Apoptosis Induction of Human Prostate Carcinoma DU145 Cells by Diallyl Disulfide via Modulation of JNK and PI3K/AKT Signaling Pathways

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
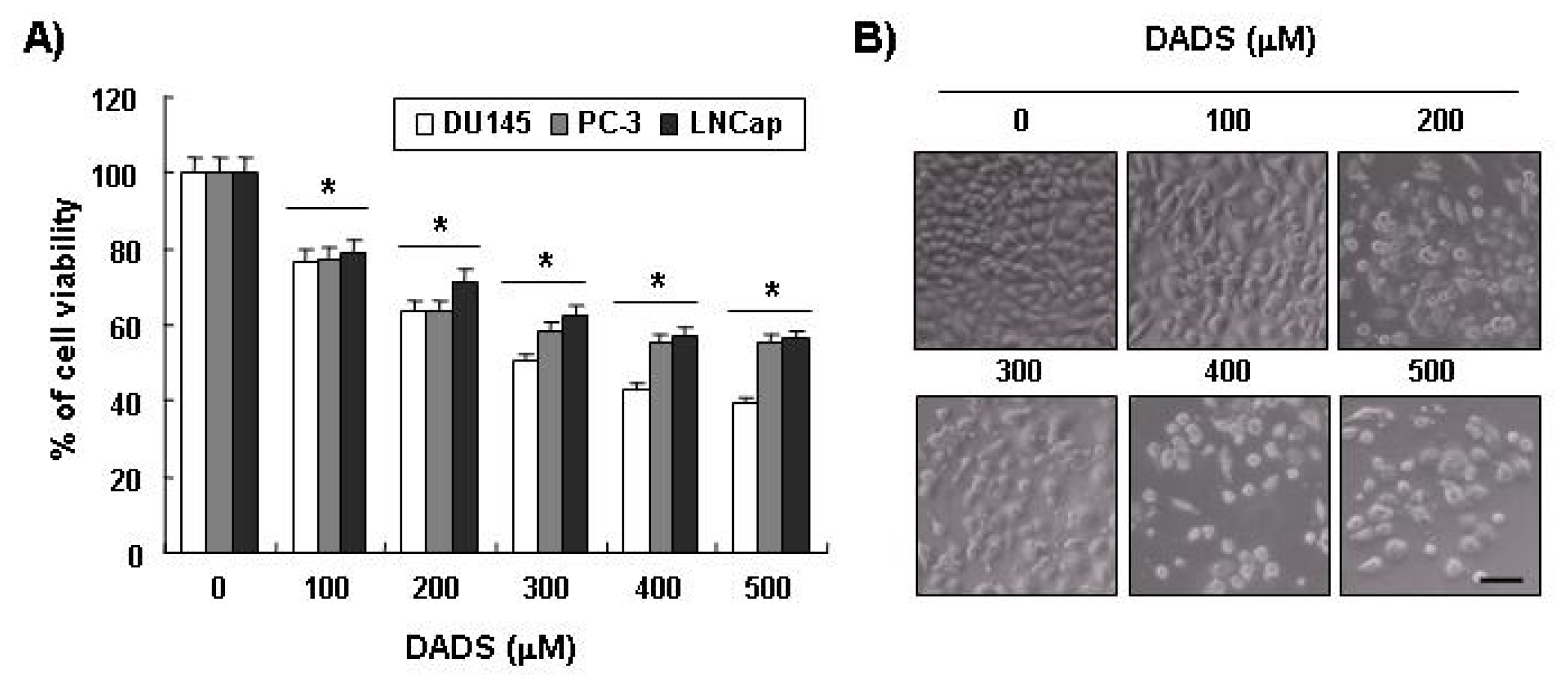
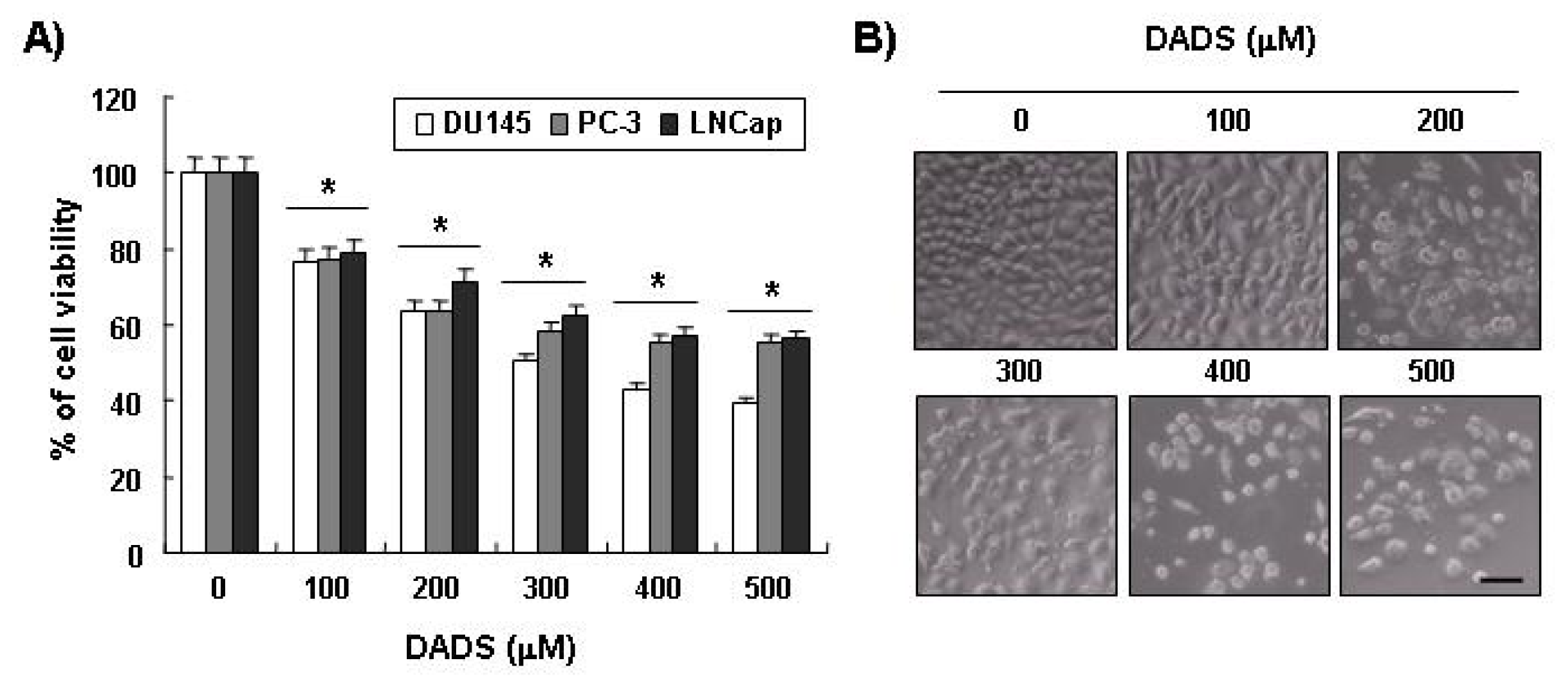
2.1. Inhibition of Cell Viability by DADS in DU145 Cells
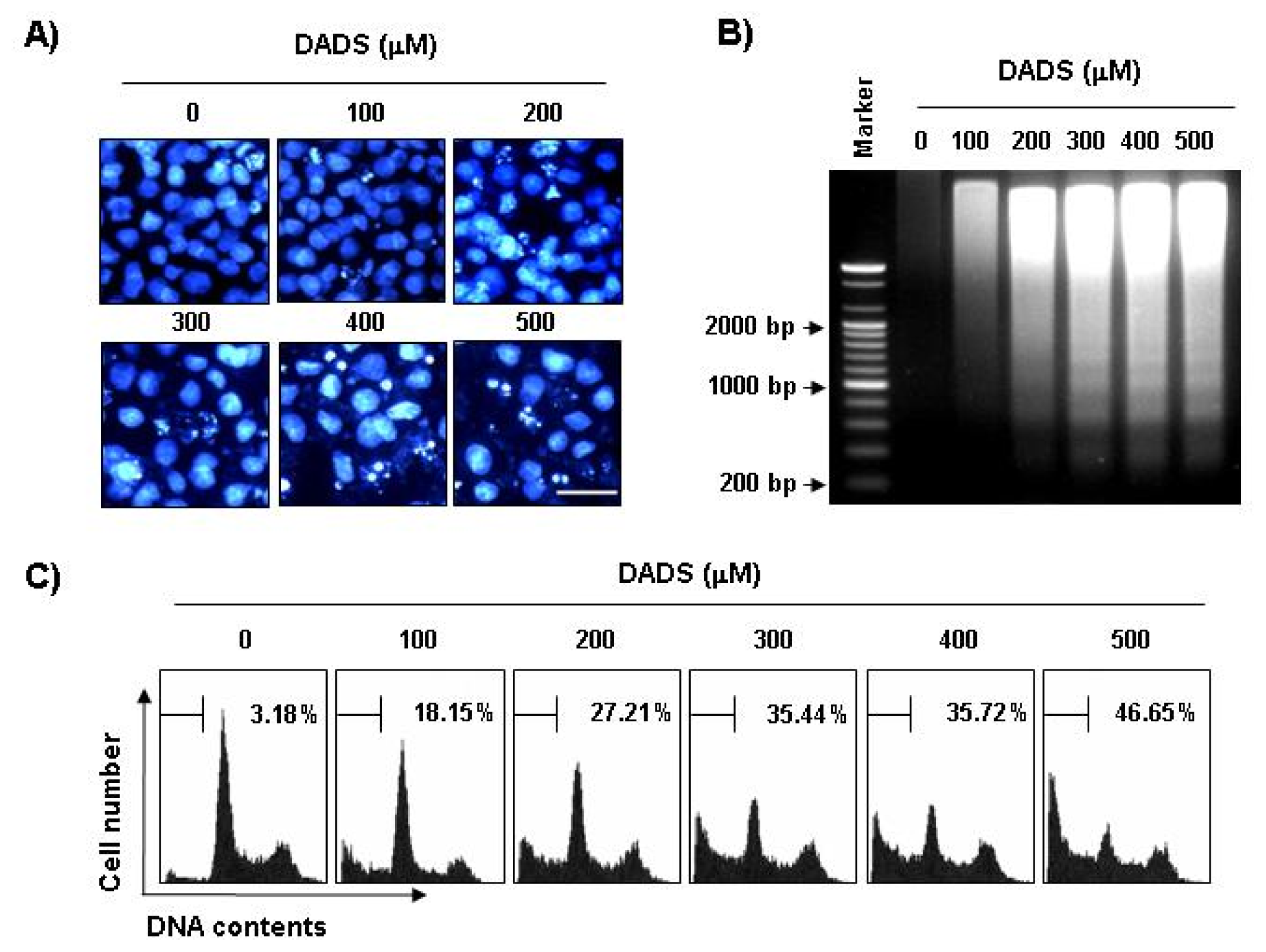
2.2. Induction of Apoptosis by DADS in DU145 Cells
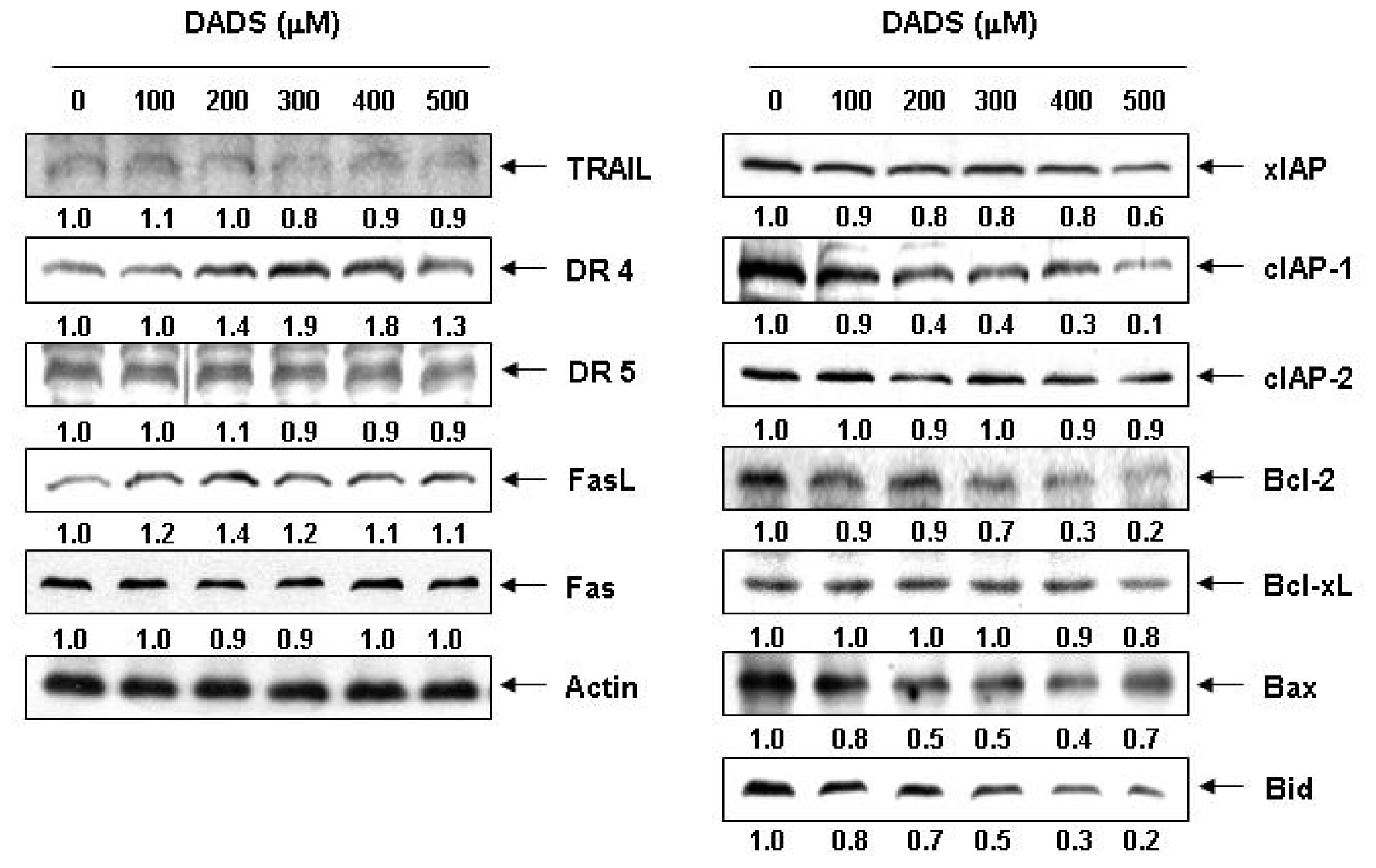
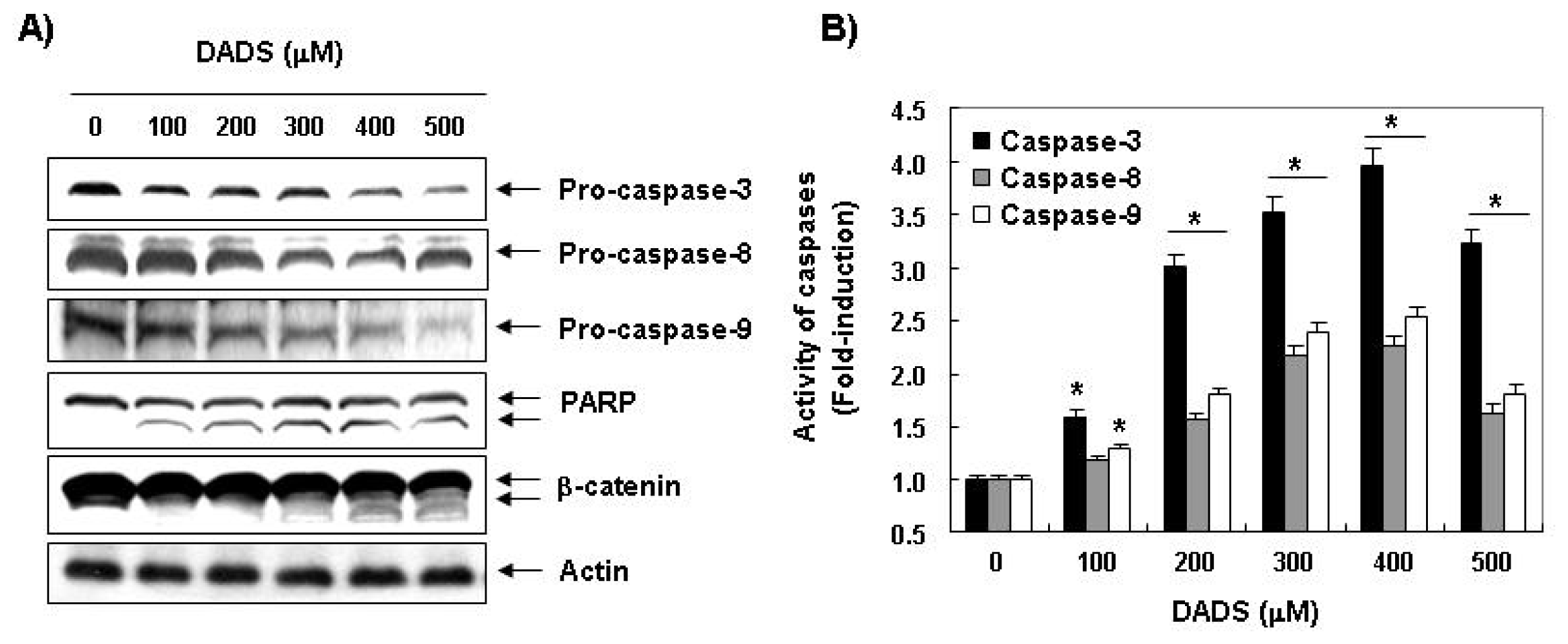
2.3. Effects of DADS on the Expression of Apoptosis-Related Proteins in DU145 Cells
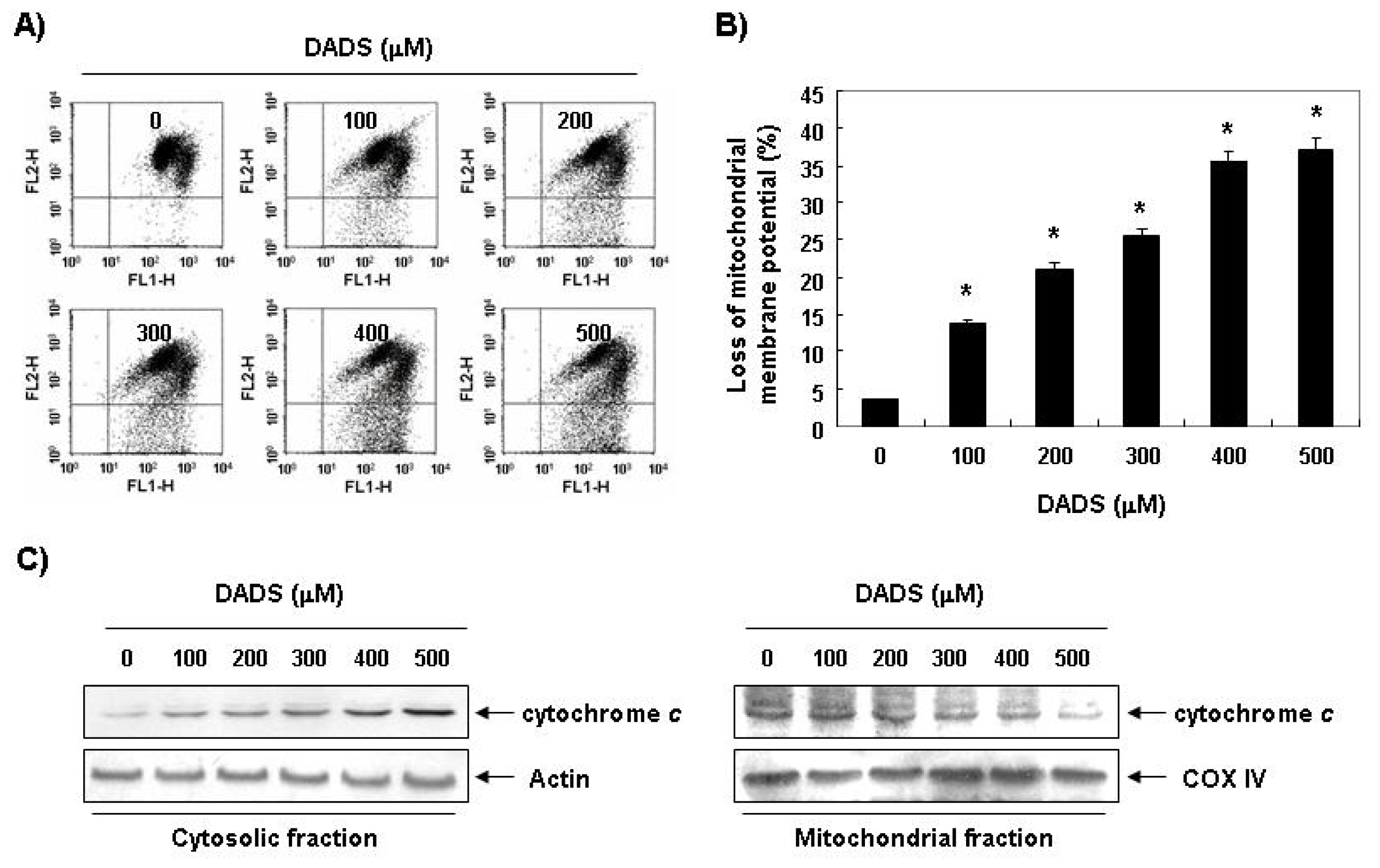
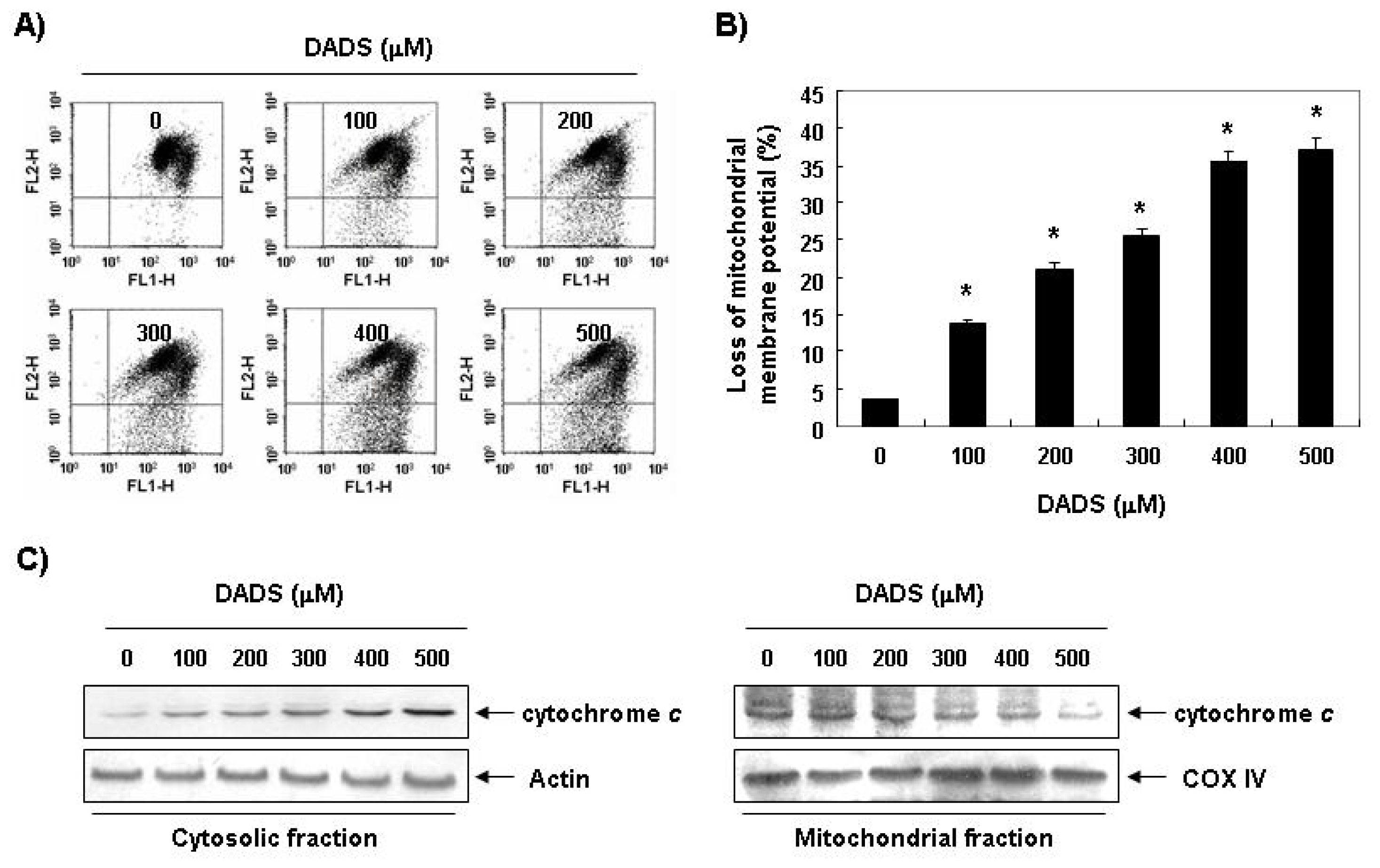
2.4. Decrease in the Levels of MMP by DADS in DU145 Cells
2.5. Activation of Caspases by DADS in DU145 Cells
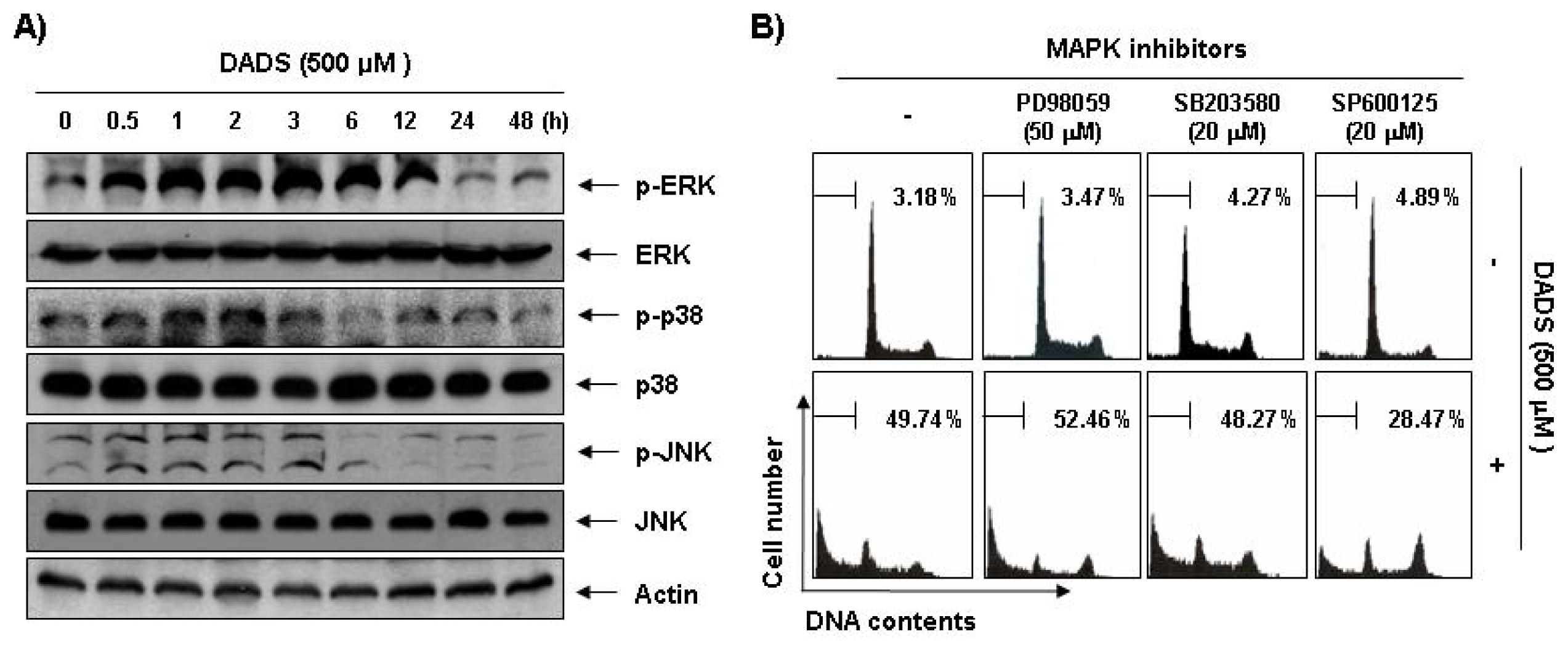
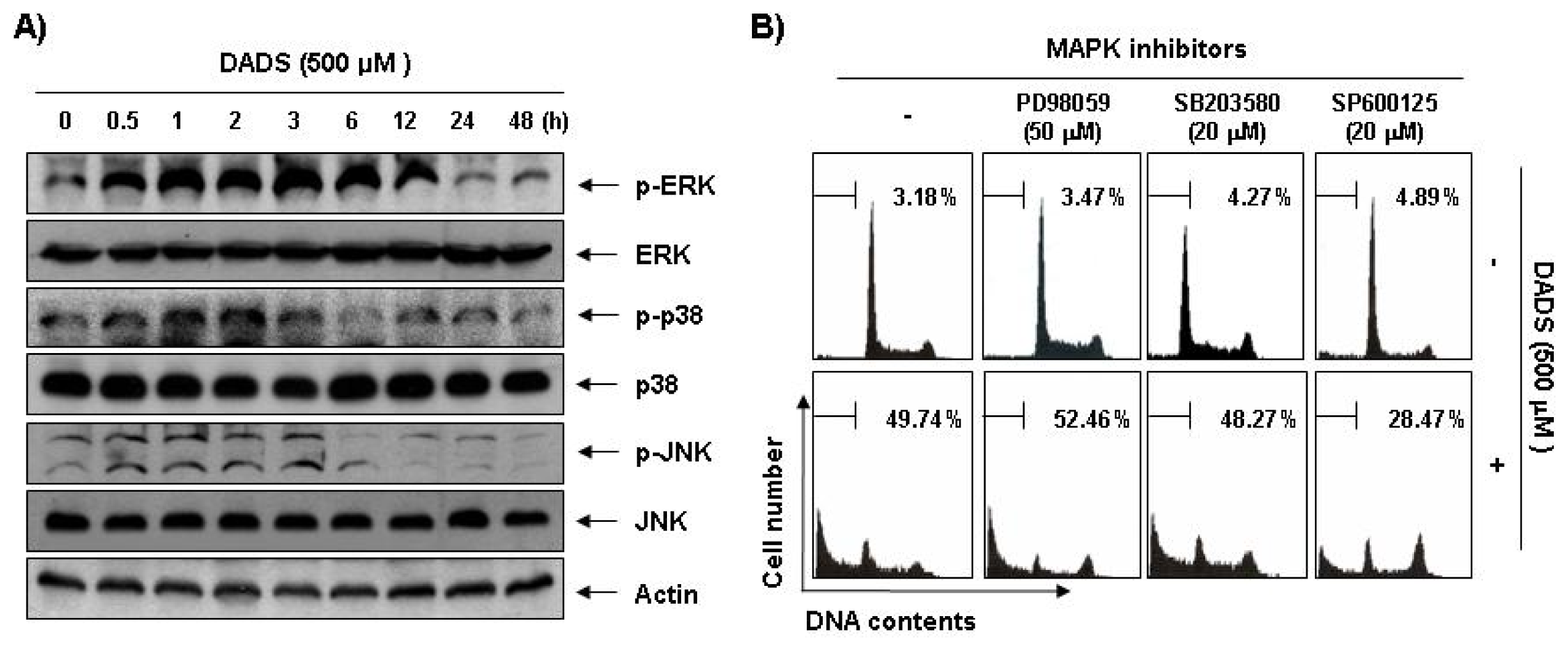
2.6. Activation of the JNK Pathway in DADS-Induced Apoptosis of DU145 Cells
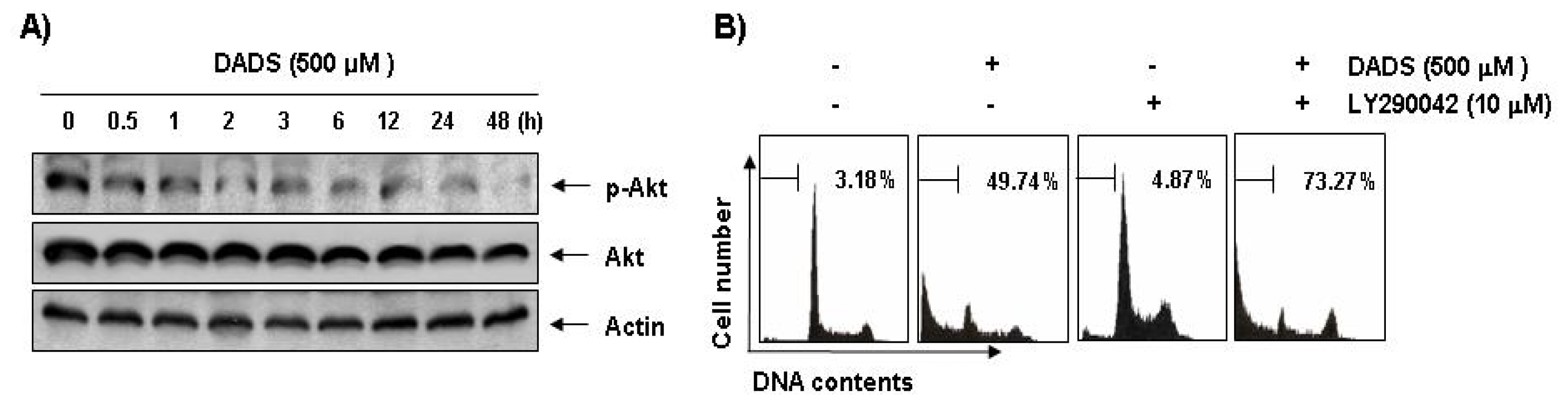
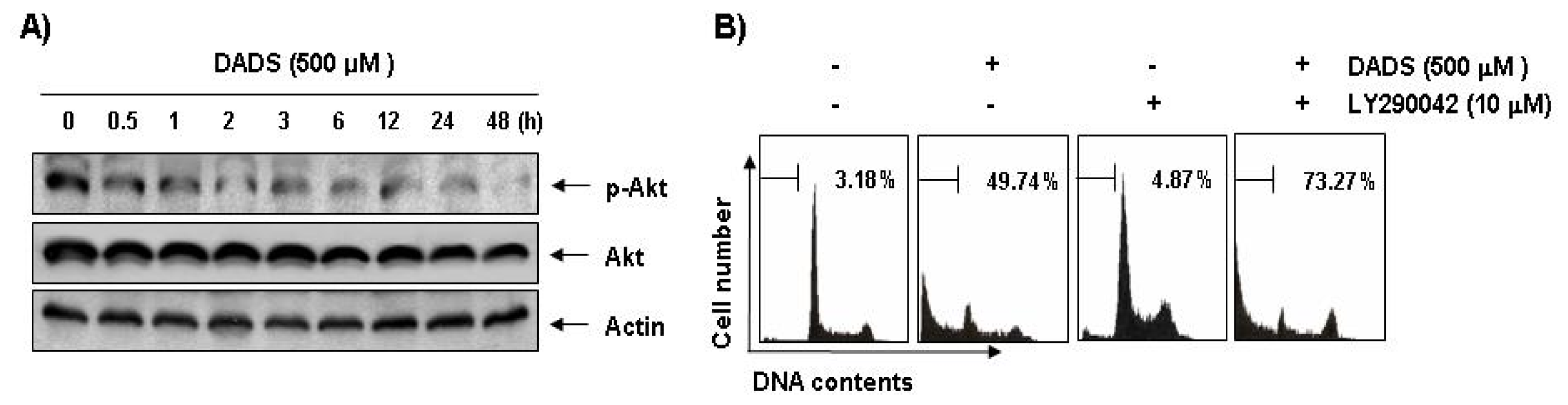
2.7. Inactivation of the PI3K/Akt Pathway in DADS-Induced Apoptosis of DU145 Cells
2.8. Discussion
3. Experimental Section
3.1. Reagents and Antibodies
3.2. Cell Culture and MTT Assay
3.3. Nuclear Staining with DAPI
3.4. DNA Fragmentation Assay
3.5. Flow Cytometric Analysis for Measurement of Sub-G1 Phase and MMP Values
3.6. Protein Extraction and Western Blotting
3.7. Caspase Activity Assay
3.8. Statistical Analysis
