabinose) protect the xylose chain toward extensive degradation by microbial enzymes and thereby allow a high concentration of arabinoxylan in the ileal supernatant, which also resulted in a significantly higher viscosity. In contrast to arabinoxylan, the linear structure of β-glucan was easily degraded by the microbial enzymes that led to a reduction in the ileal viscosity of BG bread. Nevertheless, the reduced M
w of extractable carbohydrate in ileal supernatants was not related to the ileal viscosity among all breads.
4. Experimental Section
4.1. Breads and Diets
The investigation included a control bread baked on white wheat flour (WF) and four iso-DF breads: two commercial whole grain rye breads without (GR) and with grain kernels (RK) and two experimental wheat flour based breads supplemented with arabinoxylan concentrate (AX) and β-glucan concentrate (BG), respectively. WF, GR and RK were commercial breads produced by Schulstad Lantmännen A/S (Hammerholmen, Hvidovre, Denmark) with the commercial names: Hvede Toast, Mørkt Rugbrød and Multikernerugbrød, respectively. The ingredients for WF were in decreasing order; wheat flour (68%), water, yeast, sugar, salt, vinegar, canola oil, emulsifier (E471, E472e), rye flour, barley malt flour, flour treatment agent (ascorbic acid); GR was baked on; rye whole meal, water, rye sourdough, rye bread crumps, salt, vinegar, dried sourdough rye, canola oil, yeast, barley flour, and RK was produced from: rye kernels (49%), water, rye sourdough, rye bread crumbs, wheat meal, salt, yeast, vinegar, dried sourdough rye, canola oil, rye four, barley flour. BG and AX breads were experimental breads baked at a local bakery (Konditor-Bageren, Ørum, Denmark). The ingredients for AX were; white wheat flour, arabinoxylan concentrate (wheat arabinoxylan, Manildra Group Ltd, Nowra, Australia), sugar, salt, margarine, and yeast. The BG bread was prepared from white wheat flour, β-glucan concentrate (oat β-glucan, known commercially as PromOat, BioVelop AB, Kimstad, Sweden), Vitacel WF 600 (16.9% of arabinoxylan, J. Rettenmaier & Söhne GmbH, Rosenberg, Germany), wheat gluten, sugar, salt, margarine, and yeast. The breads were cut into pieces, frozen at −20 °C and following mincing partly frozen. The minced breads were mixed with digestibility markers (chromic oxide and Celite®), vitamins, minerals and in case of AX bread with whey protein concentrate (Lacprodan 87; Arla Foods Ingredients Amba, Viby J, Denmark), in order to adjust the protein content of the diets. The feed was portioned into plastic bags containing 575 g DM per meal and stored at −20 °C until consumption.
4.2. Animal Study
The pigs used in the study were crossbreeds of Duroc × Danish Landrace × Yorkshire obtained from the swineherd at Aarhus University, Department of Animal Science, Denmark. The animal experiment was conducted according to protocols approved by the Danish Animal Experiments Inspectorate and complied with the guidelines of the Danish Ministry and Justice concerning animal experimentation and care of animals under study.
Five female pigs with average weight of 58 ± 2.8 kg were fitted surgically with a permanent simple T-cannula 15 cm anterior to the ileal-caecal junction following the procedure of Jørgensen
et al.[
33] and allowed to recover until start of the study. The pigs were fed each experimental diet for 1 week in a Latin Square design. Prior to the feeding, the diets were thawed. Total daily feed intake was 1725 g DM/d and drinking water was provided
ad libitum.
On day 6 and 7 ileal effluent was collected continuously for 4 h after the morning meal. Ileal effluent was collected in 6 mm × 20 mm polyamide autoclave bags (Buch and Holm, Herlev, Denmark) attached to the ileum cannula with a plastic zip fastener. Two to three drops of an aqueous solution of 0.2% NaN3 (Sigma-Aldrich, St. Louis, MO, USA) was added to the collection bags to prevent microbial activity. Forty grams of material from each day of collection was centrifuged (10,000× g, 4 °C, 20 min) in 50 mL tubes. Following the supernatant from the 2 collection days was pooled into 10 mL tubes and stored at −20 °C until further analysis.
