Potentiation of Anticancer Drugs: Effects of Pentoxifylline on Neoplastic Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Results
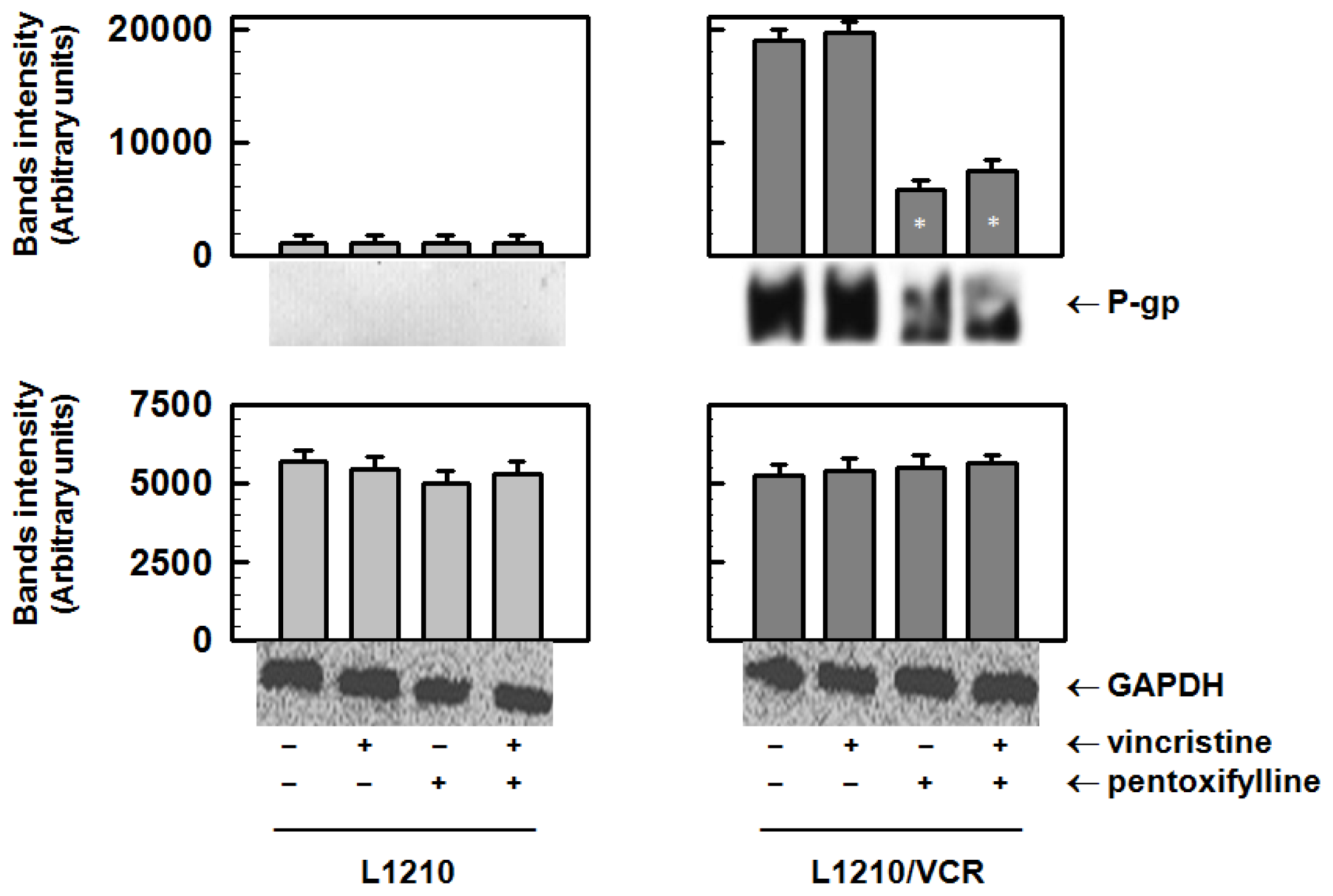
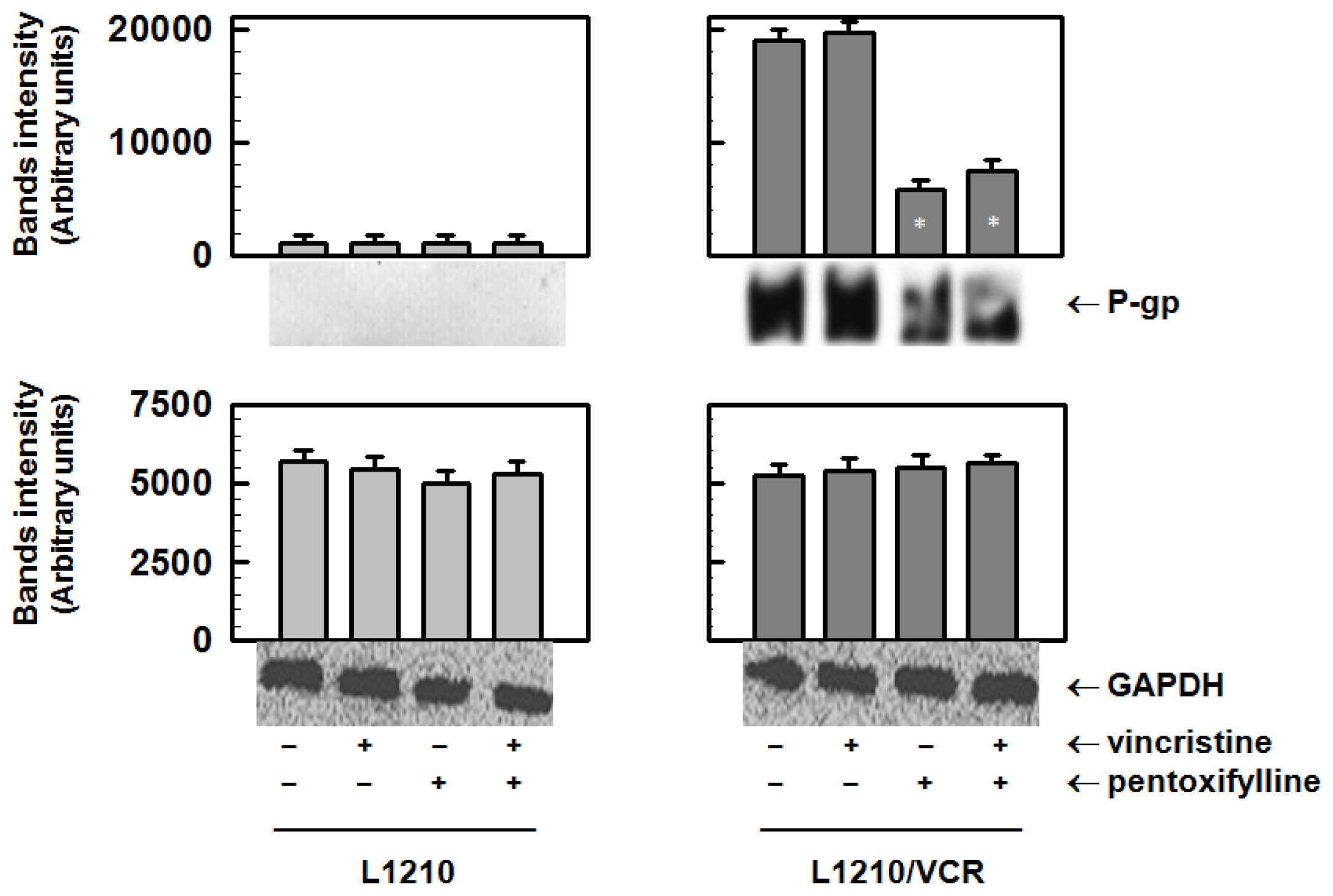
2.1.1. Pentoxifylline Reduces P-glycoprotein Protein Levels
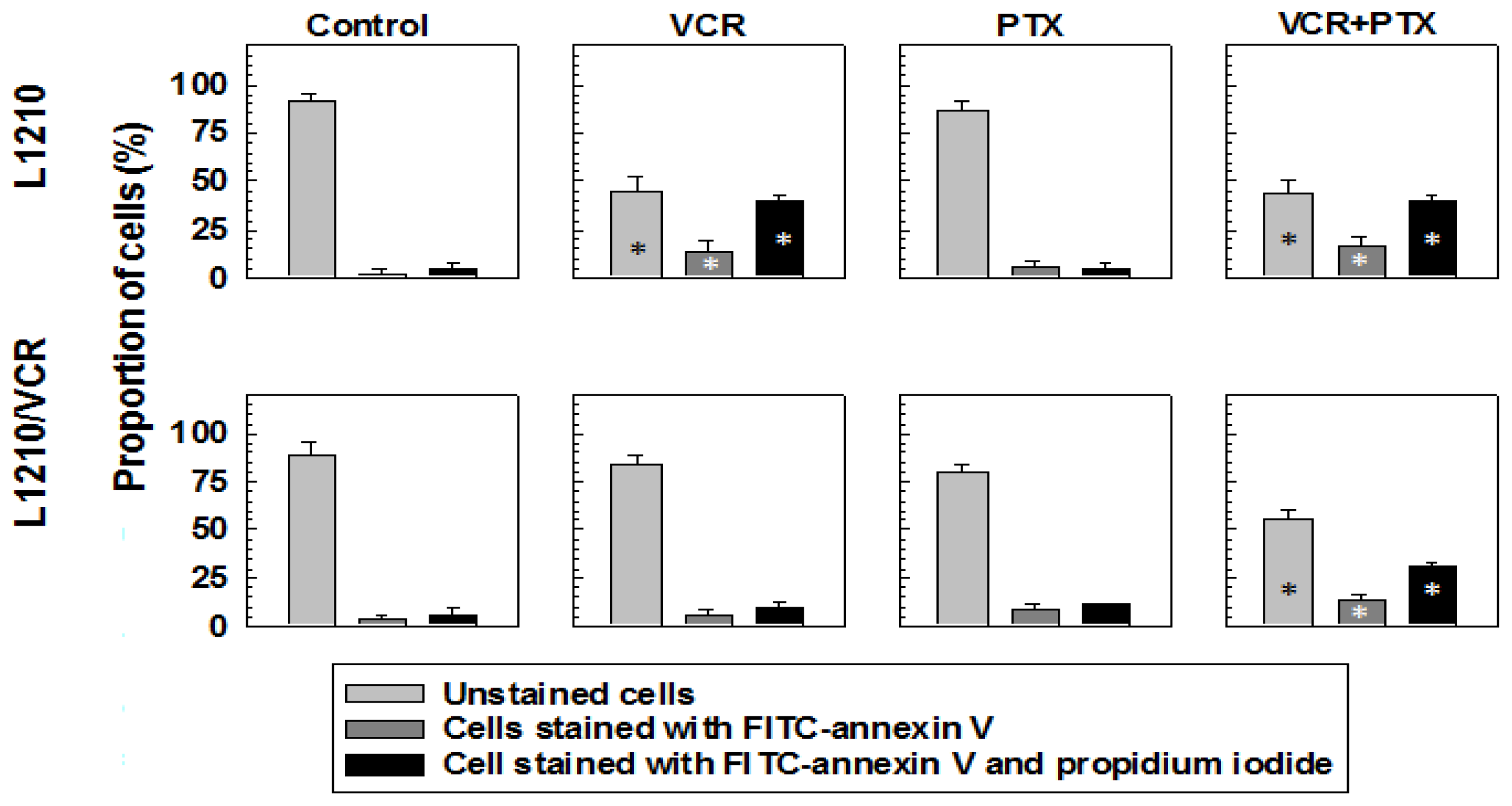
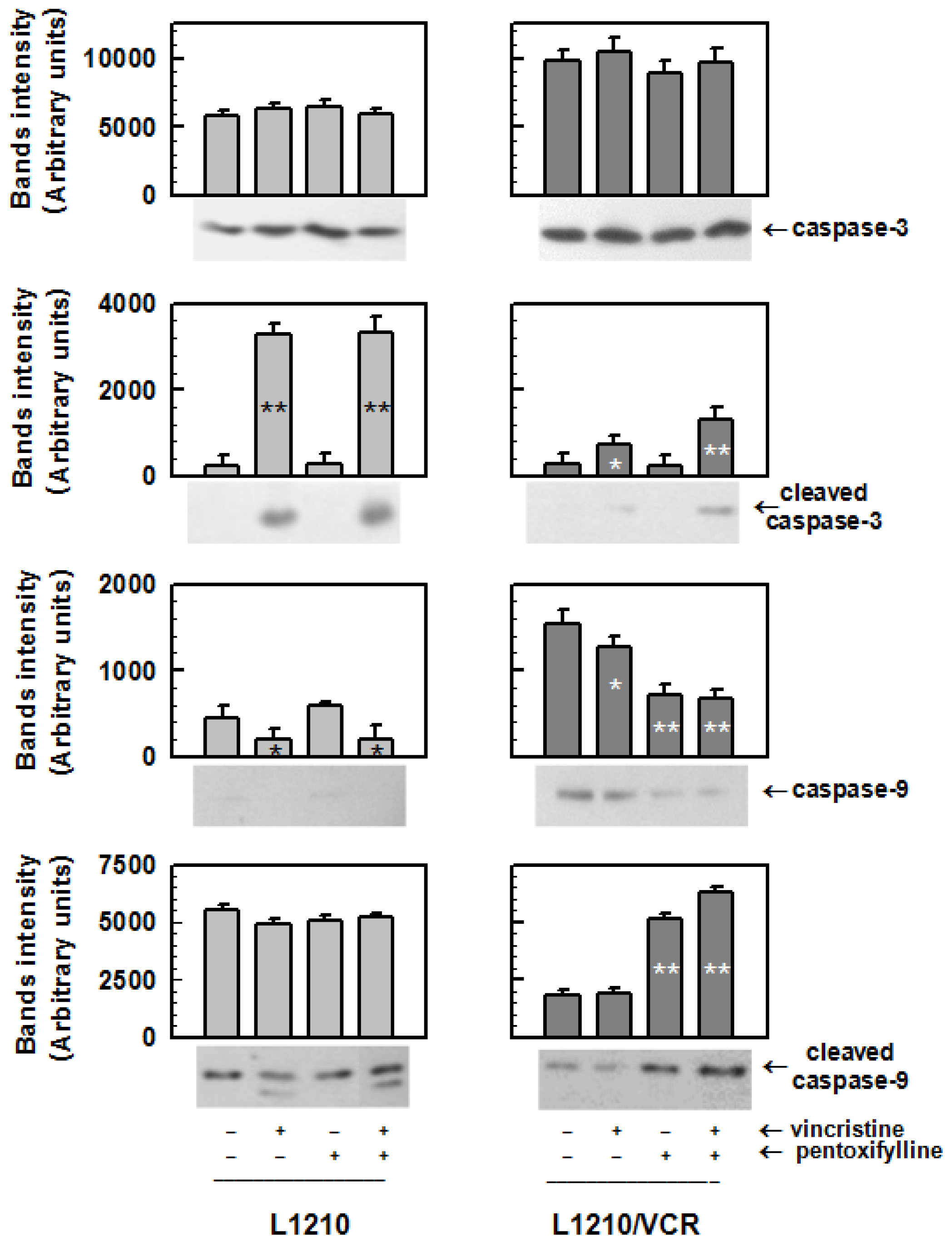