4. Conclusions
Acknowledgments
References
- Block, G.; Jensen, C.D.; Norkus, E.P.; Dalvi, T.B.; Wong, L.G.; McManus, J.F.; Hudes, M.L. Usage patterns, health, and nutritional status of long-term multiple dietary supplement users: A cross-sectional study. Nutr. J 2007, 6, 30. [Google Scholar]
- Bose, C.; Guo, J.; Zimniak, L.; Srivastava, S.K.; Singh, S.P.; Zimniak, P.; Singh, S.V. Critical role of allyl groups and disulfide chain in induction of Pi class glutathione transferase in mouse tissues in vivo by diallyl disulfide, a naturally occurring chemopreventive agent in garlic. Carcinogenesis 2002, 23, 1661–1665. [Google Scholar]
- Swiderski, F.; Dabrowska, M.; Rusaczonek, A.; Waszkiewicz-Robak, B. Bioactive substances of garlic and their role in dietoprophylaxis and dietotherapy. Rocz. Panstw. Zakl. Hig 2007, 58, 41–46. [Google Scholar]
- Iciek, M.; Kwiecień, I.; Włodek, L. Biological properties of garlic and garlic-derived organosulfur compounds. Environ. Mol. Mutagen 2009, 50, 247–265. [Google Scholar]
- Gayathri, R.; Gunadharini, D.N.; Arunkumar, A.; Senthilkumar, K.; Krishnamoorthy, G.; Banudevi, S.; Vignesh, R.C.; Arunakaran, J. Effects of diallyl disulfide (DADS) on expression of apoptosis associated proteins in androgen independent human prostate cancer cells (PC-3). Mol. Cell Biochem 2009, 320, 197–203. [Google Scholar]
