Isolation and Characterization of Saponin-Producing Fungal Endophytes from Aralia elata in Northeast China

Abstract
:1. Introduction
2. Results and Discussion
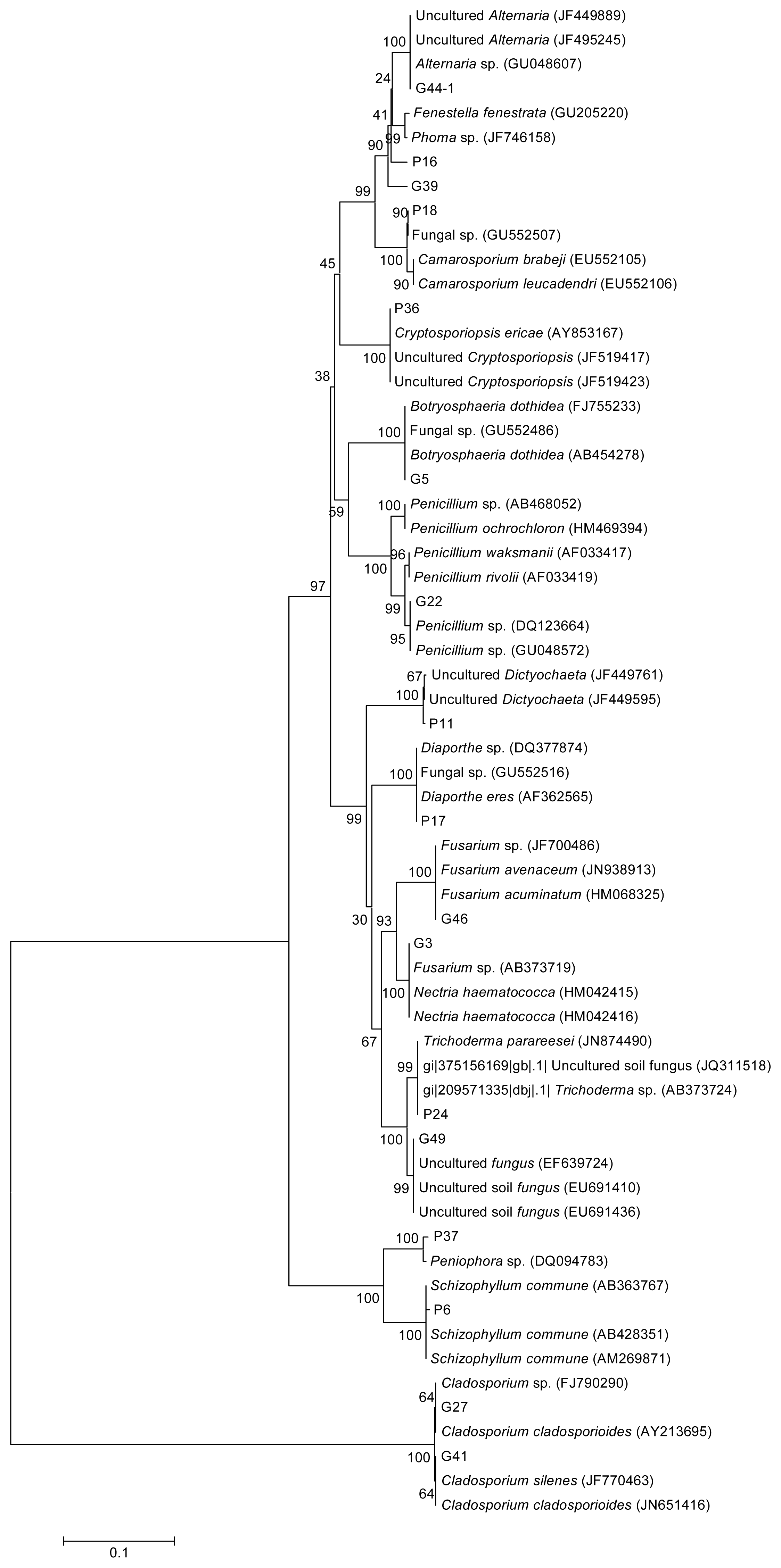
2.1. Strains Identification and Phylogenetic Analysis
2.2. Analysis of Triterpenoid Saponins and Antimicrobial Activity
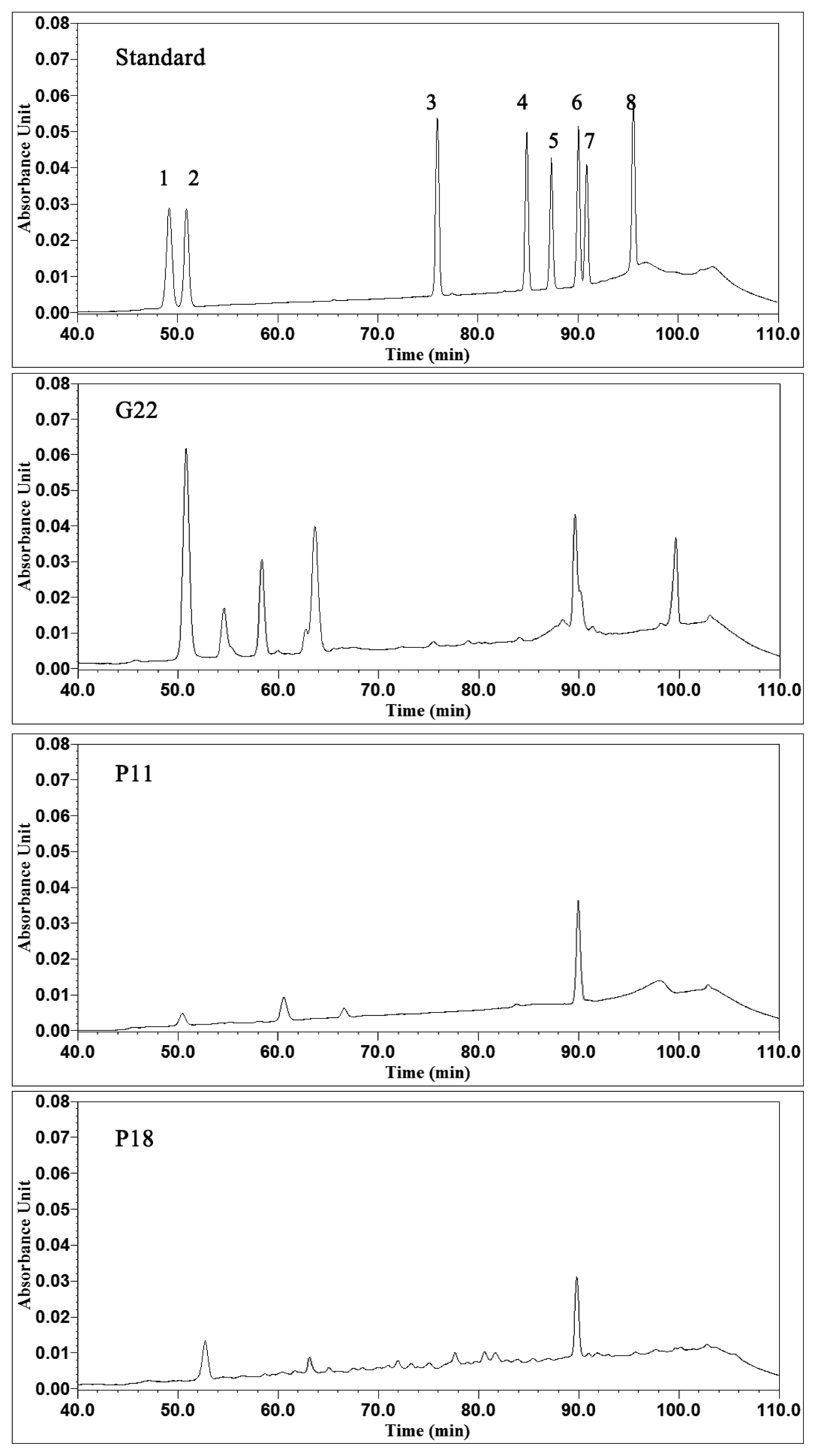
2.3. Ginsenosides Analyses
3. Experimental Section
3.1. Sampling and Isolation
3.2. DNA Extraction and PCR Amplification of the 28S rRNA Gene
3.3. Phylogenetic Analysis and Nucleotide Sequence Accession Numbers
3.4. Determination of Triterpenoid Saponins
3.5. Antimicrobial Activity of the Representative Strains
3.6. Ginsenosides Analyses
4. Conclusions
Acknowledgments
References
- Huang, W.Y.; Cai, Y.Z.; Hyde, K.D.; Corke, H.; Sun, M. Biodiversity of endophytic fungi associated with 29 traditional Chinese medicinal plants. Fungal Divers 2008, 33, 61–75. [Google Scholar]
- Hyde, K.D.; Soytong, K. The fungal endophyte dilemma. Fungal Divers 2008, 33, 163–173. [Google Scholar]
- Koukol, O. New species of Chalara occupying coniferous needles. Fungal Divers 2011, 49, 75–91. [Google Scholar]
- Li, H.; Shen, M.; Zhou, Z.; Li, T.; Wei, Y.; Lin, L. Diversity and cold adaptation of endophytic fungi from five dominant plant species collected from the Baima Snow Mountain, Southwest China. Fungal Divers 2012, 54, 79–86. [Google Scholar]
- Alberton, O.; Kuyper, T.W.; Summerbell, R.C. Dark septate root endophytic fungi increase growth of Scots pine seedlings under elevated CO2 through enhanced nitrogen use efficiency. Plant Soil 2010, 328, 459–470. [Google Scholar]
- Saikkonen, K.; Saari, S.; Helander, M. Defensive mutualism between plants and endophytic fungi? Fungal Divers 2010, 41, 101–113. [Google Scholar]
