Boesenbergia pandurata Attenuates Diet-Induced Obesity by Activating AMP-Activated Protein Kinase and Regulating Lipid Metabolism

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
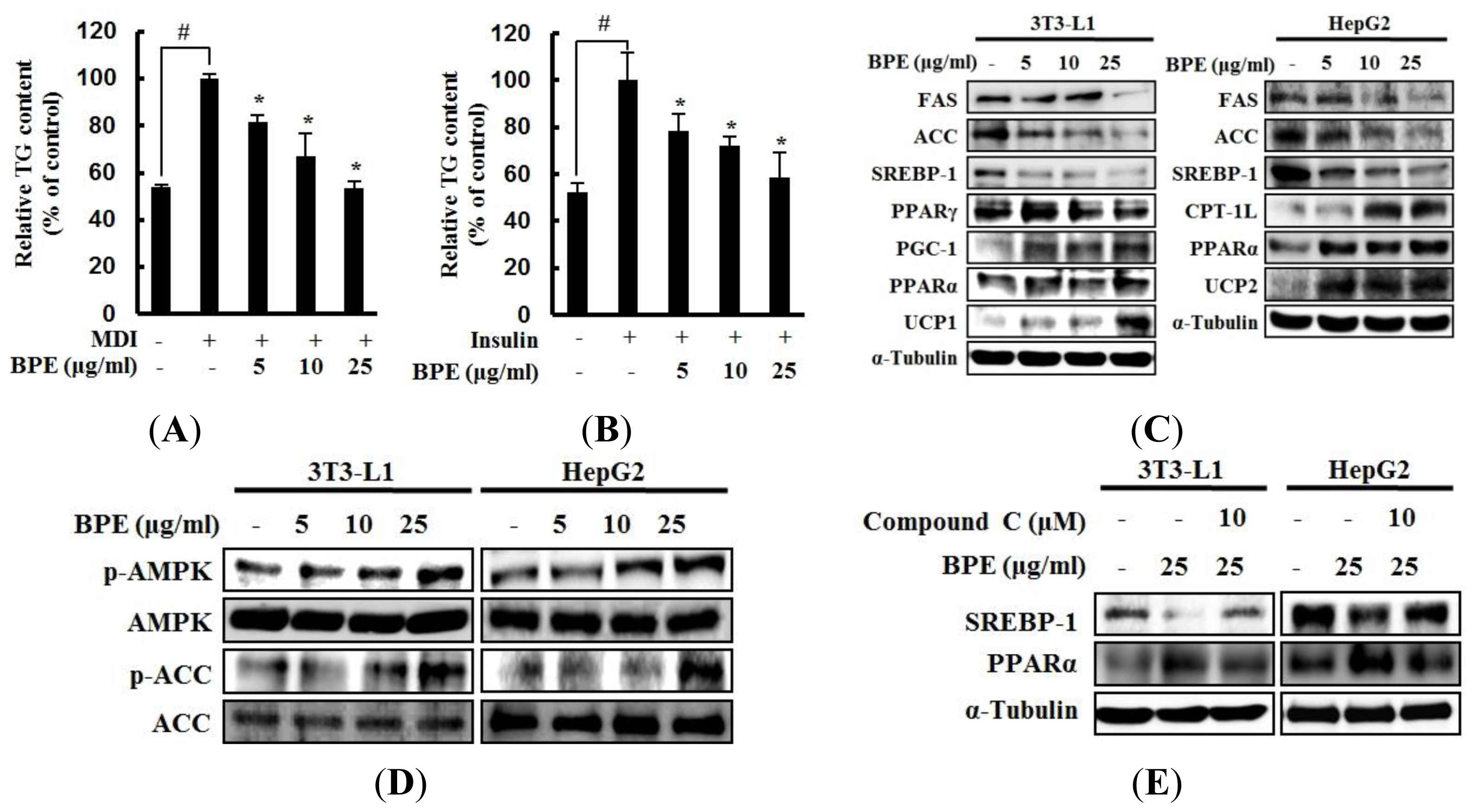
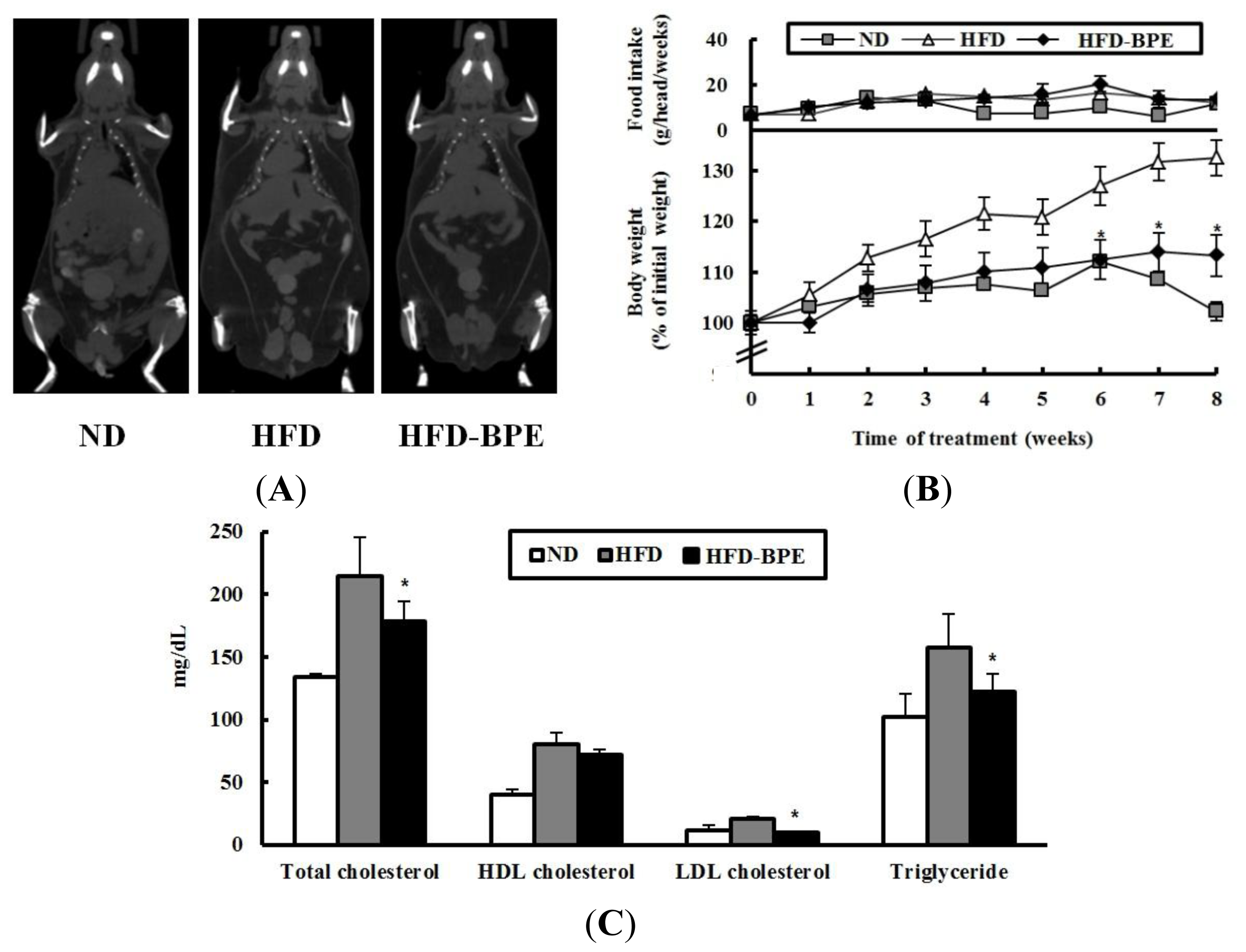
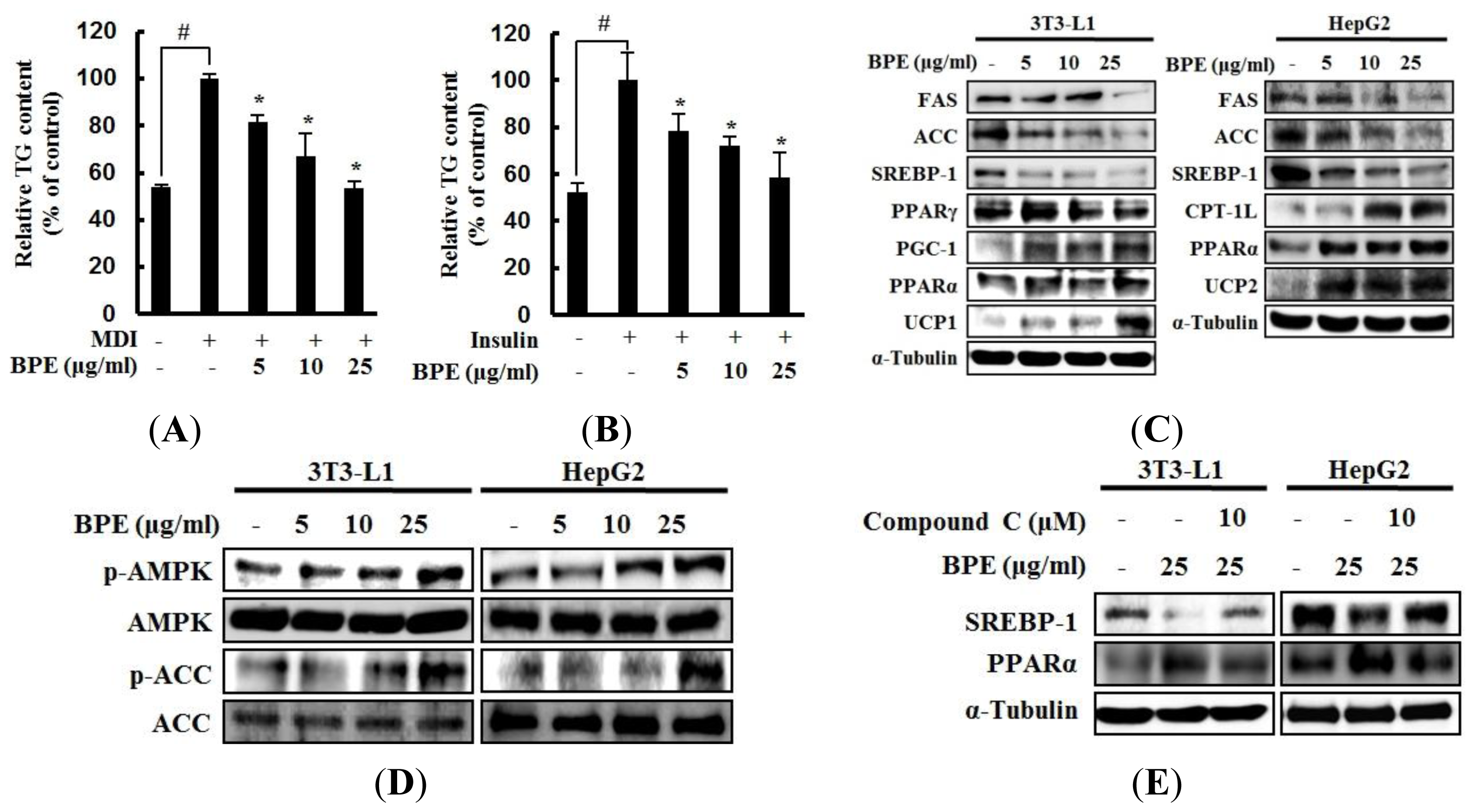
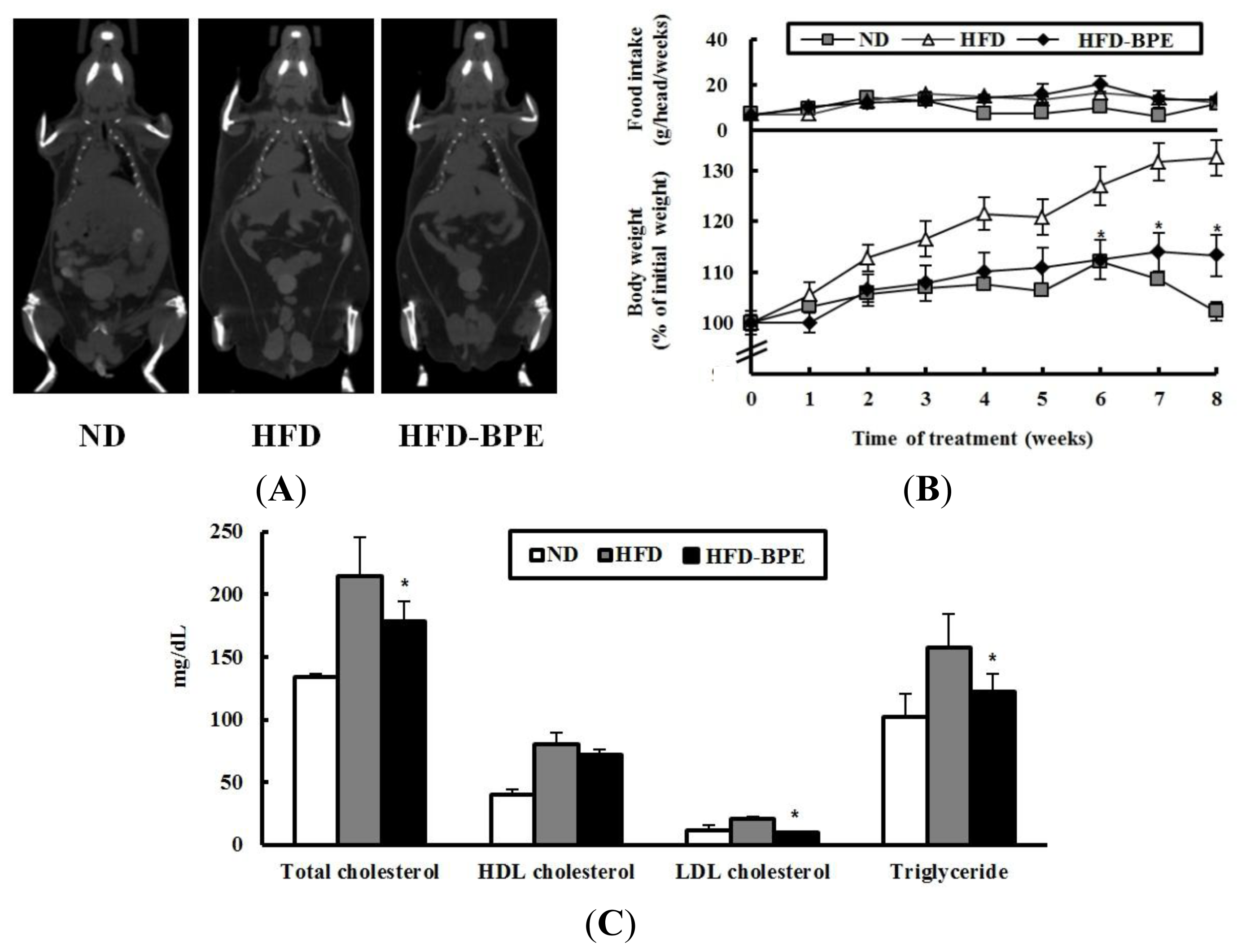
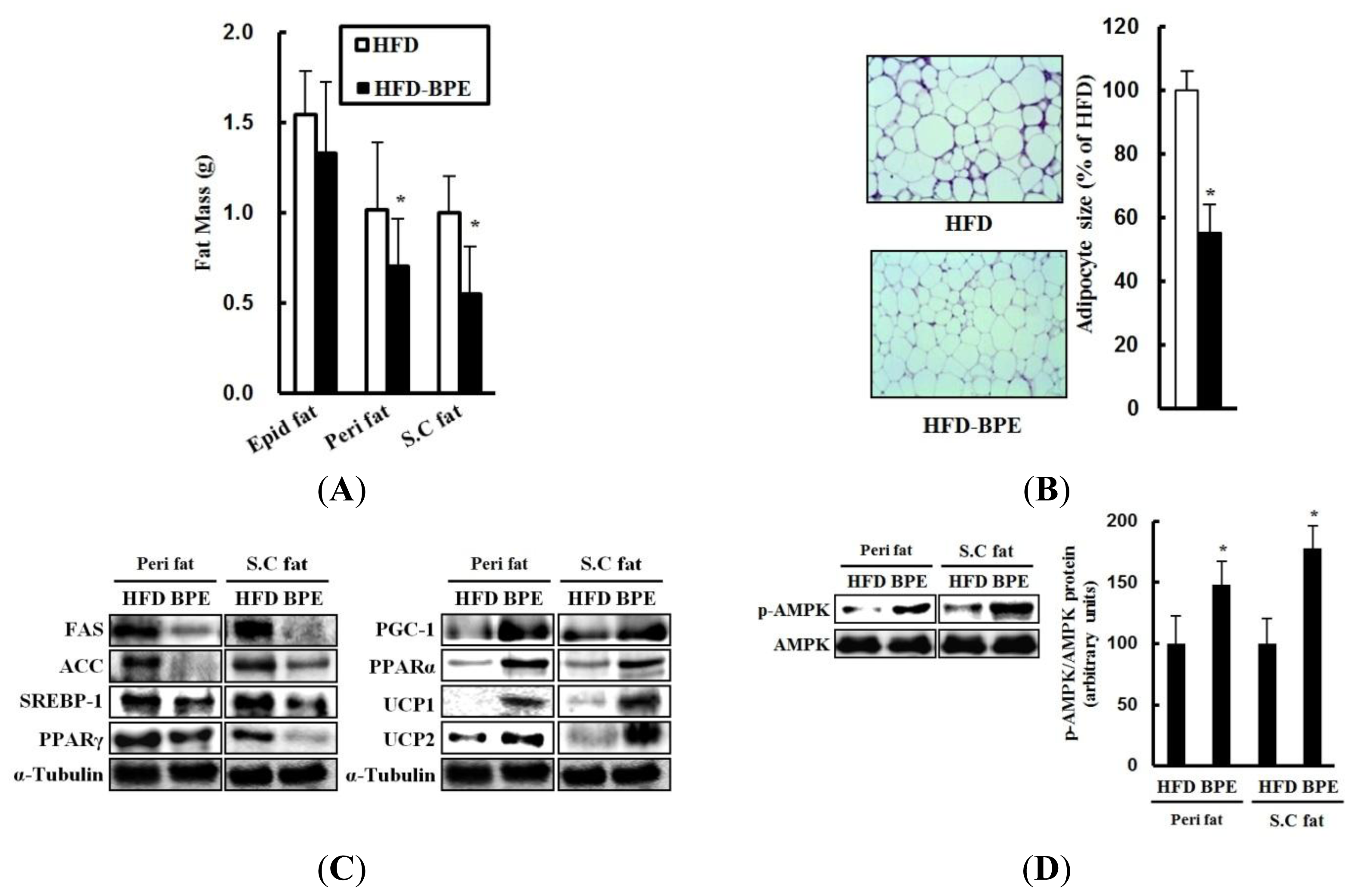
2. Results and Discussion
2.1. Adipogenesis, Hepatic Lipid Accumulation, and AMPK Activation
2.2. Body Weight and Serum Lipid Parameters in HFD-Induced Obese Mice
2.3. Fat Metabolism
2.4. Hepatic Lipid Metabolism
3. Experimental Section
3.1. Plant Material
3.2. Chemical Reagents
3.3. Cell Culture and Differentiation
3.4. Animal Experiments
3.5. Histological and Biochemical Analysis
3.6. Measurement of Hepatic Triglyceride Content
3.7. Western Blot Analysis
3.8. Statistical Analysis
4. Conclusions
Acknowledgments
- Conflict of InterestThe authors declare no conflict of interest.