- Aggarwal, B.B.; Shishodia, S. Molecular targets of dietary agents for prevention and therapy of cancer. Biochem. Pharmacol 2006, 71, 1397–1421. [Google Scholar]
- Nagini, S. Cancer chemoprevention by garlic and its organosulfur compounds-panacea or promise? Anticancer Agents Med. Chem 2008, 8, 313–321. [Google Scholar]
- Kaschula, C.H.; Hunter, R.; Parker, M.I. Garlic-derived anticancer agents: Structure and biological activity of ajoene. Biofactors 2010, 36, 78–85. [Google Scholar]
- Antony, M.L.; Singh, S.V. Molecular mechanisms and targets of cancer chemoprevention by garlic-derived bioactive compound diallyl trisulfide. Indian J. Exp. Biol 2011, 49, 805–816. [Google Scholar]
- Bautista, D.M.; Movahed, P.; Hinman, A.; Axelsson, H.E.; Sterner, O.; Högestätt, E.D.; Julius, D.; Jordt, S.E.; Zygmunt, P.M. Pungent products from garlic activate the sensory ion channel TRPA1. Proc. Natl. Acad. Sci. USA 2005, 102, 12248–12252. [Google Scholar]
- Shin, D.Y.; Kim, G.Y.; Kim, J.I.; Yoon, M.K.; Kwon, T.K.; Lee, S.J.; Choi, Y.W.; Kang, H.S.; Yoo, Y.H.; Choi, Y.H. Anti-invasive activity of diallyl disulfide through tightening of tight junctions and inhibition of matrix metalloproteinase activities in LNCaP prostate cancer cells. Toxicol In Vitro 2010, 24, 1569–1576. [Google Scholar]
- Tsubura, A.; Lai, Y.C.; Kuwata, M.; Uehara, N.; Yoshizawa, K. Anticancer effects of garlic and garlic-derived compounds for breast cancer control. Anticancer Agents Med. Chem 2011, 11, 249–253. [Google Scholar]