- Osono, T.; Hirose, D. Ecology of endophytic fungi associated with leaf litter decomposition. In Applied Mycology; Rai, M., Bridge, P.D., Eds.; CAB International: Cambridge, MA, USA, 2009; pp. 92–109. [Google Scholar]
- Schulz, B.; Boyle, C.; Draeger, S.; Römmert, A.; Krohn, K. Endophyticfungi: A source of novel biologically active secondary metabolites. Mycol. Res 2002, 106, 996–1004. [Google Scholar]
- Rivera-Orduña, F.N.; Suarez-sanchez, R.A.; Flores-Bustamante, Z.R.; Gracida-Rodriguez, J.N.; Flores-Cotera, L.B. Diversity of endophytic fungi of Taxus globosa (Mexican yew). Fungal Divers 2011, 47, 65–74. [Google Scholar]
- Kusari, S.; Verma, V.C.; Lamshoeft, M.; Spiteller, M. An endophytic fungus from Azadirachta indica A. Juss. that produces azadirachtin. World J. Microbiol. Biotechnol 2012, 28, 1287–1294. [Google Scholar]
- Paul, N.C.; Kim, W.K.; Woo, S.K.; Park, S.; Yu, S.H. Fungal endophytes in roots of Aralia species and their antifungal activity. Plant Pathol. J 2007, 23, 287–294. [Google Scholar]
- Sakai, S.; Katsumata, M.; Satoh, Y.; Nagasao, M.; Miyakoshi, M.; Ida, Y.; Shoji, J. Oleanolic acid saponins from root bark of Aralia elata. Phytochemistry 1994, 35, 1319–1324. [Google Scholar]
- Mujoo, K.; Haridas, V.; Hoffmann, J.J.; Wächter, G.A.; Hutter, L.K.; Lu, Y.; Blake, M.E.; Jayatilake, G.S.; Bailey, D.; Mills, G.B.; et al. Triterpenoid saponins from Acacia victoriae (Bentham) decrease tumor cell proliferation and induce apoptosis. Cancer Res 2001, 61, 5486–5490. [Google Scholar]
- Fujimoto, J.; Sakaguchi, H.; Alki, I.; Toyoki, H.; Khatun, S.; Tamaya, T. Inhibitory effect of ginsenoside-Rb2 on invasiveness of uterine endometrial cancer cells to the basement membrane. Eur. J. Gynaecol. Oncol 2001, 22, 339–341. [Google Scholar]
