Nitric Oxide-Dependent Posttranslational Modification in Plants: An Update

Abstract
:1. Introduction
2. Metal Nitrosylation in Plants
3. Tyrosine Nitration in Plants
4. S-Nitrosylation in Plants
5. Conclusions
References
- Martinez-Ruiz, A.; Cadenas, S.; Lamas, S. Nitric oxide signaling: Classical, less classical, and nonclassical mechanisms. Free Radic. Biol. Med 2011, 51, 17–29. [Google Scholar]
- Besson-Bard, A.; Pugin, A.; Wendehenne, D. New insights into nitric oxide signaling in plants. Annu. Rev. Plant Biol 2008, 59, 21–39. [Google Scholar]
- Gupta, K.J.; Fernie, A.R.; Kaiser, W.M.; van Dongen, J.T. On the origins of nitric oxide. Trends Plant Sci 2010, 16, 160–168. [Google Scholar]
- Moreau, M.; Lindermayr, C.; Durner, J.; Klessig, D.F. NO synthesis and signaling in plants— where do we stand? Physiol. Plant 2010, 138, 372–383. [Google Scholar]
- Frohlich, A.; Durner, J. The hunt for plant nitric oxide synthase (NOS): Is one really needed? Plant Sci 2011, 181, 401–404. [Google Scholar]
- Besson-Bard, A.; Courtois, C.; Gauthier, A.; Dahan, J.; Dobrowolska, G.; Jeandroz, S.; Pugin, A.; Wendehenne, D. Nitric oxide in plants: Production and cross-talk with Ca2+ signaling. Mol. Plant 2008, 1, 218–228. [Google Scholar]
- Gaupels, F.; Kuruthukulangarakoola, G.T.; Durner, J. Upstream and downstream signals of nitric oxide in pathogen defence. Curr. Opin. Plant Biol 2011, 14, 707–714. [Google Scholar]
- Wilson, I.D.; Neill, S.J.; Hancock, J.T. Nitric oxide synthesis and signalling in plants. Plant Cell Environ 2008, 31, 622–631. [Google Scholar]
- Ahlfors, R.; Brosche, M.; Kollist, H.; Kangasjarvi, J. Nitric oxide modulates ozone-induced cell death, hormone biosynthesis and gene expression in Arabidopsis thaliana. Plant J 2009, 58, 1–12. [Google Scholar]
- Besson-Bard, A.; Gravot, A.; Richaud, P.; Auroy, P.; Duc, C.; Gaymard, F.; Taconnat, L.; Renou, J.P.; Pugin, A.; Wendehenne, D. Nitric oxide contributes to cadmium toxicity in Arabidopsis by promoting cadmium accumulation in roots and by up-regulating genes related to iron uptake. Plant Physiol 2009, 149, 1302–1315. [Google Scholar]
- Grun, S.; Lindermayr, C.; Sell, S.; Durner, J. Nitric oxide and gene regulation in plants. J. Exp. Bot 2006, 57, 507–516. [Google Scholar]
- Baudouin, E. The language of nitric oxide signalling. Plant Biol. (Stuttg) 2011, 13, 233–242. [Google Scholar]
- Leitner, M.; Vandelle, E.; Gaupels, F.; Bellin, D.; Delledonne, M. NO signals in the haze: Nitric oxide signalling in plant defence. Curr. Opin. Plant Biol 2009, 12, 451–458. [Google Scholar]
- Ma, W.; Berkowitz, G.A. The grateful dead: Calcium and cell death in plant innate immunity. Cell Microbiol 2007, 9, 2571–2585. [Google Scholar]
- Giustarini, D.; Milzani, A.; Aldini, G.; Carini, M.; Rossi, R.; Dalle-Donne, I. S-nitrosation versus S-glutathionylation of protein sulfhydryl groups by S-nitrosoglutathione. Antioxid. Redox Signal 2005, 7, 930–939. [Google Scholar]
- Gopalakrishna, R.; Chen, Z.H.; Gundimeda, U. Nitric oxide and nitric oxide-generating agents induce a reversible inactivation of protein kinase C activity and phorbol ester binding. J. Biol. Chem 1993, 268, 27180–27185. [Google Scholar]