- Park, H.S.; Kim, G.Y.; Choi, I.W.; Kim, N.D.; Hwang, H.J.; Choi, Y.W.; Choi, Y.H. Inhibition of matrix metalloproteinase activities and tightening of tight junctions by diallyl disulfide in AGS human gastric carcinoma cells. J. Food Sci 2011, 76, T105–T111. [Google Scholar]
- Park, H.Y.; Kim, N.D.; Kim, G.Y.; Hwang, H.J.; Kim, B.W.; Kim, W.J.; Choi, Y.H. Inhibitory effects of diallyl disulfide on the production of inflammatory mediators and cytokines in lipopolysaccharide-activated BV2 microglia. Toxicol. Appl. Pharmacol 2012, 262, 177–184. [Google Scholar]
- Sundaram, S.G.; Milner, J.A. Diallyl disulfide suppresses the growth of human colon tumor cell xenografts in athymic nude mice. J. Nutr 1996, 126, 1355–1361. [Google Scholar]
- Tan, H.; Ling, H.; He, J.; Yi, L.; Zhou, J.; Lin, M.; Su, Q. Inhibition of ERK and activation of p38 are involved in diallyl disulfide induced apoptosis of leukemia HL-60 cells. Arch. Pharm. Res 2008, 31, 786–793. [Google Scholar]
- Wen, J.; Zhang, Y.; Chen, X.; Shen, L.; Li, G.C.; Xu, M. Enhancement of diallyl disulfide-induced apoptosis by inhibitors of MAPKs in human HepG2 hepatoma cells. Biochem. Pharmacol 2004, 68, 323–331. [Google Scholar]
- Kwon, K.B.; Yoo, S.J.; Ryu, D.G.; Yang, J.Y.; Rho, H.W.; Kim, J.S.; Park, J.W.; Kim, H.R.; Park, B.H. Induction of apoptosis by diallyl disulfide through activation of caspase-3 in human leukemia HL-60 cells. Biochem. Pharmacol 2002, 63, 41–47. [Google Scholar]
