Effects of Curcumin on the Proliferation and Mineralization of Human Osteoblast-Like Cells: Implications of Nitric Oxide

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
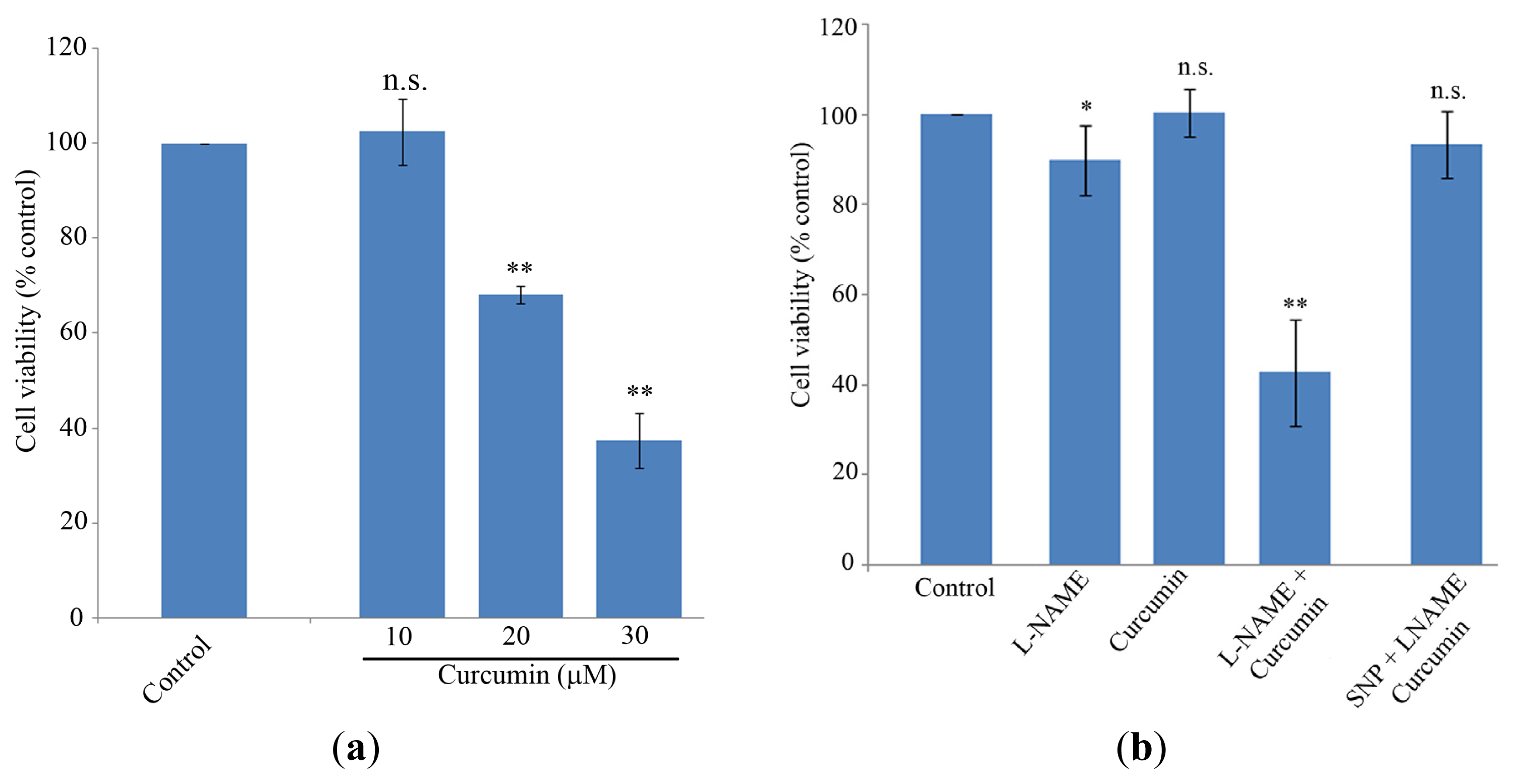