- Tada, Y.; Spoel, S.H.; Pajerowska-Mukhtar, K.; Mou, Z.; Song, J.; Wang, C.; Zuo, J.; Dong, X. Plant immunity requires conformational changes [corrected] of NPR1 via S-nitrosylation and thioredoxins. Science 2008, 321, 952–956. [Google Scholar]
- Ford, P.C. Reactions of NO and nitrite with heme models and proteins. Inorg. Chem 2010, 49, 6226–6239. [Google Scholar]
- Toledo, J.C., Jr; Augusto, O. Connecting the chemical and biological properties of nitric oxide. Chem. Res. Toxicol. 2012, 25, 975–989. [Google Scholar]
- Seth, D.; Stamler, J.S. The SNO-proteome: Causation and classifications. Curr. Opin. Chem. Biol 2010, 15, 129–136. [Google Scholar]
- Bellota-Anton, C.; Munnoch, J.; Robb, K.; Adamczyk, K.; Candelaresi, M.; Parker, A.W.; Dixon, R.; Hutchings, M.I.; Hunt, N.T.; Tucker, N.P. Spectroscopic analysis of protein Fe-NO complexes. Biochem. Soc. Trans 2011, 39, 1293–1298. [Google Scholar]
- Lewandowska, H.; Kalinowska, M.; Brzoska, K.; Wojciuk, K.; Wojciuk, G.; Kruszewski, M. Nitrosyl iron complexes—synthesis, structure and biology. Dalton Trans 2011, 40, 8273–8289. [Google Scholar]
- Stadler, J.; Bergonia, H.A.; di Silvio, M.; Sweetland, M.A.; Billiar, T.R.; Simmons, R.L.; Lancaster, J.R., Jr. Nonheme iron-nitrosyl complex formation in rat hepatocytes: Detection by electron paramagnetic resonance spectroscopy. Arch. Biochem. Biophys. 1993, 302, 4–11. [Google Scholar]
- Derbyshire, E.R.; Marletta, M.A. Structure and regulation of soluble guanylate cyclase. Annu. Rev. Biochem 2012, 81, 533–559. [Google Scholar]
- Ahern, G.P.; Klyachko, V.A.; Jackson, M.B. cGMP and S-nitrosylation: Two routes for modulation of neuronal excitability by NO. Trends Neurosci 2002, 25, 510–517. [Google Scholar]
- Durner, J.; Wendehenne, D.; Klessig, D.F. Defense gene induction in tobacco by nitric oxide, cyclic GMP, and cyclic ADP-ribose. Proc. Natl Acad. Sci. USA 1998, 95, 10328–10333. [Google Scholar]
- Ma, W.; Smigel, A.; Verma, R.; Berkowitz, G.A. Cyclic nucleotide gated channels and related signaling components in plant innate immunity. Plant Signal. Behav 2009, 4, 277–282. [Google Scholar]
- Mulaudzi, T.; Ludidi, N.; Ruzvidzo, O.; Morse, M.; Hendricks, N.; Iwuoha, E.; Gehring, C. Identification of a novel Arabidopsis thaliana nitric oxide-binding molecule with guanylate cyclase activity in vitro. FEBS Lett 2011, 585, 2693–2697. [Google Scholar]
- Gupta, K.J.; Hebelstrup, K.H.; Mur, L.A.; Igamberdiev, A.U. Plant hemoglobins: Important players at the crossroads between oxygen and nitric oxide. FEBS Lett 2011, 585, 3843–3849. [Google Scholar]
- Hill, R.D. Non-symbiotic haemoglobins-What's happening beyond nitric oxide scavenging? AoB Plants 2012, 2012, pls004. [Google Scholar]
- Igamberdiev, A.U.; Bykova, N.V.; Hill, R.D. Structural and functional properties of class 1 plant hemoglobins. IUBMB Life 2011, 63, 146–152. [Google Scholar]
- Igamberdiev, A.U.; Hill, R.D. Nitrate, NO and haemoglobin in plant adaptation to hypoxia: An alternative to classic fermentation pathways. J. Exp. Bot 2004, 55, 2473–2482. [Google Scholar]
- Perazzolli, M.; Dominici, P.; Romero-Puertas, M.C.; Zago, E.; Zeier, J.; Sonoda, M.; Lamb, C.; Delledonne, M. Arabidopsis nonsymbiotic hemoglobin AHb1 modulates nitric oxide bioactivity. Plant Cell 2004, 16, 2785–2794. [Google Scholar]