- Zhao, Y.-L.; Cai, G.-M.; Hong, X.; Shan, L.-M.; Xiao, X.-H. Anti-hepatitis B virus activities of triterpenoid saponin compound from Potentilla anserine L. Phytomedicine 2008, 15, 253–258. [Google Scholar]
- Sen, S.; Chakraborty, R.; De, B.; Mazumder, J. Plants and phytochemicals for peptic ulcer: An overview. Pharmacognosy Rev 2009, 3, 270–279. [Google Scholar]
- Davis, E.C.; Shaw, A.J. Biogeographic and phylogenetic patterns in diversity of liverwort-associated endophytes. Am. J. Bot 2008, 95, 914–924. [Google Scholar]
- Prada, H.; Avila, L.; Sierra, R.; Bernal, A.; Restrepo, S. Morphological and molecular characterization of the antagonistic interaction between the endophyte Diaporthe sp. isolated from frailejon (Espeletia sp.) and the plant pathogen Phytophthora infestans. Rev. Iberoam. Micol 2009, 26, 198–201. [Google Scholar]
- Maehara, S.; Ikeda, M.; Haraguchi, H.; Kitamura, C.; Nagoe, T.; Ohashi, K.; Shibuya, H. Microbial conversion of curcumin into colorless hydroderivatives by the endophytic fungus Diaporthe sp. associated with Curcuma longa. Chem. Pharm. Bull 2011, 59, 1042–1044. [Google Scholar]
- Bungihan, M.E.; Tan, M.A.; Kitajima, M.; Kogure, N.; Franzblau, S.G.; Dela Cruz, T.E.; Takayama, H.; Nonato, M.G. Bioactive metabolites of Diaporthe sp. P133, an endophytic fungus isolated from Pandanus amaryllifolius. J. Nat. Med 2011, 65, 606–609. [Google Scholar]
- Sebastianes, F.L.S.; Cabedo, N.; Aouad, N.E.; Valente, A.M.M.P.; Lacava, P.T.; Azevedo, J.L.; Pizzirani-Kleiner, A.A.; Cortes, D. 3-Hydroxypropionic acid as an antibacteria agent from endophytic fungi Diaporthe phaseolorum. Curr. Microbiol. 2012. [Google Scholar] [CrossRef]