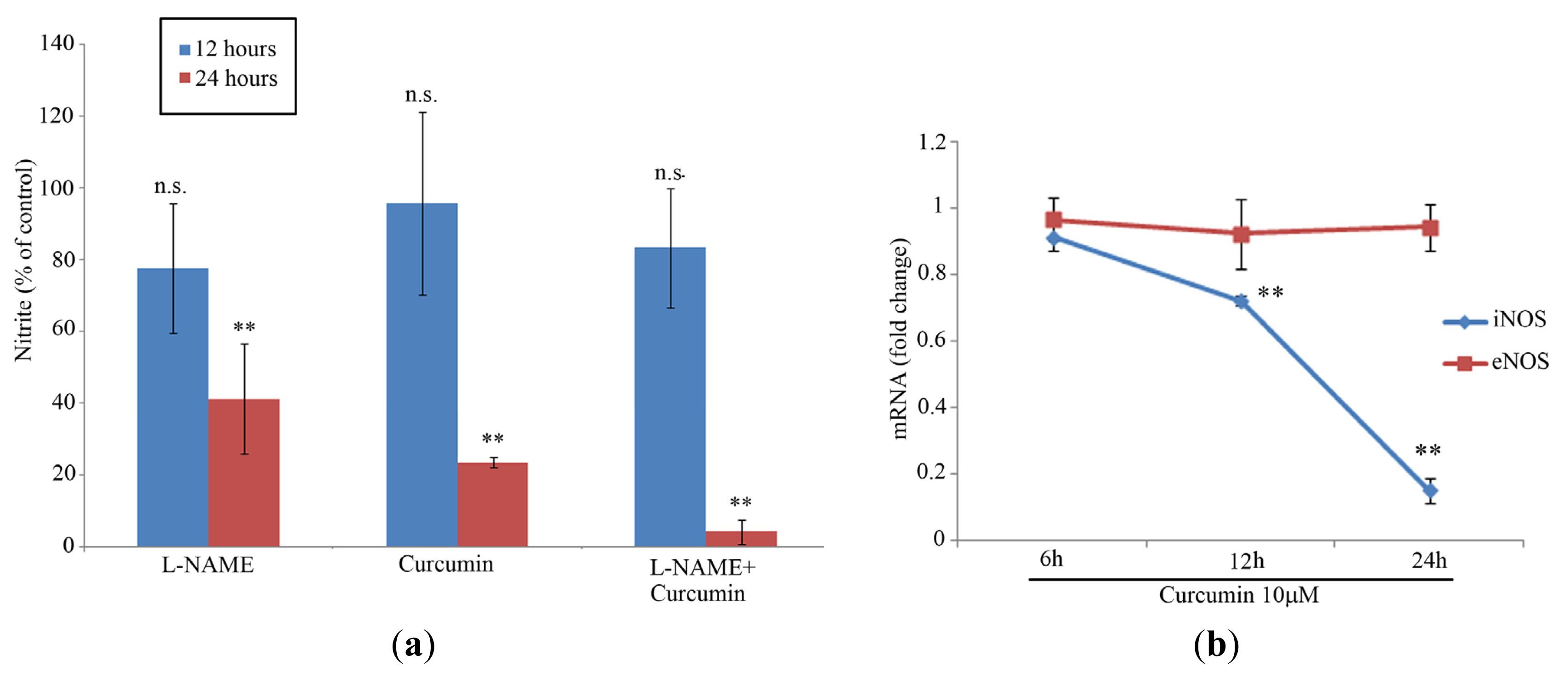
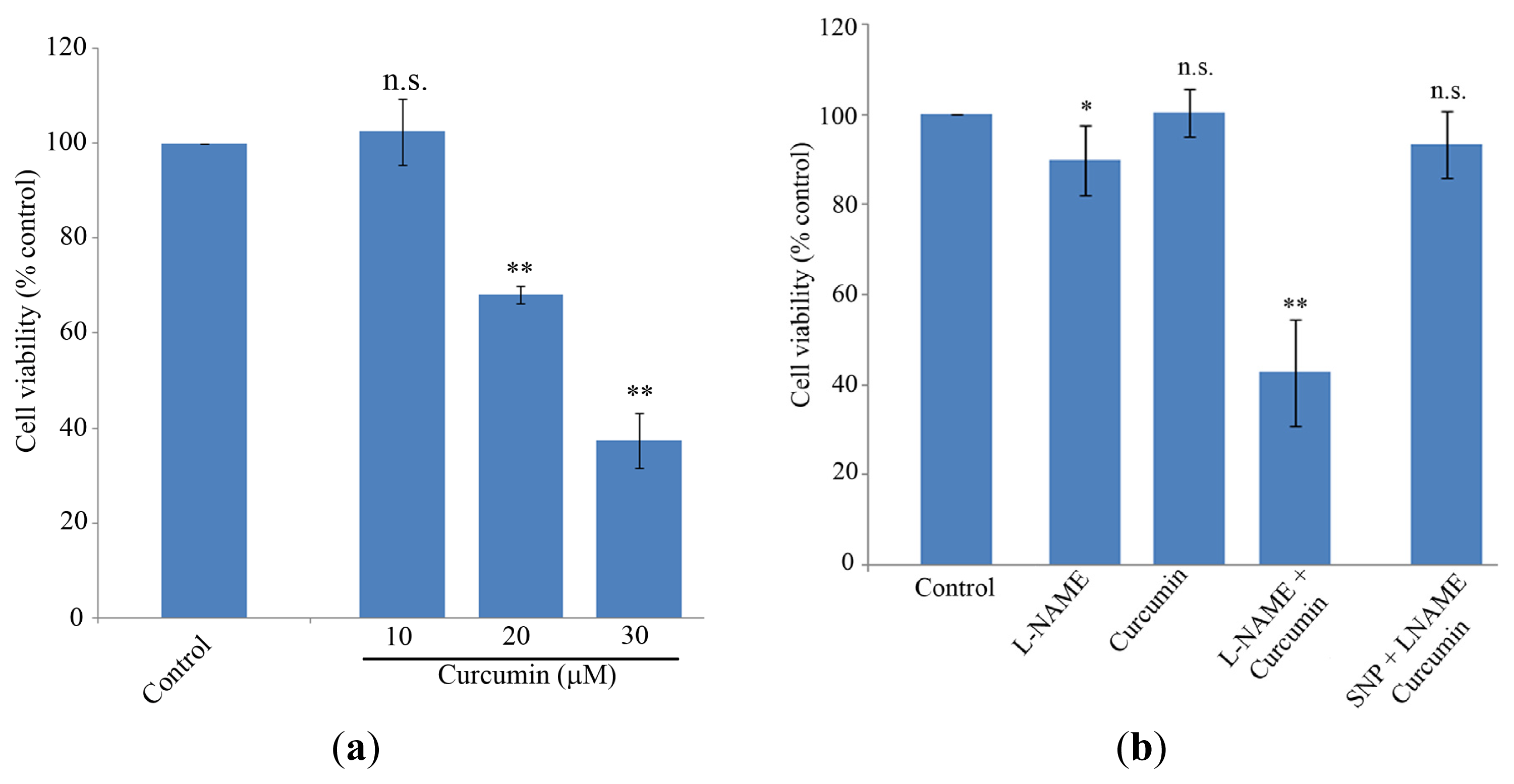
2.1. Cell Proliferation and NO Levels
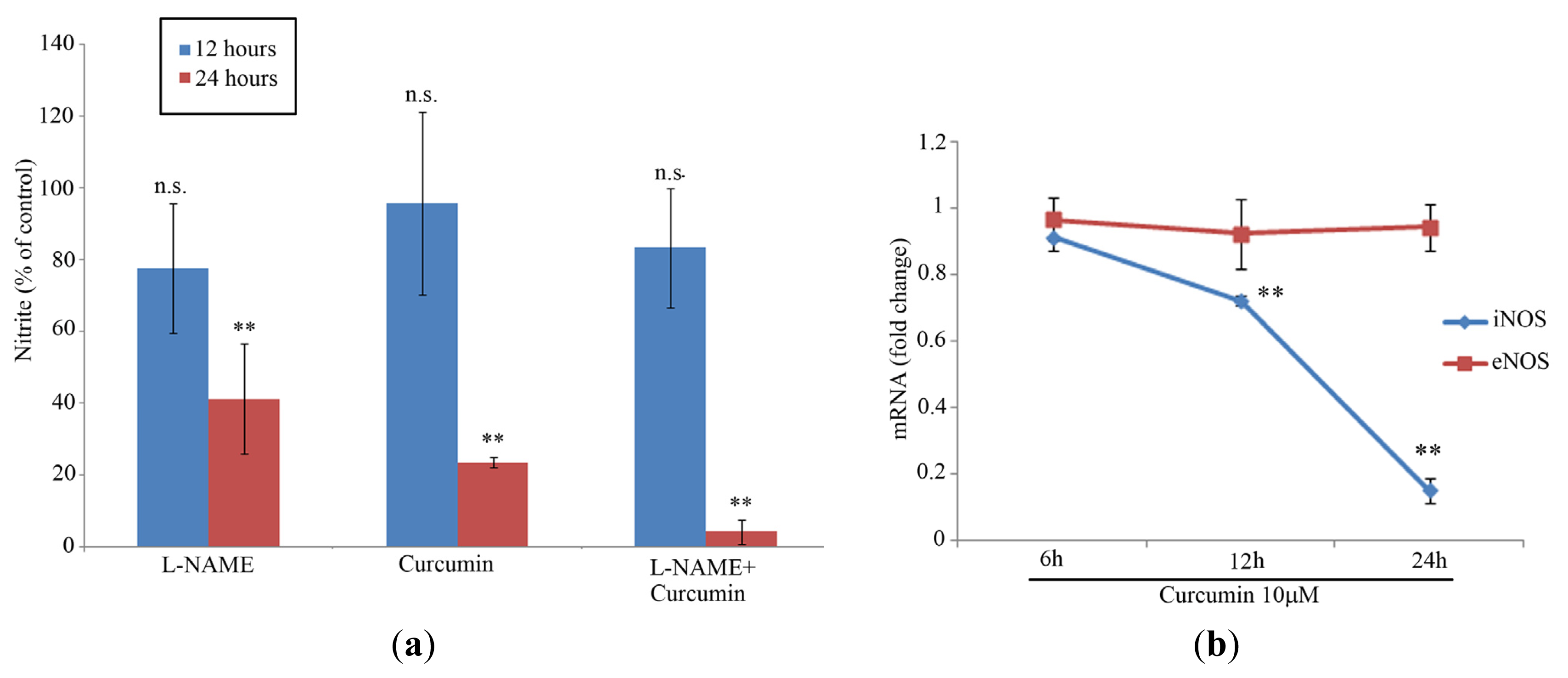
2.2. iNOS/eNOS Gene Expression