- Bottone, F.G., Jr; Baek, S.J.; Nixon, J.B.; Eling, T.E. Diallyl disulfide (DADS) induces the antitumorigenic NSAID-activated gene (NAG-1) by a p53-dependent mechanism in human colorectal HCT 116 cells. J. Nutr. 2002, 132, 773–778. [Google Scholar]
- Filomeni, G.; Aquilano, K.; Rotilio, G.; Ciriolo, M.R. Reactive oxygen species-dependent c-Jun NH2-terminal kinase/c-Jun signaling cascade mediates neuroblastoma cell death induced by diallyl disulfide. Cancer Res 2003, 63, 5940–5949. [Google Scholar]
- Arunkumar, A.; Vijayababu, M.R.; Gunadharini, N.; Krishnamoorthy, G.; Arunakaran, J. Induction of apoptosis and histone hyperacetylation by diallyl disulfide in prostate cancer cell line PC-3. Cancer Lett 2007, 251, 59–67. [Google Scholar]
- Xiao, D.; Choi, S.; Johnson, D.E.; Vogel, V.G.; Johnson, C.S.; Trump, D.L.; Lee, Y.J.; Singh, S.V. Diallyl trisulfide-induced apoptosis in human prostate cancer cells involves c-Jun N-terminal kinase and extracellular-signal regulated kinase-mediated phosphorylation of Bcl-2. Oncogene 2004, 23, 5594–5606. [Google Scholar]
- Kroemer, G.; Reed, J.C. Mitochondrial control of cell death. Nat. Med 2000, 6, 513–519. [Google Scholar]
- Jin, Z.; El-Deiry, W.S. Overview of cell death signaling pathways. Cancer Biol. Ther 2005, 4, 139–163. [Google Scholar]
- Elmore, S. Apoptosis: A review of programmed cell death. Toxicol. Pathol 2007, 35, 495–516. [Google Scholar]