- Seregelyes, C.; Igamberdiev, A.U.; Maassen, A.; Hennig, J.; Dudits, D.; Hill, R.D. NO-degradation by alfalfa class 1 hemoglobin (Mhb1): A possible link to PR-1a gene expression in Mhb1-overproducing tobacco plants. FEBS Lett 2004, 571, 61–66. [Google Scholar]
- Perazzolli, M.; Romero-Puertas, M.C.; Delledonne, M. Modulation of nitric oxide bioactivity by plant haemoglobins. J. Exp. Bot 2006, 57, 479–488. [Google Scholar]
- Herold, S.; Puppo, A. Oxyleghemoglobin scavenges nitrogen monoxide and peroxynitrite: A possible role in functioning nodules? J. Biol. Inorg. Chem 2005, 10, 935–945. [Google Scholar]
- Mathieu, C.; Moreau, S.; Frendo, P.; Puppo, A.; Davies, M.J. Direct detection of radicals in intact soybean nodules: Presence of nitric oxide-leghemoglobin complexes. Free Radic. Biol. Med 1998, 24, 1242–1249. [Google Scholar]
- Sanchez, C.; Cabrera, J.J.; Gates, A.J.; Bedmar, E.J.; Richardson, D.J.; Delgado, M.J. Nitric oxide detoxification in the rhizobia-legume symbiosis. Biochem. Soc. Trans 2011, 39, 184–188. [Google Scholar]
- Sasakura, F.; Uchiumi, T.; Shimoda, Y.; Suzuki, A.; Takenouchi, K.; Higashi, S.; Abe, M. A class 1 hemoglobin gene from Alnus firma functions in symbiotic and nonsymbiotic tissues to detoxify nitric oxide. Mol. Plant Microbe Interact 2006, 19, 441–450. [Google Scholar]
- Sturms, R.; DiSpirito, A.A.; Hargrove, M.S. Plant and cyanobacterial hemoglobins reduce nitrite to nitric oxide under anoxic conditions. Biochemistry 2011, 50, 3873–3878. [Google Scholar]
- Tiso, M.; Tejero, J.; Kenney, C.; Frizzell, S.; Gladwin, M.T. Nitrite Reductase Activity of Nonsymbiotic Hemoglobins from Arabidopsis thaliana. Biochemistry 2012, 51, 5285–5292. [Google Scholar]
- Brown, G.C. Regulation of mitochondrial respiration by nitric oxide inhibition of cytochrome c oxidase. Biochim. Biophys. Acta 2001, 1504, 46–57. [Google Scholar]
- Clark, D.; Durner, J.; Navarre, D.A.; Klessig, D.F. Nitric oxide inhibition of tobacco catalase and ascorbate peroxidase. Mol. Plant Microb. Interact 2000, 13, 1380–1384. [Google Scholar]
- Millar, A.H.; Day, D.A. Nitric oxide inhibits the cytochrome oxidase but not the alternative oxidase of plant mitochondria. FEBS Lett 1996, 398, 155–158. [Google Scholar]
- Nelson, M.J. The nitric oxide complex of ferrous soybean lipoxygenase-1. Substrate, pH, and ethanol effects on the active-site iron. J. Biol. Chem 1987, 262, 12137–12142. [Google Scholar]
- Gupta, K.J.; Shah, J.K.; Brotman, Y.; Jahnke, K.; Willmitzer, L.; Kaiser, W.M.; Bauwe, H.; Igamberdiev, A.U. Inhibition of aconitase by nitric oxide leads to induction of the alternative oxidase and to a shift of metabolism towards biosynthesis of amino acids. J. Exp. Bot 2012, 63, 1773–1784. [Google Scholar]
- Schopfer, F.J.; Baker, P.R.; Freeman, B.A. NO-dependent protein nitration: A cell signaling event or an oxidative inflammatory response? Trends Biochem. Sci 2003, 28, 646–654. [Google Scholar]
- Gunther, M.R.; Sturgeon, B.E.; Mason, R.P. Nitric oxide trapping of the tyrosyl radical-chemistry and biochemistry. Toxicology 2002, 177, 1–9. [Google Scholar]
- Bayden, A.S.; Yakovlev, V.A.; Graves, P.R.; Mikkelsen, R.B.; Kellogg, G.E. Factors influencing protein tyrosine nitration—structure-based predictive models. Free Radic. Biol. Med 2011, 50, 749–762. [Google Scholar]