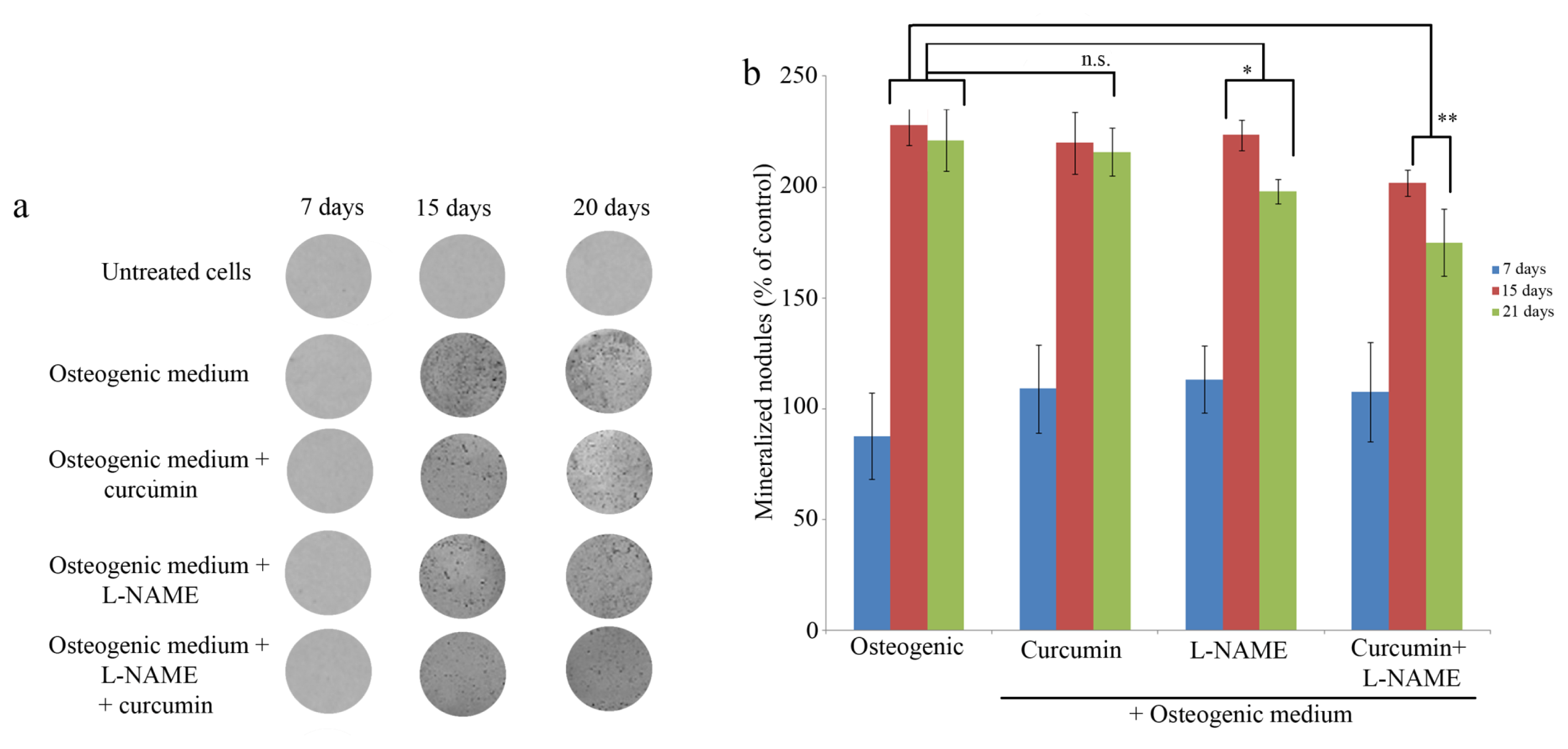
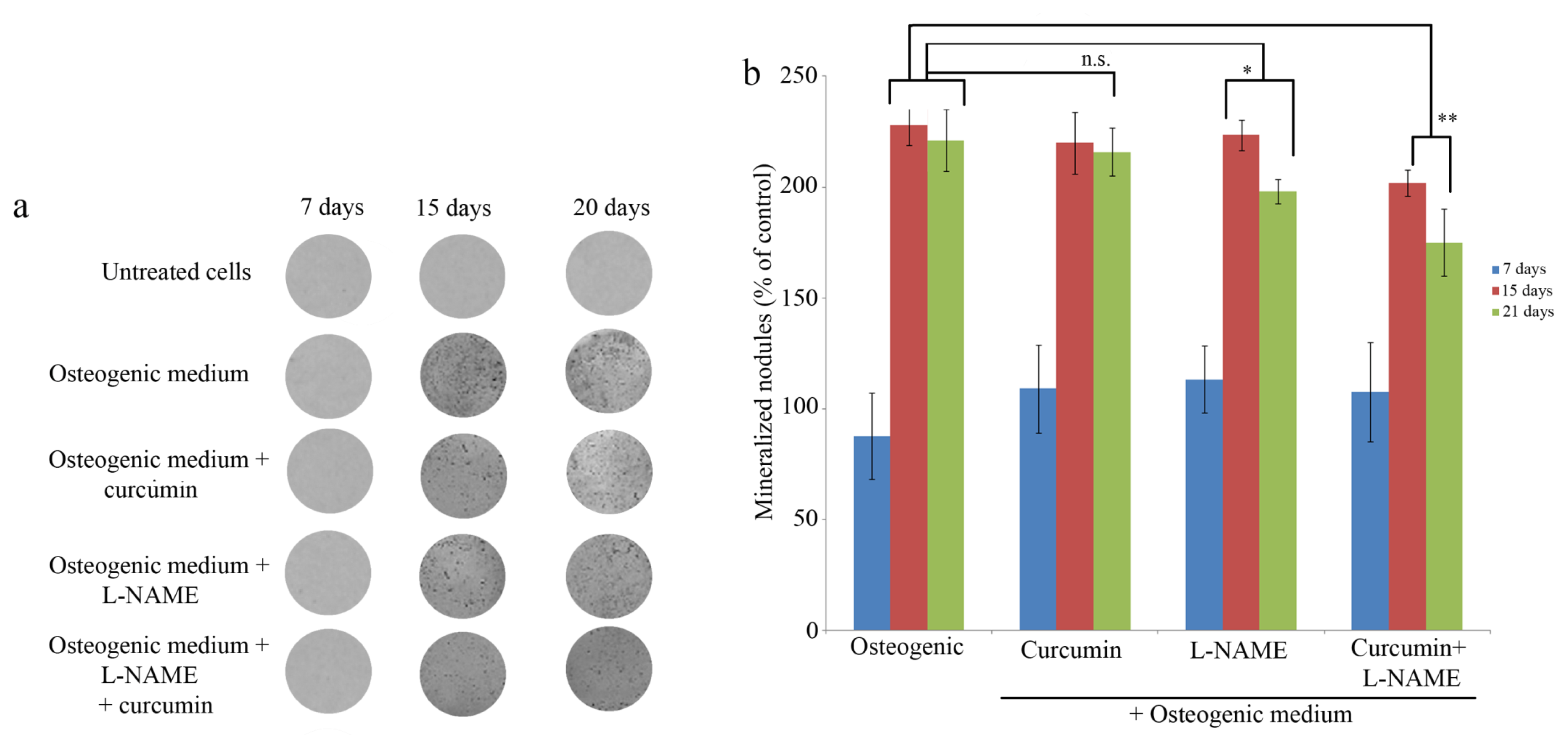
2.3. Mineralization
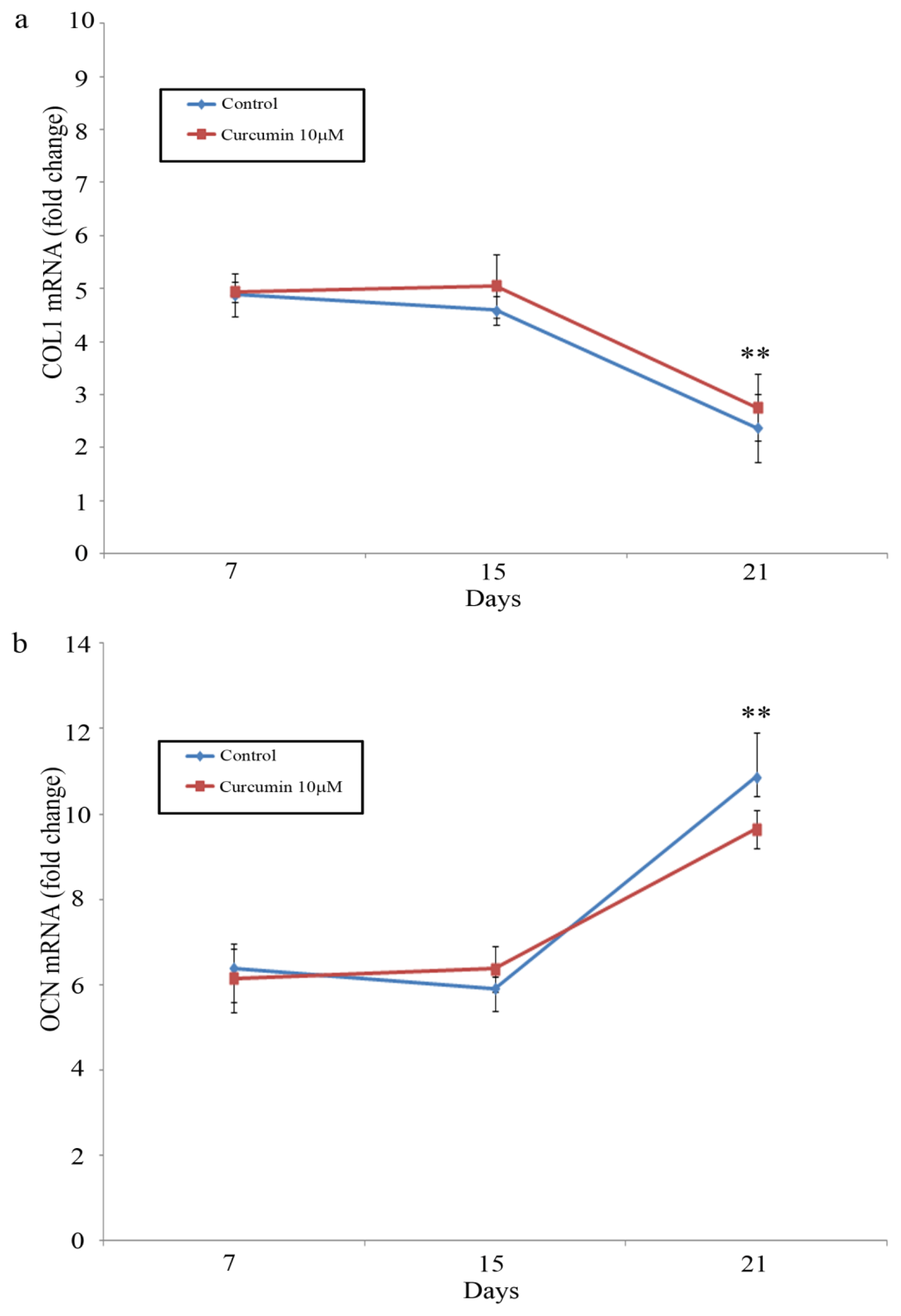
2.4. COLI/OCN Gene Expression