- Fulda, S.; Debatin, K.M. Targeting inhibitor of apoptosis proteins (IAPs) for diagnosis and treatment of human diseases. Recent Pat. Anticancer Drug Discov 2006, 1, 81–89. [Google Scholar]
- Ashkenazi, A.; Holland, P.; Eckhardt, S.G. Ligand-based targeting of apoptosis in cancer: The potential of recombinant human apoptosis ligand 2/Tumor necrosis factor-related apoptosis-inducing ligand (rhApo2L/TRAIL). J. Clin. Oncol 2008, 26, 3621–3630. [Google Scholar]
- Röder, C.; Trauzold, A.; Kalthoff, H. Impact of death receptor signaling on the malignancy of pancreatic ductal adenocarcinoma. Eur. J. Cell Biol 2011, 90, 450–455. [Google Scholar]
- Scaffidi, C.; Schmitz, I.; Zha, J.; Korsmeyer, S.J.; Krammer, P.H.; Peter, M.E. Differential modulation of apoptosis sensitivity in CD95 type I and type II cells. J. Biol. Chem 1999, 274, 22532–22538. [Google Scholar]
- Lawen, A. Apoptosis-an introduction. Bioessays 2003, 25, 888–896. [Google Scholar]
- Brenner, D.; Mak, T.W. Mitochondrial cell death effectors. Curr. Opin. Cell Biol 2009, 21, 871–877. [Google Scholar]
- Li, H.; Zhu, H.; Xu, C.J.; Yuan, J. Cleavage of BID by caspase 8 mediates the mitochondrial damage in the Fas pathway of apoptosis. Cell 1998, 94, 491–501. [Google Scholar]
- Luo, X.; Budihardjo, I.; Zou, H.; Slaughter, C.; Wang, X. Bid, a Bcl2 interacting protein, mediates cytochrome c release from mitochondria in response to activation of cell surface death receptors. Cell 1998, 94, 481–490. [Google Scholar]