- Ischiropoulos, H. Biological selectivity and functional aspects of protein tyrosine nitration. Biochem. Biophys. Res. Commun 2003, 305, 776–783. [Google Scholar]
- Abello, N.; Kerstjens, H.A.; Postma, D.S.; Bischoff, R. Protein tyrosine nitration: Selectivity, physicochemical and biological consequences, denitration, and proteomics methods for the identification of tyrosine-nitrated proteins. J. Proteome Res 2009, 8, 3222–3238. [Google Scholar]
- Vandelle, E.; Delledonne, M. Peroxynitrite formation and function in plants. Plant Sci 2011, 181, 534–539. [Google Scholar]
- Cecconi, D.; Orzetti, S.; Vandelle, E.; Rinalducci, S.; Zolla, L.; Delledonne, M. Protein nitration during defense response in Arabidopsis thaliana. Electrophoresis 2009, 30, 2460–2468. [Google Scholar]
- Romero-Puertas, M.C.; Laxa, M.; Matte, A.; Zaninotto, F.; Finkemeier, I.; Jones, A.M.; Perazzolli, M.; Vandelle, E.; Dietz, K.J.; Delledonne, M. S-nitrosylation of peroxiredoxin II E promotes peroxynitrite-mediated tyrosine nitration. Plant Cell 2007, 19, 4120–4130. [Google Scholar]
- Wisastra, R.; Poelstra, K.; Bischoff, R.; Maarsingh, H.; Haisma, H.J.; Dekker, F.J. Antibody-free detection of protein tyrosine nitration in tissue sections. Chembiochem 2011, 12, 2016–2020. [Google Scholar]
- Berton, P.; Dominguez-Romero, J.C.; Wuilloud, R.G.; Sanchez-Calvo, B.; Chaki, M.; Carreras, A.; Valderrama, R.; Begara-Morales, J.C.; Corpas, F.J.; Barroso, J.B.; et al. Determination of nitrotyrosine in Arabidopsis thaliana cell cultures with a mixed-mode solid-phase extraction cleanup followed by liquid chromatography time-of-flight mass spectrometry. Anal. Bioanal. Chem 2012, 404, 1495–1503. [Google Scholar]
- Chaki, M.; Fernandez-Ocana, A.M.; Valderrama, R.; Carreras, A.; Esteban, F.J.; Luque, F.; Gomez-Rodriguez, M.V.; Begara-Morales, J.C.; Corpas, F.J.; Barroso, J.B. Involvement of reactive nitrogen and oxygen species (RNS and ROS) in sunflower-mildew interaction. Plant Cell Physiol 2009, 50, 265–279. [Google Scholar]
- Tanou, G.; Filippou, P.; Belghazi, M.; Job, D.; Diamantidis, G.; Fotopoulos, V.; Molassiotis, A. Oxidative and nitrosative-based signaling and associated post-translational modifications orchestrate the acclimation of citrus plants to salinity stress. Plant J 2012, 72, 585–595. [Google Scholar]
- Sharov, V.S.; Dremina, E.S.; Galeva, N.A.; Gerstenecker, G.S.; Li, X.; Dobrowsky, R.T.; Stobaugh, J.F.; Schoneich, C. Fluorogenic Tagging of Peptide and Protein 3-Nitrotyrosine with 4-(Aminomethyl)-benzenesulfonic Acid for Quantitative Analysis of Protein Tyrosine Nitration. Chromatographia 2010, 71, 37–53. [Google Scholar]
- Morot-Gaudry-Talarmain, Y.; Rockel, P.; Moureaux, T.; Quillere, I.; Leydecker, M.T.; Kaiser, W.M.; Morot-Gaudry, J.F. Nitrite accumulation and nitric oxide emission in relation to cellular signaling in nitrite reductase antisense tobacco. Planta 2002, 215, 708–715. [Google Scholar]
- Saito, S.; Yamamoto-Katou, A.; Yoshioka, H.; Doke, N.; Kawakita, K. Peroxynitrite generation and tyrosine nitration in defense responses in tobacco BY-2 cells. Plant Cell Physiol 2006, 47, 689–697. [Google Scholar]