- Xiang, Y.; Lu, A.; Wu, W. Identification of Taxus cuspidata sieb. et Zucc. endophytic fungi-new species, species known and their metabolite. J. For. Res 2003, 14, 290–294. [Google Scholar]
- Dey, P.; Banerjee, J.; Maiti, M.K. Comparative lipid profiling of two endophytic fungal isolates—Colletotrichum sp. and Alternaria sp. having potential utilities as biodiesel feedstock. Bioresource Technol 2011, 102, 5815–5823. [Google Scholar]
- Chen, G.; Zhu, Y.; Wang, H.Z.; Wang, S.J.; Zhang, R.Q. The metabolites of a mangrove endophytic fungus, Penicillium thomi. J. Asian Nat. Prod. Res 2007, 9, 159–164. [Google Scholar]
- Guo, Z.; Cheng, F.; Zou, K.; Wang, J.; She, Z.; Lin, Y. Secondary metabolites from the mangrove endophytic fungus Penicillium sp. (SBE-8). Nat. Prod. Commun 2009, 4, 1481–1483. [Google Scholar]
- Hamayun, M.; Khan, S.A.; Iqbal, I.; Ahmad, B.; Lee, I.J. Isolation of a gibberellin-producing fungus (Penicillium sp. MH7) and growth promotion of Crown daisy (Chrysanthemum coronarium). J. Microbiol. Biotechnol 2010, 20, 202–207. [Google Scholar]
- Ahmad, N.; Hamayun, M.; Khan, S.A.; Khan, A.L.; Lee, I.J.; Shin, D.H. Gibberellin-producing endophytic fungi isolated from Monochoria vaginalis. J. Microbiol. Biotechnol 2011, 20, 1744–1749. [Google Scholar]
- Redecke, D. Specific PCR primers to identify arbuscular mycorrhizal fungi within colonized roots. Mycorrhiza 2000, 10, 73–80. [Google Scholar]
- Yang, H.; Wu, H.; Wang, X.; Cui, Z.; Li, Y. Selection and characteristics of a switchgrass-colonizing microbial community to produce extracellular cellulases and xylanases. Bioresour. Technol 2011, 102, 3546–3550. [Google Scholar]
- GenBank. Basic Local Alignment Search Tool. Available online: http://blast.ncbi.nlm.nih.gov/Blast.cgi accessed on 12 June 2012.