- Lazebnik, Y.A.; Kaufmann, S.H.; Desnoyers, S.; Poirier, G.G.; Earnshaw, W.C. Cleavage of poly(ADP-ribose) polymerase by a proteinase with properties like ICE. Nature 1994, 371, 346–347. [Google Scholar]
- Fukuda, K. Apoptosis-associated cleavage of β-catenin in human colon cancer and rat hepatoma cells. Int. J. Biochem. Cell Biol 1999, 31, 519–529. [Google Scholar]
- Danson, S.; Dean, E.; Dive, C.; Ranson, M. IAPs as a target for anticancer therapy. Curr. Cancer Drug Targets 2007, 7, 785–794. [Google Scholar]
- Xia, Z.; Dickens, M.; Raingeaud, J.; Davis, R.J.; Greenberg, M.E. Opposing effects of ERK and JNK-p38 MAP kinases on apoptosis. Science 1995, 270, 1326–1331. [Google Scholar]
- Boutros, T.; Chevet, E.; Metrakos, P. Mitogen-activated protein (MAP) kinase/MAP kinase phosphatase regulation: roles in cell growth, death, and cancer. Pharmacol. Rev 2008, 60, 261–310. [Google Scholar]
- Barragan, M.; Bellosillo, B.; Campas, C.; Colomer, D.; Pons, G.; Gil, J. Involvement of protein kinase C and phosphatidylinositol 3-kinase pathways in the survival of B-cell chronic lymphocytic leukemia cells. Blood 2002, 99, 2969–2976. [Google Scholar]
- Zhang, X.; Jin, B.; Huang, C. The PI3K/Akt pathway and its downstream transcriptional factors as targets for chemoprevention. Curr. Cancer Drug Targets 2007, 7, 305–316. [Google Scholar]