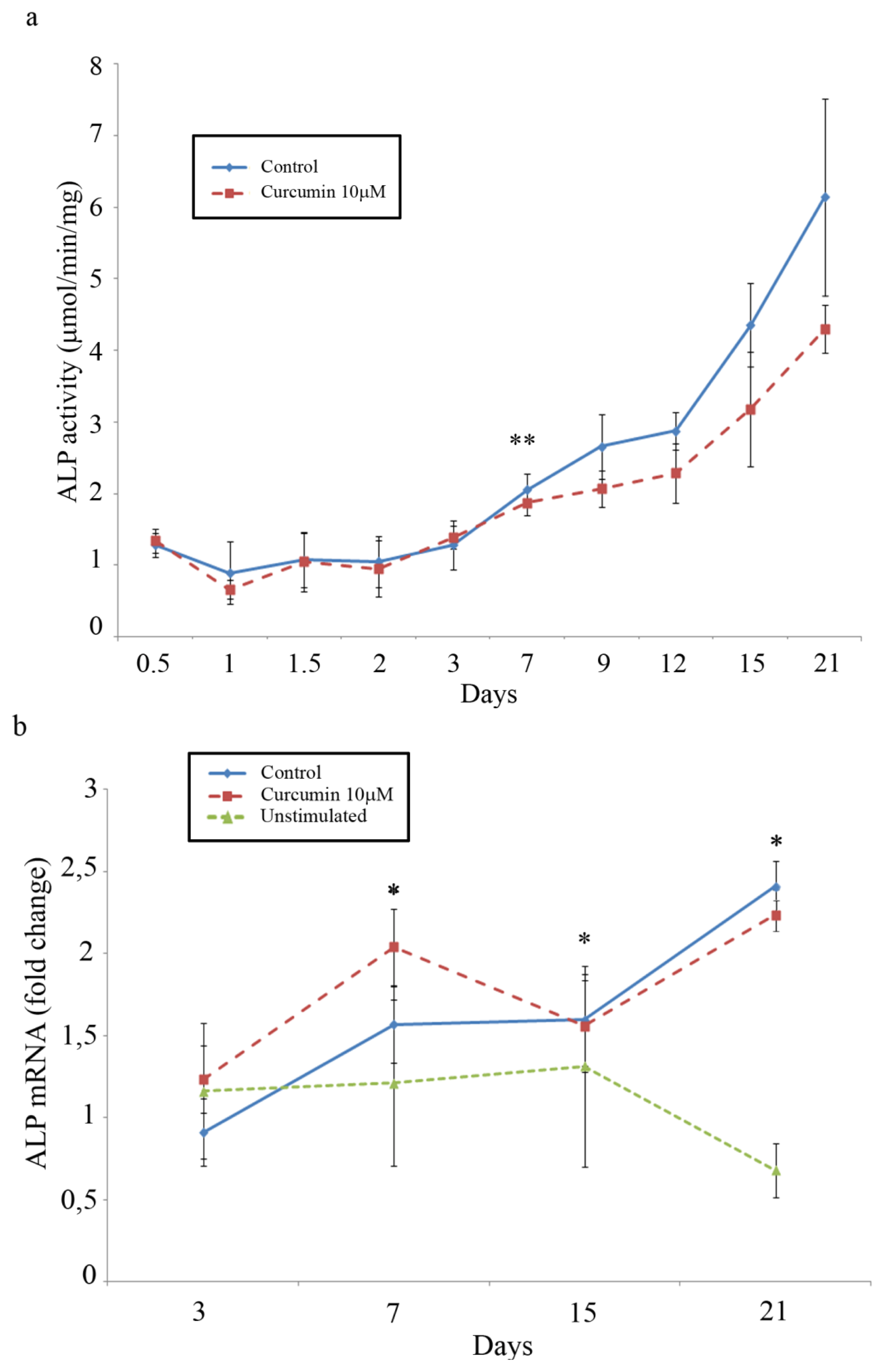
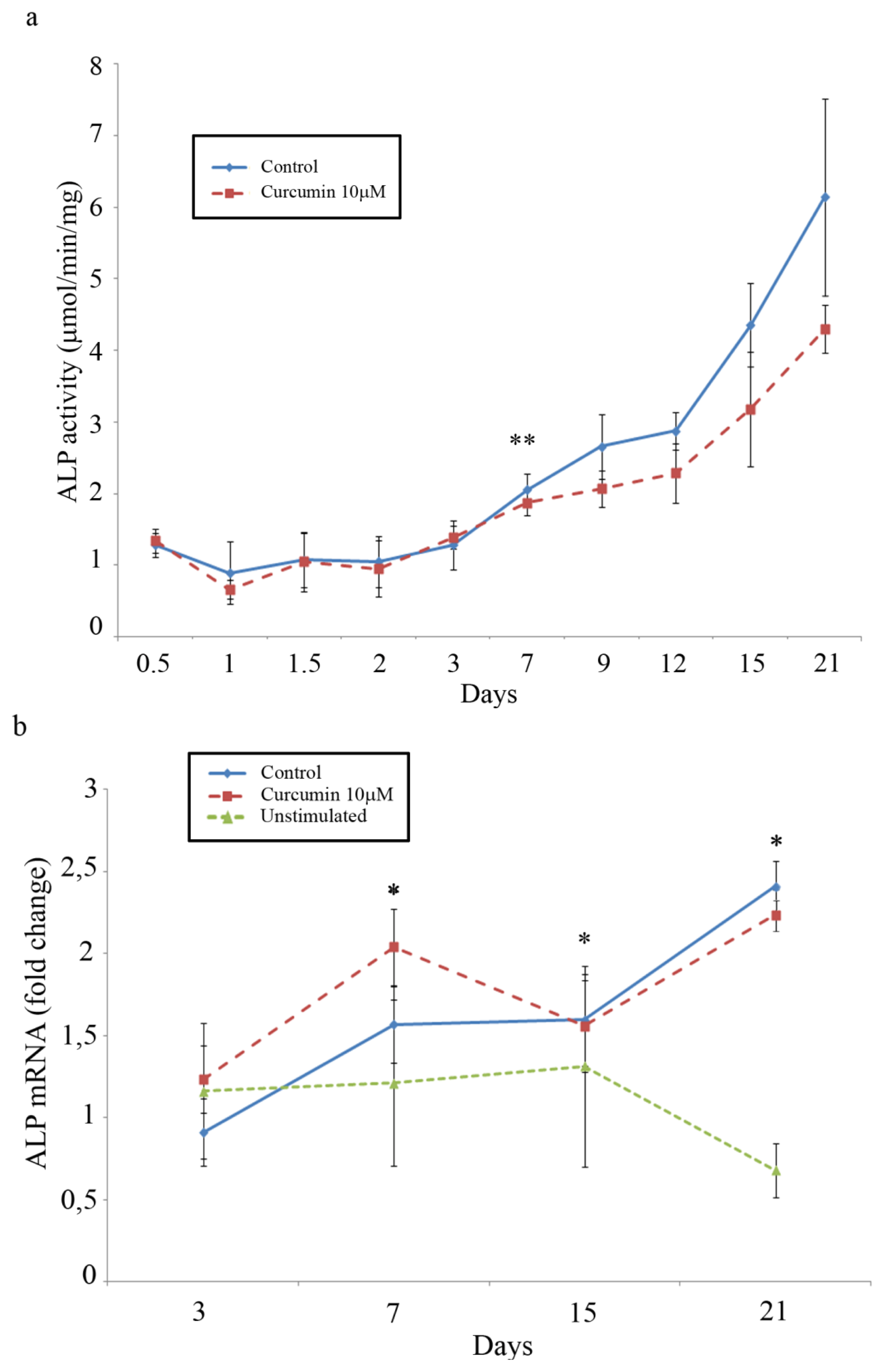
2.5. ALP Activity and Gene Expression
2.6. Discussion
3. Experimental Section
3.1. Tissue Culture
3.2. Cell Proliferation
3.3. Determination of Nitrite/Nitrate (NOx) Concentrations
3.4. RNA Purification
3.5. Quantitative Reverse Transcription-Polymerase Chain Reaction (RT-PCR) for iNOS, eNOS, Type I Collagen (COLI), Alkaline Phosphatase, Osteocalcin (OCN) and β-actin Genes
3.6. Alkaline Phosphatase (ALP) Activity
3.7. Mineralization
3.8. Data Analysis
4. Conclusions
- Conflict of InterestThe authors declare no conflict of interest.