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likehood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol 2011, 28, 2731–2739. [Google Scholar]
- Liu, H.G.; Li, T.; Zhao, Y.L.; Zhang, J.; Wang, Y.Z. Determination of some metabolites of Cordyceps sobolifera. Afr. J. Microbiol. Res 2011, 5, 5518–5522. [Google Scholar]
- Hormazabal, E.; Piontelli, E. Endophytic fungi from chilean native gymnosperms: Antimicrobial activity against human and phytopathogenic fungi. World J. Microbiol. Biotechnol 2009, 25, 813–819. [Google Scholar]



{kind=link}
{kind=link}
| Strain ID (Strains) | Accession no. | Closest species (Accession no.) | Coverage (%) |
|---|---|---|---|
| P17 (24) | JQ807963 | Fungal sp. (GU552516) | 100 |
| Diaporthe sp. (DQ377874) | 100 | ||
| Diaporthe eres (AF362565) | 100 | ||
| G44-1 (12) | JQ807974 | Uncultured Alternaria (JF495245) | 100 |
| Uncultured Alternaria (JF449889) | 100 | ||
| Alternaria sp. (GU048607) | 100 | ||
| G22 (8) | JQ807910 | Penicillium sp. (GU048572) | 100 |
| Penicillium sp. (DQ123664) | 100 | ||
| Penicillium rivolii (GU033419) | 99.3 | ||
| G27 (8) | JQ807969 | Cladosporium sp. (FJ790290) | 100 |
| Cladosporium cladosporioides (AY213695) | 100 | ||
| Passalora fulva (AB100653) | 100 | ||
| P6 (6) | JQ807984 | Schizophyllum commune (AM269871) | 99.8 |
| Schizophyllum commune (AB428351) | 99.8 | ||
| Schizophyllum commune (AB363767) | 99.8 | ||
| P11(5) | JQ807967 | Uncultured Dictyochaeta (JF449595) | 99.3 |
| Uncultured Dictyochaeta (JF449592) | 98.8 | ||
| Dictyochaeta simplex (AF178559) | 98 | ||
| P18(5) | JQ807982 | Fungal sp. (GU552507) | 100 |
| Camarosporium leucadendri (EU552106) | 99.7 | ||
| Camarosporium brabeji (EU552105) | 99.7 | ||
| G3 (5) | JQ807935 | Nectria haematococca (HM042416) | 100 |
| Nectria haematococca (AB373719) | 100 | ||
| G41 (4) | JQ807972 | Cladosporium cladosporioides (JN651416) | 100 |
| Cladosporium silenes (JF770463) | 100 | ||
| Uncultured Cladosporium (JF449832) | 100 | ||
| G46 (4) | JQ807956 | Fusarium avenaceum (JN938913) | 100 |
| Fusarium sp. (JF700486) | 100 | ||
| Fusarium avenaceum (HM068325) | 100 | ||
| P37 (3) | JQ807942 | Peniophora sp. (HM595610) | 99.8 |
| Peniophoraceae sp. (AB576771) | 99.8 | ||
| Peniophoraceae sp. (DQ094783) | 99.8 | ||
| P24(3) | JQ807922 | Uncultured soil fungus (JQ311518) | 99.5 |
| Trichoderma parareesei (JN874490) | 99.3 | ||
| Trichoderma sp. (AB373724) | 99.1 | ||
| P16 (3) | JQ807976 | Uncultured Pleosporales (JF691161) | 100 |
| Uncultured Epicoccum (JF449817) | 100 | ||
| Uncultured Epicoccum (JF449816) | 100 | ||
| P36 (2) | JQ807968 | Uncultured Cryptosporiopsis (JF519417) | 100 |
| Uncultured Cryptosporiopsis (JF519423) | 99.8 | ||