- Tak, J.K.; Lee, J.H.; Park, J.W. Resveratrol and piperine enhance radiosensitivity of tumor cells. BMB Rep 2012, 45, 242–246. [Google Scholar]
- Wang, M.F.; Liao, Y.F.; Hung, Y.C.; Lin, C.L.; Hour, T.C.; Lue, K.H.; Hung, H.C.; Liu, G.Y. Hydroxydibenzoylmethane induces apoptosis through repressing ornithine decarboxylase in human promyelocytic leukemia HL-60 cells. Exp. Mol. Med 2011, 43, 189–196. [Google Scholar]



© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Shin, D.Y.; Kim, G.-Y.; Lee, J.H.; Choi, B.T.; Yoo, Y.H.; Choi, Y.H. Apoptosis Induction of Human Prostate Carcinoma DU145 Cells by Diallyl Disulfide via Modulation of JNK and PI3K/AKT Signaling Pathways. Int. J. Mol. Sci. 2012, 13, 14158-14171. https://doi.org/10.3390/ijms131114158
Shin DY, Kim G-Y, Lee JH, Choi BT, Yoo YH, Choi YH. Apoptosis Induction of Human Prostate Carcinoma DU145 Cells by Diallyl Disulfide via Modulation of JNK and PI3K/AKT Signaling Pathways. International Journal of Molecular Sciences. 2012; 13(11):14158-14171. https://doi.org/10.3390/ijms131114158
Chicago/Turabian StyleShin, Dong Yeok, Gi-Young Kim, Jun Hyuk Lee, Byung Tae Choi, Young Hyun Yoo, and Yung Hyun Choi. 2012. "Apoptosis Induction of Human Prostate Carcinoma DU145 Cells by Diallyl Disulfide via Modulation of JNK and PI3K/AKT Signaling Pathways" International Journal of Molecular Sciences 13, no. 11: 14158-14171. https://doi.org/10.3390/ijms131114158