Results on carbohydrate digestibility and physicochemical properties of ileal effluent are described elsewhere [
24].
4.3. Sample Preparation for HPSEC Analysis
The fiber concentrates, breads and the supernatants of ileal digesta were analyzed as illustrated in
Figure 1 and described in detail below.
4.3.1. Solubilization of DF from the Fiber Concentrates
Extraction of DF from the fiber concentrates was done according to the procedure of Suortti [
4] illustrated in
Figure 1 as Procedure A. One hundred milligrams of arabinoxylan and β-glucan concentrate were weighed into 50 mL plastic tubes. Five mL of 50% ethanol was added, then the sample was mixed, placed in a boiling water bath for 15 min and cooled. Additionally, 5 mL of 50% ethanol was added, mixed, centrifuged (1000×
g) and the precipitate collected. Twenty milliliters of 0.05 M aqueous NaOH solution was added to the precipitate and mixed overnight at room temperature and filtered through a 0.2 μm minisart filter (Sartorius Stedim Biotech Gmbh, Germany) prior to HPSEC analysis.
4.3.2. Water Extraction and in vitro Digestion of Breads
The breads were extracted in azid-containing water or subjected to
in vitro enzymatic digestion according to Procedures B and C, as illustrated in
Figure 1.
Prior to extraction, the breads were minced for 1 min using a hand blender (Braun, Kronberg, Germany) and then the material was weighed in triplicate into a tube adjusted to a DM content of 13% in the solution, in order to mirror the approximate final DM content of ileal digesta. For in vitro digestion (Procedure C), the breads were initially incubated with a 11.68 mL pepsin solution (12 mg, 3.260 U/mL, EC.3.4.2.3.1., Merck, Darmstadt, Germany) in 0.1 M HCl in a shaking water bath (150 strokes/min) at 39 °C for 2 h. Following pH was adjusted to 7.0 by addition of a 6.54 mL solution of porcine pancreatin (50 mg, 8 × USP, EC. 232-468-9, Sigma-Aldrich, St.Louis, MO, USA) in 1 M NaHCO3, and incubated in the same water bath for further 2 h. Water extraction (Procedure B) followed the same procedure except that the enzyme solutions were replaced with an aqueous solution of 0.02% NaN3 (Sigma-Aldrich, St. Louis, MO, USA). After the water and in vitro incubations, the bread mixtures were centrifuged and 5 mL of supernatants were transferred to a 50 mL plastic tube and treated with ethanol: Five milliliter of 96% ethanol was added to the supernatant, mixed, and placed in a boiling water bath for 15 min, cooled and supernatant was discarded. Another 5 mL of 50% ethanol was added, the sample was mixed, centrifuged (1000× g) and the precipitate collected. Five milliliters of 0.02% NaN3 was added to the precipitate and the mixture was boiled for 10 min and cooled. The samples were filtered and injected into the HPSEC system.
For a parallel set of samples the precipitate after in vitro digestion and subsequent ethanol treatment was further treated with either lichenase or β-xylanase in order to purify the arabinoxylan and the β-glucan, respectively (Procedure E). The precipitate of the BG bread was incubated with 50 μL of β-xylanase (from Trichoderma viride, 2300 U/mL, Megazyme International Ireland, Wicklow, Ireland) in 5 mL of 0.1 M sodium acetate buffer (pH 5) at 50 °C for 2 h, whereas precipitates of WF, GR, RK and AX breads were incubated with 100 μL of lichenase (from Bacillus subtilis, 1000 U/mL, Megazyme International Ireland) in 5 mL of 0.1 M sodium acetate and 2 mL of 0.5 M Na2HPO4 (added to increase pH to 6.5–6.7) at 60 °C for 2 h in the water bath. During the incubation, the samples were occasionally stirred. Afterwards, samples from both incubations were cooled and mixed with 40 mL of 96% ethanol in an ice box for 30 min. The samples were centrifuged (1000× g), and supernatants were discarded, while precipitates were re-dissolved in 5 mL of 0.02% aqueous NaN3 solution in a boiling water bath for 10 min. After cooling, the samples were filtered and analyzed by HPSEC.