References
- Aggarwal, B.B.; Kumar, A.; Bharti, A.C. Anticancer potential of curcumin: Preclinical and clinical studies. Anticancer Res 2003, 23, 363–398. [Google Scholar]
- Singh, S. From exotic spice to modern drug? Cell 2007, 130, 765–768. [Google Scholar]
- Bar-Sela, G.; Schaffer, M. An Evidence-Based Perspective of Curcuma longa (Turmeric) for Cancer Patients; Springer, Netherlands: Dordrecht, The Netherlands, 2011; Volume Chapter 11, p. 225. [Google Scholar]
- Walters, D.K.; Muff, R.; Langsam, B.; Born, W.; Fuchs, B. Cytotoxic effects of curcumin on osteosarcoma cell lines. Invest. New Drugs 2008, 26, 289–297. [Google Scholar]
- Biswas, S.; Rahman, I. Modulation of steroid activity in chronic inflammation: A novel anti-inflammatory role for curcumin. Mol. Nutr. Food Res 2008, 52, 987–994. [Google Scholar]
- Goel, A.; Jhurani, S.; Aggarwal, B.B. Multi-targeted therapy by curcumin: How spicy is it? Mol. Nutr. Food Res 2008, 52, 1010–1030. [Google Scholar]
- Lopez-Lazaro, M. Anticancer and carcinogenic properties of curcumin: Considerations for its clinical development as a cancer chemopreventive and chemotherapeutic agent. Mol. Nutr. Food Res 2008, 52, S103–S127. [Google Scholar]
- Bharti, A.C.; Takada, Y.; Aggarwal, B.B. Curcumin (diferuloylmethane) inhibits receptor activator of NF-κB ligand-induced NF-κB activation in osteoclast precursors and suppresses osteoclastogenesis. J. Immunol 2004, 172, 5940–5947. [Google Scholar]
- Chan, W.H.; Wu, H.Y.; Chang, W.H. Dosage effects of curcumin on cell death types in a human osteoblast cell line. Food Chem. Toxicol 2006, 44, 1362–1371. [Google Scholar]
- Notoya, M.; Nishimura, H.; Woo, J.T.; Nagai, K.; Ishihara, Y.; Hagiwara, H. Curcumin inhibits the proliferation and mineralization of cultured osteoblasts. Eur. J. Pharmacol 2006, 534, 55–62. [Google Scholar]
- Von, M.I.; Krebbel, H.; Kuckelkorn, U.; Heider, U.; Jakob, C.; Kaiser, M.; Fleissner, C.; Terpos, E.; Sezer, O. Curcumin diminishes human osteoclastogenesis by inhibition of the signalosome-associated I kappaB kinase. J. Cancer Res. Clin. Oncol 2009, 135, 173–179. [Google Scholar]
- Yamaguchi, M.; Hamamoto, R.; Uchiyama, S.; Ishiyama, K. Effects of flavonoid on calcium content in femoral tissue culture and parathyroid hormone-stimulated osteoclastogenesis in bone marrow culture in vitro. Mol. Cell Biochem 2007, 303, 83–88. [Google Scholar]
- Folwarczna, J.; Zych, M.; Trzeciak, H.I. Effects of curcumin on the skeletal system in rats. Pharmacol. Rep 2010, 62, 900–909. [Google Scholar]
- Guimaraes, M.R.; Coimbra, L.S.; de Aquino, S.G.; Spolidorio, L.C.; Kirkwood, K.L.; Rossa, C., Jr. Potent anti-inflammatory effects of systemically administered curcumin modulate periodontal disease in vivo. J. Periodontal Res. 2011, 46, 269–279. [Google Scholar]
- Hussan, F.; Ibraheem, N.G.; Kamarudin, T.A.; Shuid, A.N.; Soelaiman, I.N.; Othman, F. Curcumin protects against ovariectomy-induced bone changes in rat model. Evid. Based Complement. Altern. Med 2012, 2012, 1–7. [Google Scholar]
- Zhou, T.; Chen, D.; Li, Q.; Sun, X.; Song, Y.; Wang, C. Curcumin inhibits inflammatory response and bone loss during experimental periodontitis in rats. Acta Odontol. Scand. 2012. [Google Scholar] [CrossRef]
- Mori, M.; Gotoh, T. Regulation of nitric oxide production by arginine metabolic enzymes. Biochem. Biophys. Res. Commun 2000, 275, 715–719. [Google Scholar]
- Mungrue, I.N.; Bredt, D.S.; Stewart, D.J.; Husain, M. From molecules to mammals: What’s NOS got to do with it? Acta Physiol. Scand 2003, 179, 123–135. [Google Scholar]
- Riancho, J.A.; Salas, E.; Zarrabeitia, M.T.; Olmos, J.M.; Amado, J.A.; Fernandez-Luna, J.L.; Gonzalez-Macias, J. Expression and functional role of nitric oxide synthase in osteoblast-like cells. J. Bone Miner. Res 1995, 10, 439–446. [Google Scholar]
- MacPherson, H.; Noble, B.S.; Ralston, S.H. Expression and functional role of nitric oxide synthase isoforms in human osteoblast-like cells. Bone 1999, 24, 179–185. [Google Scholar]
- Hikiji, H.; Shin, W.S.; Oida, S.; Takato, T.; Koizumi, T.; Toyo-oka, T. Direct action of nitric oxide on osteoblastic differentiation. FEBS Lett 1997, 410, 238–242. [Google Scholar]
- Koyama, A.; Otsuka, E.; Inoue, A.; Hirose, S.; Hagiwara, H. Nitric oxide accelerates the ascorbic acid-induced osteoblastic differentiation of mouse stromal ST2 cells by stimulating the production of prostaglandin E(2). Eur. J. Pharmacol 2000, 391, 225–231. [Google Scholar]
- Saura, M.; Tarin, C.; Zaragoza, C. Recent insights into the implication of nitric oxide in osteoblast differentiation and proliferation during bone development. Sci. World J 2010, 10, 624–632. [Google Scholar]
- Ortiz-Ortiz, M.A.; Moran, J.M.; Bravosanpedro, J.M.; Gonzalez-Polo, R.A.; Niso-Santano, M.; Anantharam, V.; Kanthasamy, A.G.; Soler, G.; Fuentes, J.M. Curcumin enhances paraquat-induced apoptosis of N27 mesencephalic cells via the generation of reactive oxygen species. Neurotoxicology 2009, 30, 1008–1018. [Google Scholar]
- Pan, M.H.; Lin-Shiau, S.Y.; Lin, J.K. Comparative studies on the suppression of nitric oxide synthase by curcumin and its hydrogenated metabolites through down-regulation of IkappaB kinase and NFκB activation in macrophages. Biochem. Pharmacol 2000, 60, 1665–1676. [Google Scholar]