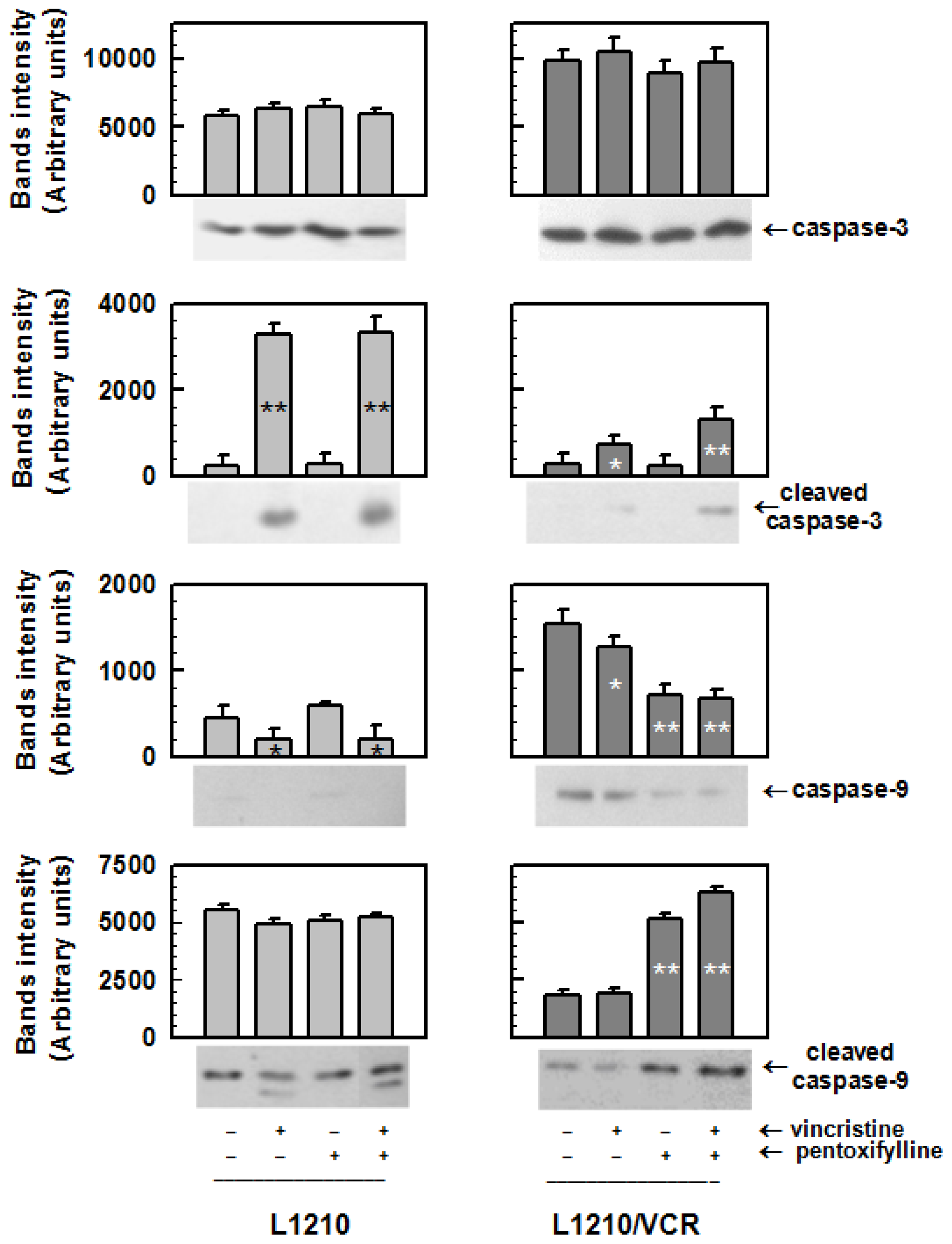
2.1.2. Drug Reversal Effects of Pentoxifylline Are Associated with the Stimulation of Apoptosis
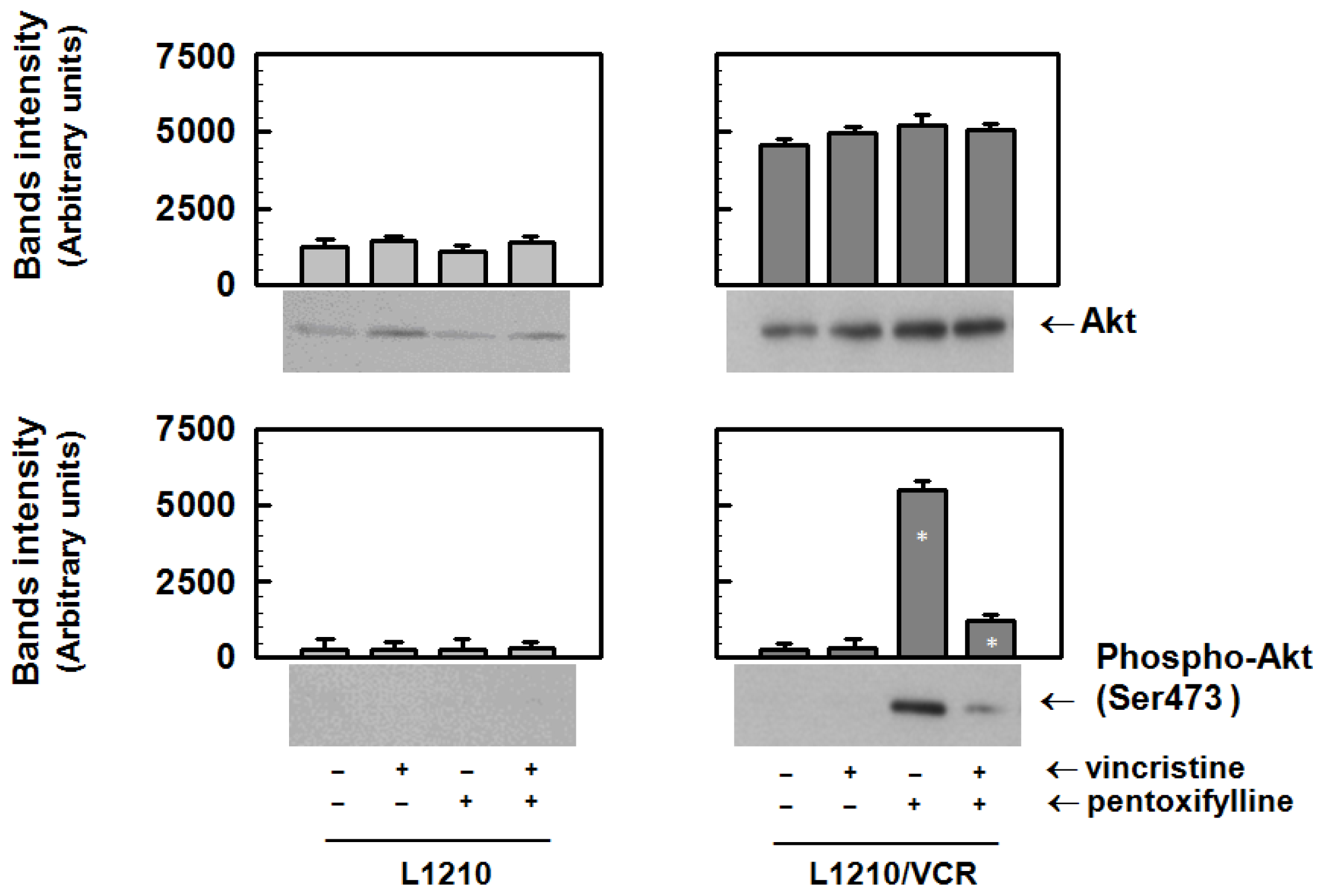
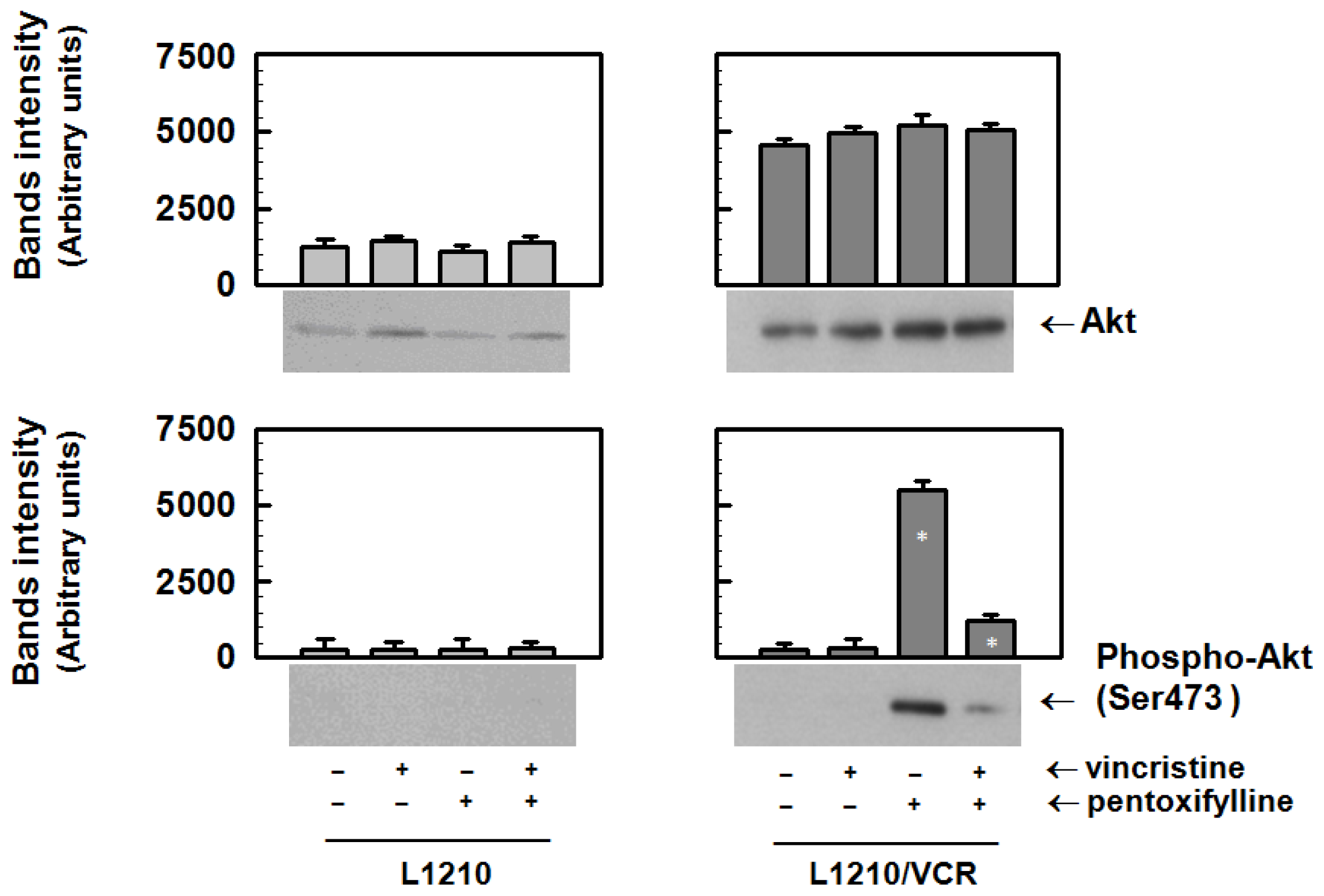
2.1.3. Pentoxifylline Influences the Activation of Akt Kinase in Resistant L1210/VCR Cells
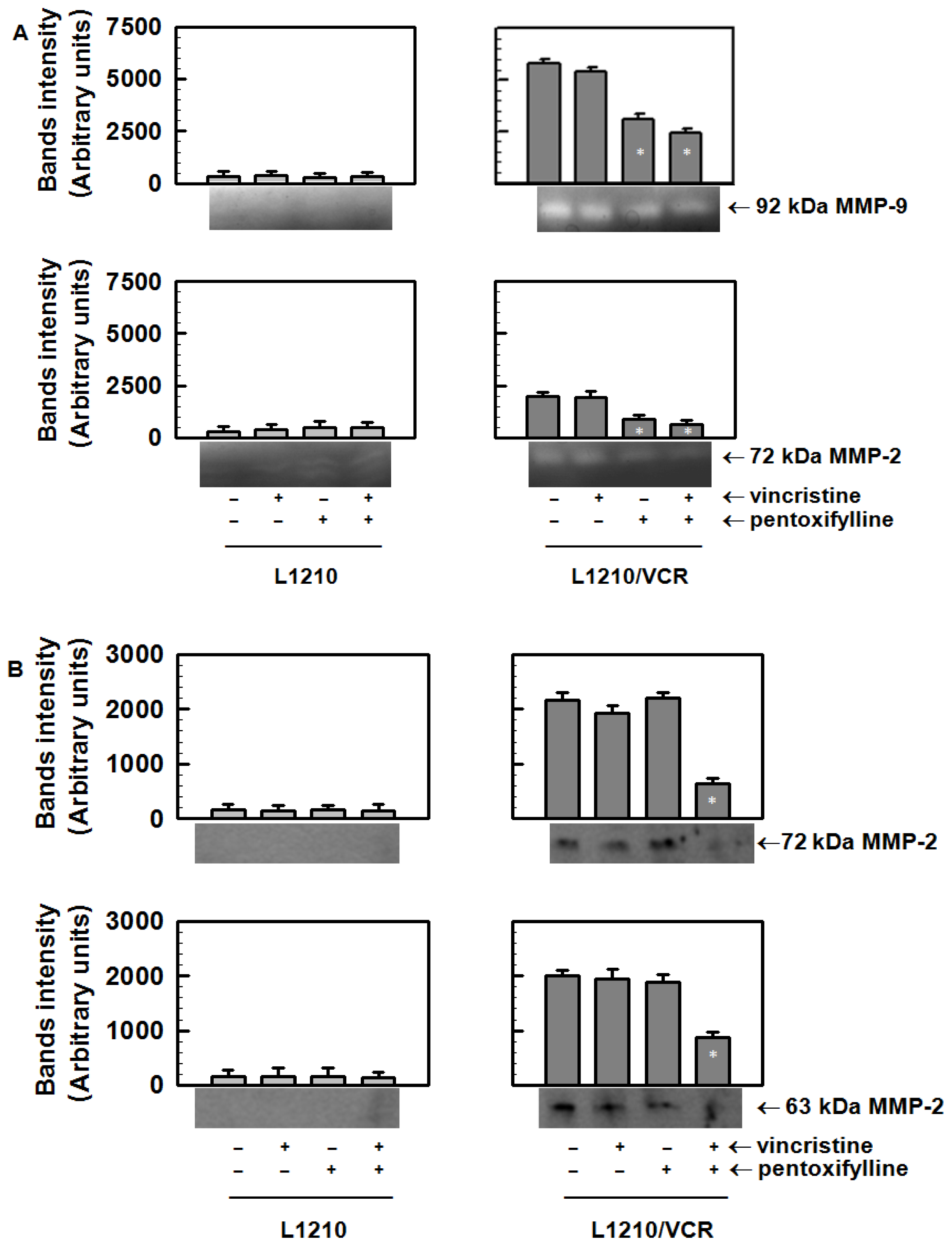