- Valderrama, R.; Corpas, F.J.; Carreras, A.; Fernandez-Ocana, A.; Chaki, M.; Luque, F.; Gomez-Rodriguez, M.V.; Colmenero-Varea, P.; del Rio, L.A.; Barroso, J.B. Nitrosative stress in plants. FEBS Lett 2007, 581, 453–461. [Google Scholar]
- Galetskiy, D.; Lohscheider, J.N.; Kononikhin, A.S.; Popov, I.A.; Nikolaev, E.N.; Adamska, I. Phosphorylation and nitration levels of photosynthetic proteins are conversely regulated by light stress. Plant Mol. Biol 2011, 77, 461–473. [Google Scholar]
- Galetskiy, D.; Lohscheider, J.N.; Kononikhin, A.S.; Popov, I.A.; Nikolaev, E.N.; Adamska, I. Mass spectrometric characterization of photooxidative protein modifications in Arabidopsis thaliana thylakoid membranes. Rapid Commun. Mass Spectrom 2011, 25, 184–190. [Google Scholar]
- Chaki, M.; Valderrama, R.; Fernandez-Ocana, A.M.; Carreras, A.; Lopez-Jaramillo, J.; Luque, F.; Palma, J.M.; Pedrajas, J.R.; Begara-Morales, J.C.; Sanchez-Calvo, B.; et al. Protein targets of tyrosine nitration in sunflower (Helianthus annuus L.) hypocotyls. J. Exp. Bot 2009, 60, 4221–4234. [Google Scholar]
- Lozano-Juste, J.; Colom-Moreno, R.; Leon, J. In vivo protein tyrosine nitration in Arabidopsis thaliana. J. Exp. Bot 2011, 62, 3501–3517. [Google Scholar]
- Chaki, M.; Valderrama, R.; Fernandez-Ocana, A.M.; Carreras, A.; Gomez-Rodriguez, M.V.; Lopez-Jaramillo, J.; Begara-Morales, J.C.; Sanchez-Calvo, B.; Luque, F.; et al. High temperature triggers the metabolism of S-nitrosothiols in sunflower mediating a process of nitrosative stress which provokes the inhibition of ferredoxin-NADP reductase by tyrosine nitration. Plant Cell Environ 2011, 34, 1803–1818. [Google Scholar]
- Alvarez, C.; Lozano-Juste, J.; Romero, L.C.; Garcia, I.; Gotor, C.; Leon, J. Inhibition of Arabidopsis O-acetylserine(thiol)lyase A1 by tyrosine nitration. J. Biol. Chem 2011, 286, 578–586. [Google Scholar]
- Melo, P.M.; Silva, L.S.; Ribeiro, I.; Seabra, A.R.; Carvalho, H.G. Glutamine synthetase is a molecular target of nitric oxide in root nodules of Medicago truncatula and is regulated by tyrosine nitration. Plant Physiol 2011, 157, 1505–1517. [Google Scholar]
- Astier, J.; Rasul, S.; Koen, E.; Manzoor, H.; Besson-Bard, A.; Lamotte, O.; Jeandroz, S.; Durner, J.; Lindermayr, C.; Wendehenne, D. S-nitrosylation: An emerging post-translational protein modification in plants. Plant Sci 2011, 181, 527–533. [Google Scholar]
- Lindermayr, C.; Durner, J. S-Nitrosylation in plants: Pattern and function. J. Proteomics 2009, 73, 1–9. [Google Scholar]
- Wang, Y.; Yun, B.W.; Kwon, E.; Hong, J.K.; Yoon, J.; Loake, G.J. S-nitrosylation: An emerging redox-based post-translational modification in plants. J. Exp. Bot 2006, 57, 1777–1784. [Google Scholar]
- Feechan, A.; Kwon, E.; Yun, B.W.; Wang, Y.; Pallas, J.A.; Loake, G.J. A central role for S-nitrosothiols in plant disease resistance. Proc. Natl. Acad. Sci. USA 2005, 102, 8054–8059. [Google Scholar]
- Rusterucci, C.; Espunya, M.C.; Diaz, M.; Chabannes, M.; Martinez, M.C. S-nitrosoglutathione reductase affords protection against pathogens in Arabidopsis, both locally and systemically. Plant Physiol 2007, 143, 1282–1292. [Google Scholar]
- Yu, M.; Yun, B.W.; Spoel, S.H.; Loake, G.J. A sleigh ride through the SNO: Regulation of plant immune function by protein S-nitrosylation. Curr. Opin. Plant Biol 2012, 15, 424–430. [Google Scholar]