| Cryptosporiopsis ericae (AY853167) | 99.8 | ||
| G5 (2) | JQ807983 | Fungal sp. (GU552486) | 100 |
| Botryosphaeria dothidea (AB454278) | 100 | ||
| Botryosphaeria dothidea (FJ755233) | 100 | ||
| G49 (1) | JQ807918 | Uncultured soil fungus (EU691410) | 100 |
| Uncultured soil fungus (EU691436) | 100 | ||
| Uncultured fungus (EF639724) | 100 | ||
| G39 (1) | JQ807981 | Corynespora smithii (GU323201) | 94.6 |
| Fenestella fenestrate (GU205220) | 94.6 | ||
| Phoma sp. (JF746158) | 94.3 |
| Isolate ID | Mean ± Stdev (mg/mL) (p < 0.05) | Expected species |
|---|---|---|
| G22 | 2.049 ± 0.044 a | Penicillium sp. |
| P11 | 0.162 ± 0.004 b | Dictyochaeta sp. |
| P18 | 0.156 ± 0.006 b | Camarosporium leucadendri |
| G27 | 0.131 ± 0.003 c | Cladosporium sp. |
| P6 | 0.120 ± 0.003 cd | Schizophyllum commune |
| G49 | 0.113 ± 0.005 cd | Uncultured soil fungus |
| P37 | 0.109 ± 0.006 d | Peniophora sp. |
| P17 | 0.084 ± 0.005 e | Diaporthe sp. |
| P16 | 0.066 ± 0.005 ef | Epicoccum sp. |
| G41 | 0.066 ± 0.006 efg | Cladosporium cladosporioides |
| G46 | 0.065 ± 0.003 efg | Fusarium avenaceum |
| P24 | 0.060 ± 0.006 fg | Trichoderma parareesei |
| G44-1 | 0.059 ± 0.002 fg | Alternaria sp. |
| G39 | 0.048 ± 0.001 gh | Corynespora smithii |
| G5 | 0.038 ± 0.004 h | Botryosphaeria dothidea |
| G3 | 0.032 ± 0.005 h | Nectria haematococca |
| P36 | 0.030 ± 0.002 h | Cryptosporiopsis sp. |
| Test strains | Representative endophytic strains | ||
|---|---|---|---|
| G22 | P11 | P18 | |
| Staphylococcus aureus ACCC10499 | ++ | + | − |
| Bacillus subtitis ACCC10243 | − | − | − |
| Klebsiella pneumoniae ACCC10498 | − | +++ | + |
| Pseudomonas aeruginosa ACCC10500 | − | − | − |
| Phytophthora cactorum ACCC36421 | − | − | − |
| Rhizoctonia solani ACCC36233 | − | + | − |
| Aspergillus niger ACCC30005 | − | − | − |
| Fusarium sporotrichioides | − | + | − |
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Wu, H.; Yang, H.; You, X.; Li, Y. Isolation and Characterization of Saponin-Producing Fungal Endophytes from Aralia elata in Northeast China. Int. J. Mol. Sci. 2012, 13, 16255-16266. https://doi.org/10.3390/ijms131216255
Wu H, Yang H, You X, Li Y. Isolation and Characterization of Saponin-Producing Fungal Endophytes from Aralia elata in Northeast China. International Journal of Molecular Sciences. 2012; 13(12):16255-16266. https://doi.org/10.3390/ijms131216255
Chicago/Turabian StyleWu, Hao, Hongyan Yang, Xiangling You, and Yuhua Li. 2012. "Isolation and Characterization of Saponin-Producing Fungal Endophytes from Aralia elata in Northeast China" International Journal of Molecular Sciences 13, no. 12: 16255-16266. https://doi.org/10.3390/ijms131216255