2.1.4. Pentoxifylline Affects the Release of Matrix Metalloproteinases from L1210/VCR Cells
2.2. Discussion
3. Experimental Section
3.1. Materials
3.2. Cell Culture and Exposure of Cells to Pentoxifylline and Vincristine
3.3. Preparation of Cellular Protein Fractions and Western Blot Analysis
3.4. Detection of Apoptotic Cells by FACS Analysis
3.5. Effect of Pentoxifylline on the Release of Metalloproteinases from Sensitive L1210 and Drug-Resistant L1210/VCR Cells
3.6. Gelatin Zymography
3.7. Statistical Analysis
4. Conclusions
Acknowledgments
References
- Breier, A.; Barancik, M.; Sulova, Z.; Uhrik, B. P-glycoprotein—implications of metabolism of neoplastic cells and cancer therapy. Curr. Cancer Drug Targets 2005, 5, 457–468. [Google Scholar]
- Colabufo, N.A.; Berardi, F.; Cantore, M.; Contino, M.; Inglese, C.; Niso, M.; Perrone, R. Perspectives of P-glycoprotein modulating agents in oncology and neurodegenerative diseases: Pharmaceutical, biological, and diagnostic potentials. J. Med. Chem 2010, 53, 1883–1897. [Google Scholar]
- Butanda-Ochoa, A.; Hojer, G.; Morales-Tlalpan, V.; Diaz-Munoz, M. Recognition and