- Hess, D.T.; Matsumoto, A.; Kim, S.O.; Marshall, H.E.; Stamler, J.S. Protein S-nitrosylation: Purview and parameters. Nat. Rev. Mol. Cell Biol 2005, 6, 150–166. [Google Scholar]
- Marino, S.M.; Gladyshev, V.N. Structural analysis of cysteine S-nitrosylation: A modified acid-based motif and the emerging role of trans-nitrosylation. J. Mol. Biol 2010, 395, 844–859. [Google Scholar]
- Benhar, M.; Forrester, M.T.; Hess, D.T.; Stamler, J.S. Regulated protein denitrosylation by cytosolic and mitochondrial thioredoxins. Science 2008, 320, 1050–1054. [Google Scholar]
- Benhar, M.; Forrester, M.T.; Stamler, J.S. Protein denitrosylation: Enzymatic mechanisms and cellular functions. Nat. Rev. Mol. Cell. Biol 2009, 10, 721–732. [Google Scholar]
- Jaffrey, S.R.; Snyder, S.H. The biotin switch method for the detection of S-nitrosylated proteins. Sci. STKE 2001, 2001. [Google Scholar] [CrossRef]
- Fares, A.; Rossignol, M.; Peltier, J.B. Proteomics investigation of endogenous S-nitrosylation in Arabidopsis. Biochem. Biophys. Res. Commun 2011, 416, 331–336. [Google Scholar]
- Lindermayr, C.; Saalbach, G.; Durner, J. Proteomic identification of S-nitrosylated proteins in Arabidopsis. Plant Physiol 2005, 137, 921–930. [Google Scholar]
- Romero-Puertas, M.C.; Campostrini, N.; Matte, A.; Righetti, P.G.; Perazzolli, M.; Zolla, L.; Roepstorff, P.; Delledonne, M. Proteomic analysis of S-nitrosylated proteins in Arabidopsis thaliana undergoing hypersensitive response. Proteomics 2008, 8, 1459–1469. [Google Scholar]
- Abat, J.K.; Mattoo, A.K.; Deswal, R. S-nitrosylated proteins of a medicinal CAM plant Kalanchoe pinnata-ribulose-1,5-bisphosphate carboxylase/oxygenase activity targeted for inhibition. FEBS J 2008, 275, 2862–2872. [Google Scholar]
- Abat, J.K.; Deswal, R. Differential modulation of S-nitrosoproteome of Brassica juncea by low temperature: Change in S-nitrosylation of Rubisco is responsible for the inactivation of its carboxylase activity. Proteomics 2009, 9, 4368–4380. [Google Scholar]
- Tanou, G.; Job, C.; Rajjou, L.; Arc, E.; Belghazi, M.; Diamantidis, G.; Molassiotis, A.; Job, D. Proteomics reveals the overlapping roles of hydrogen peroxide and nitric oxide in the acclimation of citrus plants to salinity. Plant J 2009, 60, 795–804. [Google Scholar]
- Lin, A.; Wang, Y.; Tang, J.; Xue, P.; Li, C.; Liu, L.; Hu, B.; Yang, F.; Loake, G.J.; Chu, C. Nitric oxide and protein S-nitrosylation are integral to hydrogen peroxide-induced leaf cell death in rice. Plant Physiol 2012, 158, 451–464. [Google Scholar]
- Astier, J.; Besson-Bard, A.; Lamotte, O.; Bertoldo, J.; Bourque, S.; Terenzi, H.; Wendehenne, D. Nitric oxide inhibits the ATPase activity of the chaperone-like AAA+ATPase CDC48, a target for S-nitrosylation in cryptogein signaling in tobacco cells. Biochem. J. 2012. [Google Scholar] [CrossRef]
- Palmieri, M.C.; Lindermayr, C.; Bauwe, H.; Steinhauser, C.; Durner, J. Regulation of plant glycine decarboxylase by S-nitrosylation and glutathionylation. Plant Physiol 2010, 152, 1514–1528. [Google Scholar]
- Ortega-Galisteo, A.P.; Rodriguez-Serrano, M.; Pazmino, D.M.; Gupta, D.K.; Sandalio, L.M.; Romero-Puertas, M.C. S-Nitrosylated proteins in pea (Pisum sativum L.) leaf peroxisomes: Changes under abiotic stress. J. Exp. Bot 2012, 63, 2089–2103. [Google Scholar]