References
- Després, J.P.; Lemieux, I. Abdominal obesity and metabolic syndrome. Nature 2006, 444, 881–887. [Google Scholar]
- James, P.T.; Rigby, N.; Leach, R. The obesity epidemic, metabolic syndrome and future prevention strategies. Eur. J. Cardiovasc. Prev. Rehabil 2004, 11, 3–8. [Google Scholar]
- Fujioka, K. Management of obesity as a chronic disease: Nonpharmacologic, pharmacologic, and surgical options. Obes. Res 2002, 10, S116–S123. [Google Scholar]
- Semenkovich, C.F. Insulin resistance and atherosclerosis. J. Clin. Invest 2006, 116, 1813–1822. [Google Scholar]
- Shi, Y.; Burn, P. Lipid metabolic enzymes: Emerging drug targets for the treatment of obesity. Nat. Rev. Drug. Discov 2004, 3, 695–710. [Google Scholar]
- Savage, D.B.; Petersen, K.F.; Shulman, GI. Disordered lipid metabolism and the pathogenesis of insulin resistance. Physiol. Rev 2007, 87, 507–520. [Google Scholar]
- Olefsky, J.M. Fat talks, liver and muscle listen. Cell 2008, 134, 914–916. [Google Scholar]
- Harvatine, K.J.; Bauman, D.E. SREBP1 and thyroid hormone responsive spot 14 (S14) are involved in the regulation of bovine mammary lipid synthesis during diet-induced milk fat depression and treatment with CLA. J. Nutr 2006, 136, 2468–2474. [Google Scholar]
- Boden, G. Fatty acid-induced inflammation and insulin resistance in skeletal muscle and liver. Curr. Diab. Rep 2006, 6, 177–181. [Google Scholar]
- Chen, N.; Bezzina, R.; Hinch, E. Green tea, black tea, and epigallocatechin modify body composition, improve glucose tolerance, and differentially alter metabolic gene expression in rats fed a high-fat diet. Nutr. Res 2009, 29, 784–793. [Google Scholar]
- Choi, H.; Eo, H.; Park, K. A water-soluble extract from Cucurbita moschata shows anti-obesity effects by controlling lipid metabolism in a high fat diet-induced obesity mouse model. Biochem. Biophys. Res. Commun 2007, 359, 419–425. [Google Scholar]
- Hardie, D.G. AMPK: A key regulator of energy balance in the single cell and the whole organism. Int. J. Obes. (Lond.) 2008, 32, S7–S12. [Google Scholar]
- Ouadda, A.B.; Levy, E.; Ziv, E. Increased hepatic lipogenesis in insulin resistance and type 2 diabetes is associated with AMPK signalling pathway up-regulation in Psammomys obesus. Biosci. Rep 2009, 29, 283–292. [Google Scholar]
- Osler, M.E.; Zierath, J.R. Adenosine 5′-monophosphate-activated protein kinase regulation of fatty acid oxidation in skeletal muscle. Endocrinology 2008, 149, 935–941. [Google Scholar]
- Hasani-Ranjbar, S.; Nayebi, N.; Larijani, B.; Abdollahi, M. A systematic review of the efficacy and safety of herbal medicines used in the treatment of obesity. World J. Gastroenterol 2009, 15, 3073–3085. [Google Scholar]
- Lagouge, M.; Argmann, C.; Gerhart-Hines, Z. Resveratrol improves mitochondrial function and protects against metabolic disease by activating SIRT1 and PGC-1alpha. Cell 2006, 127, 1109–1122. [Google Scholar]