- Chakravarti, N.; Myers, J.N.; Aggarwal, B.B. Targeting constitutive and interleukin-6-inducible signal transducers and activators of transcription 3 pathway in head and neck squamous cell carcinoma cells by curcumin (diferuloylmethane). Int. J. Cancer 2006, 119, 1268–1275. [Google Scholar]
- Leow, P.C.; Tian, Q.; Ong, Z.Y.; Yang, Z.; Ee, P.L. Antitumor activity of natural compounds, curcumin and PKF118-310, as Wnt/beta-catenin antagonists against human osteosarcoma cells. Invest. New Drugs 2010, 28, 766–782. [Google Scholar]
- Epstein, J.; Sanderson, I.R.; Macdonald, T.T. Curcumin as a therapeutic agent: The evidence from in vitro, animal and human studies. Br. J. Nutr 2010, 103, 1545–1557. [Google Scholar]
- Milacic, V.; Banerjee, S.; Landis-Piwowar, K.R.; Sarkar, F.H.; Majumdar, A.P.; Dou, Q.P. Curcumin inhibits the proteasome activity in human colon cancer cells in vitro and in vivo. Cancer Res 2008, 68, 7283–7292. [Google Scholar]
- Reuter, S.; Eifes, S; Dicato, M.; Aggarwal, B.B.; Diederich, M. Modulation of anti-apoptotic and survival pathways by curcumin as a strategy to induce apoptosis in cancer cells. Biochem. Pharmacol. 2008, 76, 1340–1351. [Google Scholar]
- Chen, H.W.; Huang, H.C. Effect of curcumin on cell cycle progression and apoptosis in vascular smooth muscle cells. Br. J. Pharmacol 1998, 124, 1029–1040. [Google Scholar]
- Chen, H.T.; Tsou, H.K.; Chang, C.H.; Tang, C.H. Hepatocyte growth factor increases osteopontin expression in human osteoblasts through PI3K, Akt, c-Src, and AP-1 signaling pathway. PLoS One 2012, 7. [Google Scholar] [CrossRef]
- Yan, L.; Yinghui, T.; Bo, Y.; Gang, Z.; Xian, X.; Lu, Z. Effect of calcitonin gene-related peptide on nitric oxide production in osteoblasts: An experimental study. Cell Biol. Int 2011, 35, 757–765. [Google Scholar]
- Chen, R.M.; Chen, T.L.; Chiu, W.T.; Chang, C.C. Molecular mechanism of nitric oxide-induced osteoblast apoptosis. J. Orthop. Res 2005, 23, 462–468. [Google Scholar]
- Son, M.J.; Lee, S.B.; Byun, Y.J.; Lee, H.O.; Kim, H.S.; Kwon, O.J.; Jeong, S.W. Sodium nitroprusside induces autophagic cell death in glutathione-depleted osteoblasts. J. Biochem. Mol. Toxicol 2010, 24, 313–322. [Google Scholar]
- Wang, J.; Shang, F.; Jiang, R.; Liu, L.; Wang, S.; Hou, J.; Huan, M.; Mei, Q. Nitric oxide-donating genistein prodrug: Design, synthesis, and bioactivity on MC3T3-E1 cells. J. Pharmacol. Sci 2007, 104, 82–89. [Google Scholar]
- Ralston, S.H.; Todd, D.; Helfrich, M.; Benjamin, N.; Grabowski, P.S. Human osteoblast-like cells produce nitric oxide and express inducible nitric oxide synthase. Endocrinology 1994, 135, 330–336. [Google Scholar]
- Riancho, J.A.; Salas, E.; Zarrabeitia, M.T.; Olmos, J.M.; Amado, J.A.; Fernández-Luna, J.L.; González-Macías, J. Expression and functional role of nitric oxide synthase in osteoblast-like cells. J. Bone Miner. Res 1995, 10, 439–446. [Google Scholar]
- Cheng, G.; Zhai, Y.; Chen, K.; Zhou, J.; Han, G.; Zhu, R.; Ming, L.; Song, P.; Wang, J. Sinusoidal electromagnetic field stimulates rat osteoblast differentiation and maturation via activation of NO-cGMP-PKG pathway. Nitric Oxide 2011, 25, 316–325. [Google Scholar]
- Otsuka, E.; Hirano, K.; Matsushita, S.; Inoue, A.; Hirose, S.; Yamaguchi, A.; Hagiwara, H. Effects of nitric oxide from exogenous nitric oxide donors on osteoblastic metabolism. Eur. J. Pharmacol 1998, 349, 345–350. [Google Scholar]
- Bonthius, D.J.; Bonthius, N.E.; Li, S.; Karacay, B. The protective effect of neuronal nitric oxide synthase (nNOS) against alcohol toxicity depends upon the NO-cGMP-PKG pathway and NF-kappaB. Neurotoxicology 2008, 29, 1080–1091. [Google Scholar]
- Yamaguchi, M.; Moore, T.W.; Sun, A.; Snyder, J.P.; Shoji, M. Novel curcumin analogue UBS109 potently stimulates osteoblastogenesis and suppresses osteoclastogenesis: Involvement in Smad activation and NF-kappaB inhibition. Integr. Biol. (Camb.) 2012, 4, 905–913. [Google Scholar]

© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Moran, J.M.; Roncero-Martin, R.; Rodriguez-Velasco, F.J.; Calderon-Garcia, J.F.; Rey-Sanchez, P.; Vera, V.; Canal-Macias, M.L.; Pedrera-Zamorano, J.D. Effects of Curcumin on the Proliferation and Mineralization of Human Osteoblast-Like Cells: Implications of Nitric Oxide. Int. J. Mol. Sci. 2012, 13, 16104-16118. https://doi.org/10.3390/ijms131216104
Moran JM, Roncero-Martin R, Rodriguez-Velasco FJ, Calderon-Garcia JF, Rey-Sanchez P, Vera V, Canal-Macias ML, Pedrera-Zamorano JD. Effects of Curcumin on the Proliferation and Mineralization of Human Osteoblast-Like Cells: Implications of Nitric Oxide. International Journal of Molecular Sciences. 2012; 13(12):16104-16118. https://doi.org/10.3390/ijms131216104
Chicago/Turabian StyleMoran, Jose M., Raul Roncero-Martin, Francisco J. Rodriguez-Velasco, Julian F. Calderon-Garcia, Purificacion Rey-Sanchez, Vicente Vera, Maria L. Canal-Macias, and Juan D. Pedrera-Zamorano. 2012. "Effects of Curcumin on the Proliferation and Mineralization of Human Osteoblast-Like Cells: Implications of Nitric Oxide" International Journal of Molecular Sciences 13, no. 12: 16104-16118. https://doi.org/10.3390/ijms131216104