- Belenghi, B.; Romero-Puertas, M.C.; Vercammen, D.; Brackenier, A.; Inze, D.; Delledonne, M.; van Breusegem, F. Metacaspase activity of Arabidopsis thaliana is regulated by S-nitrosylation of a critical cysteine residue. J. Biol. Chem 2007, 282, 1352–1358. [Google Scholar]
- Holtgrefe, S.; Gohlke, J.; Starmann, J.; Druce, S.; Klocke, S.; Altmann, B.; Wojtera, J.; Lindermayr, C.; Scheibe, R. Regulation of plant cytosolic glyceraldehyde 3-phosphate dehydrogenase isoforms by thiol modifications. Physiol. Plant 2008, 133, 211–228. [Google Scholar]
- Lindermayr, C.; Saalbach, G.; Bahnweg, G.; Durner, J. Differential inhibition of Arabidopsis methionine adenosyltransferases by protein S-nitrosylation. J. Biol. Chem 2006, 281, 4285–4291. [Google Scholar]
- Lindermayr, C.; Sell, S.; Muller, B.; Leister, D.; Durner, J. Redox regulation of the NPR1-TGA1 system of Arabidopsis thaliana by nitric oxide. Plant Cell 2010, 22, 2894–2907. [Google Scholar]
- Serpa, V.; Vernal, J.; Lamattina, L.; Grotewold, E.; Cassia, R.; Terenzi, H. Inhibition of AtMYB2 DNA-binding by nitric oxide involves cysteine S-nitrosylation. Biochem. Biophys. Res. Commun 2007, 361, 1048–1053. [Google Scholar]
- Terrile, M.C.; Paris, R.; Calderon-Villalobos, L.I.; Iglesias, M.J.; Lamattina, L.; Estelle, M.; Casalongue, C.A. Nitric oxide influences auxin signaling through S-nitrosylation of the Arabidopsis TRANSPORT INHIBITOR RESPONSE 1 auxin receptor. Plant J 2012, 70, 492–500. [Google Scholar]
- Wang, Y.Q.; Feechan, A.; Yun, B.W.; Shafiei, R.; Hofmann, A.; Taylor, P.; Xue, P.; Yang, F.Q.; Xie, Z.S.; Pallas, J.A.; et al. S-nitrosylation of AtSABP3 antagonizes the expression of plant immunity. J. Biol. Chem 2009, 284, 2131–2137. [Google Scholar]
- Yun, B.W.; Feechan, A.; Yin, M.; Saidi, N.B.; le Bihan, T.; Yu, M.; Moore, J.W.; Kang, J.G.; Kwon, E.; Spoel, S.H.; et al. S-nitrosylation of NADPH oxidase regulates cell death in plant immunity. Nature 2011, 478, 264–268. [Google Scholar]
- Wawer, I.; Bucholc, M.; Astier, J.; Anielska-Mazur, A.; Dahan, J.; Kulik, A.; Wyslouch-Cieszynska, A.; Zareba-Koziol, M.; Krzywinska, E.; Dadlez, M.; et al. Regulation of Nicotiana tabacum osmotic stress-activated protein kinase and its cellular partner GAPDH by nitric oxide in response to salinity. Biochem. J 2010, 429, 73–83. [Google Scholar]
- Astier, J.; Kulik, A.; Koen, E.; Besson-Bard, A.; Bourque, S.; Jeandroz, S.; Lamotte, O.; Wendehenne, D. Protein S-nitrosylation: What’s going on in plants? Free Radic. Biol. Med 2012, 53, 1101–1110. [Google Scholar]
- Dong, X. NPR1, all things considered. Curr. Opin. Plant Biol 2004, 7, 547–552. [Google Scholar]
- Slaymaker, D.H.; Navarre, D.A.; Clark, D.; del Pozo, O.; Martin, G.B.; Klessig, D.F. The tobacco salicylic acid-binding protein 3 (SABP3) is the chloroplast carbonic anhydrase, which exhibits antioxidant activity and plays a role in the hypersensitive defense response. Proc. Natl. Acad. Sci. USA 2002, 99, 11640–11645. [Google Scholar]
- Torres, M.A.; Dangl, J.L.; Jones, J.D. Arabidopsis gp91phox homologues AtrbohD and AtrbohF are required for accumulation of reactive oxygen intermediates in the plant defense response. Proc. Natl. Acad. Sci. USA 2002, 99, 517–522. [Google Scholar]