- Lee, M.S.; Kim, C.T.; Kim, Y. Green tea (-)-epigallocatechin-3-gallate reduces body weight with regulation of multiple genes expression in adipose tissue of diet-induced obese mice. Ann. Nutr. Metab 2009, 54, 151–157. [Google Scholar]
- Kim, W.S.; Lee, Y.S.; Cha, S.H. Berberine improves lipid dysregulation in obesity by controlling central and peripheral AMPK activity. Am. J. Physiol. Endocrinol. Metab 2009, 296, E812–E819. [Google Scholar]
- Yun, J.M.; Kwon, H.; Hwang, J.K. In vitro anti-inflammatory activity of panduratin A isolated from Kaempferia pandurata in RAW264.7 cells. Planta Med 2003, 69, 1102–1108. [Google Scholar]
- Shindo, K.; Kato, M.; Kinoshita, A.; Kobayashi, A.; Koike, Y. Analysis of antioxidant activities contained in the Boesenbergia pandurata Schult. Rhizome. Biosci. Biotechnol. Biochem 2006, 70, 2281–2284. [Google Scholar]
- Lee, Y.S.; Kim, W.S.; Kim, K.H. Berberine, a natural plant product, activates AMP-activated protein kinase with beneficial metabolic effects in diabetic and insulin-resistant states. Diabetes 2006, 55, 2256–2264. [Google Scholar]
- Kim, D.Y.; Lee, M.S.; Jo, K.; Lee, K.-E. Therapeutic potential of panduratin A, LKB1-dependent AMP-activated protein kinase stimulator, with activation of PPARα/δ for the treatment of obesity. Diabetes. Obes. Metab 2011, 13, 584–593. [Google Scholar]
- Wellen, K.E.; Hotamisligil, G.S. Obesity-induced inflammatory changes in adipose tissue. J. Clin. Invest 2003, 112, 1785–1788. [Google Scholar]
- Amini, Z.; Boyd, B.; Doucet, J.; Ribnicky, D.M.; Stephens, J.M. St. John’s Wort inhibits adipocyte differentiation and induces insulin resistance in adipocytes. Biochem. Biophys. Res. Commun 2009, 338, 146–149. [Google Scholar]
- Han, C.; Wang, J.; Li, L.; Zhang, Z.; Wang, L.; Pan, Z. The role of insulin and glucose in goose primary hepatocyte triglyceride accumulation. J. Exp. Biol 2009, 212, 1553–1558. [Google Scholar]
- Tuchinda, P.; Reutrakul, V.; Claeson, P. Anti-inflammatory cyclohexenyl chalcone derivatives in Boesenbergia pandurata. Phytochemistry 2002, 59, 169–173. [Google Scholar]

© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Kim, D.-Y.; Kim, M.-S.; Sa, B.-K.; Kim, M.-B.; Hwang, J.-K. Boesenbergia pandurata Attenuates Diet-Induced Obesity by Activating AMP-Activated Protein Kinase and Regulating Lipid Metabolism. Int. J. Mol. Sci. 2012, 13, 994-1005. https://doi.org/10.3390/ijms13010994
Kim D-Y, Kim M-S, Sa B-K, Kim M-B, Hwang J-K. Boesenbergia pandurata Attenuates Diet-Induced Obesity by Activating AMP-Activated Protein Kinase and Regulating Lipid Metabolism. International Journal of Molecular Sciences. 2012; 13(1):994-1005. https://doi.org/10.3390/ijms13010994
Chicago/Turabian StyleKim, Dae-Young, Myung-Suk Kim, Bo-Kyung Sa, Mi-Bo Kim, and Jae-Kwan Hwang. 2012. "Boesenbergia pandurata Attenuates Diet-Induced Obesity by Activating AMP-Activated Protein Kinase and Regulating Lipid Metabolism" International Journal of Molecular Sciences 13, no. 1: 994-1005. https://doi.org/10.3390/ijms13010994