The step of ethanol treatment in our procedures was previously described by Andersson
et al.[
1].
4.3.3. Isolation and Purification of DF from Ileal Effluent
The ileal supernatant was thawed and 5 mL was transferred to a 50 mL plastic tube. Five milliliters of 96% ethanol and 5 mL of 50% ethanol were added to the supernatant and treated according to the ethanol treatment described in section 4.3.2. After removal of the liquid phase, 5 mL of 0.02% of NaN3 was added to the precipitate and the mixture was boiled for 10 min and cooled. The sample was filtered and injected to the HPSEC (Procedure D). For a parallel set of samples, ileal supernatant after the ethanol treatment was further treated with lichenase or β-xylanase for purification of arabinoxylan and β-glucan (Procedure F) as described in section 4.3.2.
4.3.4. Side Activity of Lichenase and β-Xylanase
Lichenase and β-xylanase were tested for side activity on pure standards of arabinoxylan (wheat arabinoxylan, high viscosity 47 cSt., Megazyme International Ireland) and β-glucan (oat β-glucan, high viscosity 69 cSt., Megazyme International Ireland), respectively under conditions as described in section 4.3.2.
4.4. Analysis
4.4.1. Carbohydrates of Fiber Concentrates, Breads and Ileal Supernatants
The starch and total content of NSP of the fiber concentrates, the breads and the corresponding digesta supernatants were measured as described by Bach Knudsen [
34] with the modification that the polysaccharides in starch free residue were hydrolyzed with 2 M H
2SO
4 for 1 h instead of 1 M H
2SO
4 for 2 h. The content of β-glucan in breads was measured by the enzymatic-colorimetric method of McCleary and Glennie-Holmes [
35], whereas the β-glucan in β-glucan concentrate and ileal supernatants was determined as non-cellulosic (NCP) glucose by hydrolysis without prior solubilization with 12 M H
2SO
4. The arabinoxylan content in fiber concentrates, breads and ileal supernatants was calculated as the sum of arabinose and xylose content obtained by the NSP method. A/X ratio of arabinoxylan was calculated as a quotient of arabinose and xylose content.
4.4.3. Viscosity of Bread Extracts and Ileal Effluents
The bread after water incubation and in vitro and in vivo digestion were centrifuged (10,000× g, 4 °C, 20 min) in 50 mL tubes and the viscosity of the supernatants was measured in a Brookfield DV-II cone/plate viscometer (Brookfield Engineering Laboratories Inc., Stoughton, MA, USA) at 39 °C at shear rates between 2.25 and 450 s−1. The cone of 0.8° angle had a diameter of 4.8 cm. Values of viscosity at shear rate 45 s−1 were used.
4.5. Statistical Analyses
The M
w, proportion of M
w <200 kDa and >200 kDa of two different extraction methods on five breads were analyzed using a two-way analysis-of-variance model (
Table 2). The same response variables obtained from ethanol treated ileal contents were analyzed using a mixed model (
Table 3). Moreover, the response variables of the lichenase purified arabinoxylan in the
in vitro digested WF, GR, RK and AX breads and their ileal supernatant counterparts were analyzed using a one-way analysis-of-variance model (
Tables 4 and
5). Log values of viscosity of water extracts and
in vitro digested breads were calculated by one-way analysis, and of ileal supernatants by a mixed model (
Table 6).
All statistical analysis was performed using Statistical Analysis Software (SAS Institute Inc., Cary, NC, USA). All data were expressed as least squares means with differences considered statistically significant at p < 0.05.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}