- Besson-Bard, A.; Griveau, S.; Bedioui, F.; Wendehenne, D. Real-time electrochemical detection of extracellular nitric oxide in tobacco cells exposed to cryptogein, an elicitor of defence responses. J. Exp. Bot 2008, 59, 3407–3414. [Google Scholar]
- Foissner, I.; Wendehenne, D.; Langebartels, C.; Durner, J. In vivo imaging of an elicitor-induced nitric oxide burst in tobacco. Plant J 2000, 23, 817–824. [Google Scholar]
- Lamotte, O.; Gould, K.; Lecourieux, D.; Sequeira-Legrand, A.; Lebrun-Garcia, A.; Durner, J.; Pugin, A.; Wendehenne, D. Analysis of nitric oxide signaling functions in tobacco cells challenged by the elicitor cryptogein. Plant Physiol 2004, 135, 516–529. [Google Scholar]
- Bae, H.; Choi, S.M.; Yang, S.W.; Pai, H.S.; Kim, W.T. Suppression of the ER-localized AAA ATPase NgCDC48 inhibits tobacco growth and development. Mol. Cells 2009, 28, 57–65. [Google Scholar]
- Muller, J.; Piffanelli, P.; Devoto, A.; Miklis, M.; Elliott, C.; Ortmann, B.; Schulze-Lefert, P.; Panstruga, R. Conserved ERAD-like quality control of a plant polytopic membrane protein. Plant Cell 2005, 17, 149–163. [Google Scholar]
- O'Quin, J.B.; Bourassa, L.; Zhang, D.; Shockey, J.M.; Gidda, S.K.; Fosnot, S.; Chapman, K.D.; Mullen, R.T.; Dyer, J.M. Temperature-sensitive post-translational regulation of plant omega-3 fatty-acid desaturases is mediated by the endoplasmic reticulum-associated degradation pathway. J. Biol. Chem 2010, 285, 21781–21796. [Google Scholar]
- Park, S.; Rancour, D.M.; Bednarek, S.Y. In planta analysis of the cell cycle-dependent localization of AtCDC48A and its critical roles in cell division, expansion, and differentiation. Plant Physiol 2008, 148, 246–258. [Google Scholar]
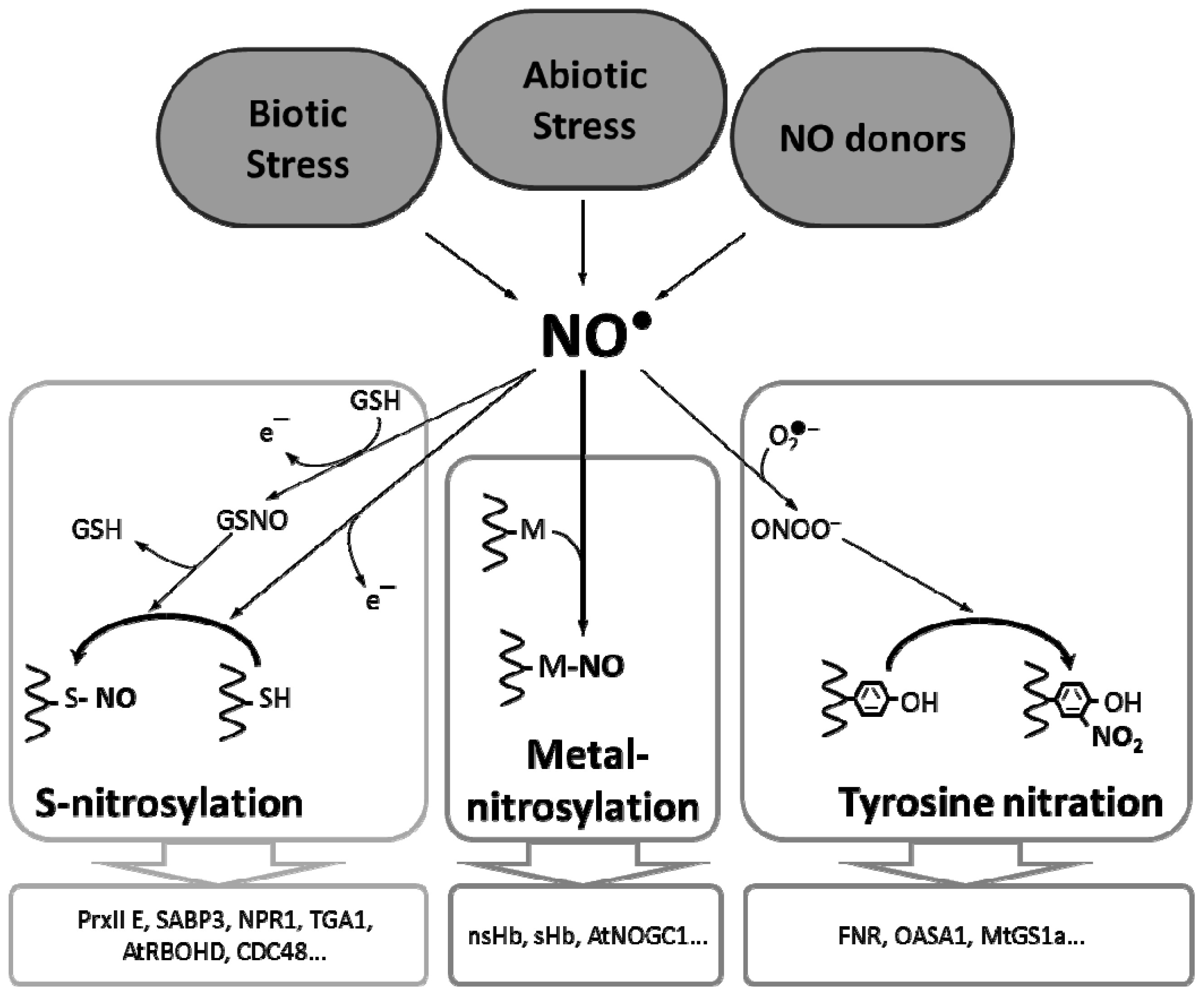

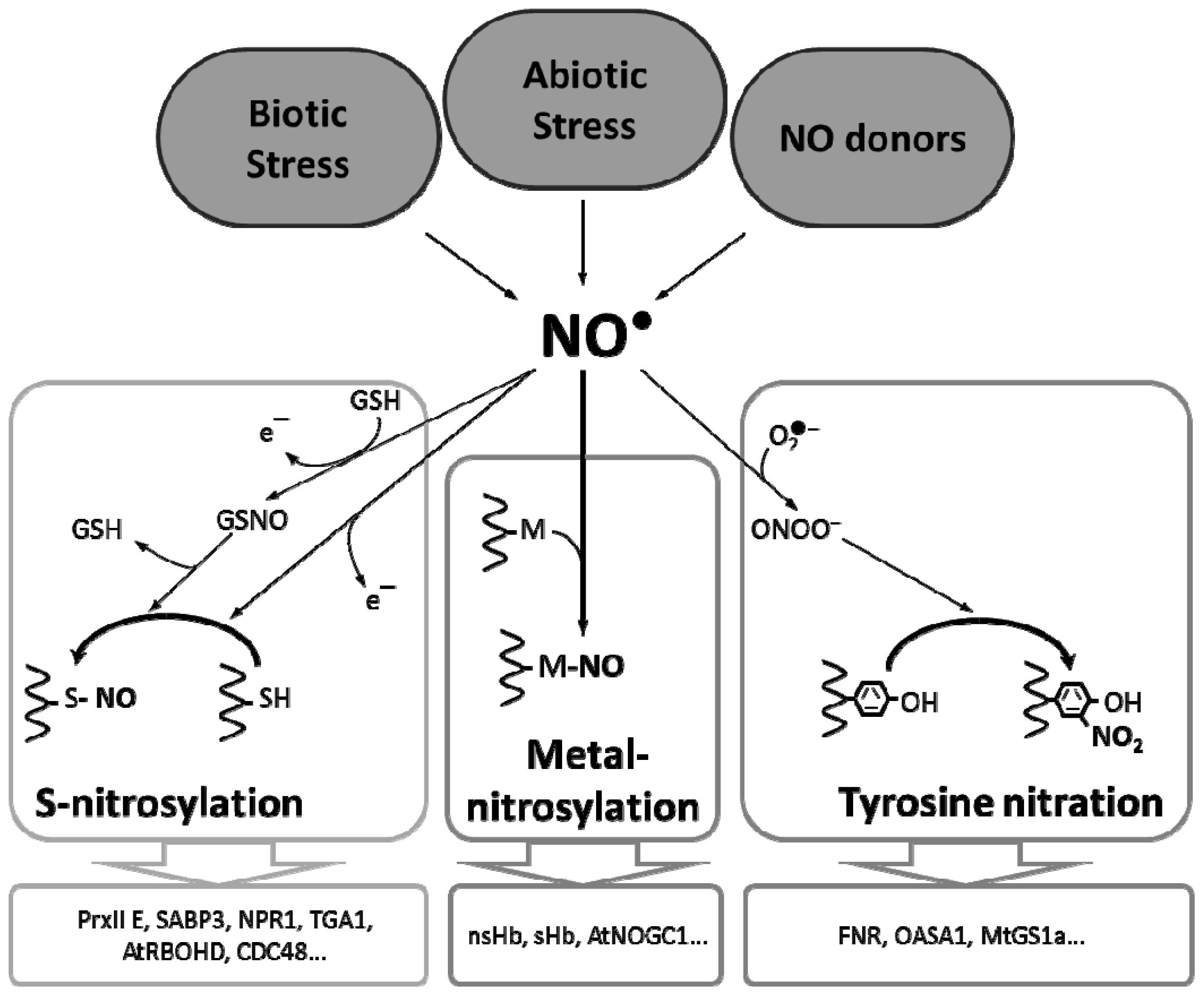{kind=link}
| Posttranslational modification | Target protein | References |
|---|---|---|
| Metal nitrosylation | NO-dependant Gunylate cyclase 1 (AtNOGC1) | [24] |
| Hemoglobins | [25–37] | |
| Aconitase | [42] | |
| Tyrosine nitration | Ferredoxin-NADP oxidoreductase (FNR) | [63] |
| O-acetylserine(thiol)lyase A1 (OASA1) | [64] | |
| Guanylate cyclase | [65] | |
| S-nitrosylation | Peroxiredoxin II E (PrxII E) | [50] |
| Nonexpressor of pathogenesis-related gene 1 (NPR1) | [75,90] | |
| Transcription factor TGA1 | [90] | |
| Salicylic acid binding protein 3 (SABP3) | [93] | |
| Respiratory burst oxidase homologue D (RBOHD) | [94] | |
| Cell division cycle 48 (CDC48) | [84] | |
| Transport inhibitor response 1 (TIR1) | [92] |
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Astier, J.; Lindermayr, C. Nitric Oxide-Dependent Posttranslational Modification in Plants: An Update. Int. J. Mol. Sci. 2012, 13, 15193-15208. https://doi.org/10.3390/ijms131115193
Astier J, Lindermayr C. Nitric Oxide-Dependent Posttranslational Modification in Plants: An Update. International Journal of Molecular Sciences. 2012; 13(11):15193-15208. https://doi.org/10.3390/ijms131115193
Chicago/Turabian StyleAstier, Jeremy, and Christian Lindermayr. 2012. "Nitric Oxide-Dependent Posttranslational Modification in Plants: An Update" International Journal of Molecular Sciences 13, no. 11: 15193-15208. https://doi.org/10.3390/ijms131115193