activation of ryanodine receptors by purines. Curr. Med. Chem 2006, 13, 647–657. [Google Scholar]
- Duman, D.G.; Ozdemir, F.; Birben, E.; Keskin, O.; Eksioglu-Demiralp, E.; Celikel, C.; Kalayci, O.; Kalayci, C. Effects of pentoxifylline on TNF-alpha production by peripheral blood mononuclear cells in patients with nonalcoholic steatohepatitis. Dig. Dis. Sci 2007, 52, 2520–2524. [Google Scholar]
- Klotz, K.N. Adenosine receptors and their ligands. Naunyn Schmiedebergs Arch. Pharmacol 2000, 362, 382–391. [Google Scholar]
- Miyamoto, K.; Sakai, R.; Kurita, M.; Ohmae, S.; Sanae, F.; Sawanishi, H.; Hasegawa, T.; Takagi, K. Effects of alkyl substituents of xanthine on phosphodiesterase isoenzymes. Biol. Pharm. Bull 1995, 18, 431–434. [Google Scholar]
- Alas, S.; Bonavida, B.; Emmanouilides, C. Potentiation of fludarabine cytotoxicity on non-Hodgkin’s lymphoma by pentoxifylline and rituximab. Anticancer Res 2000, 20, 2961–2966. [Google Scholar]
- Rauko, P.; Sedlak, J.; Duraj, J.; Szekeres, M.F.; Novotny, L. Pentoxifylline stimulates drug-induced apoptosis in leukemic cells. Neoplasma 1998, 45, 296–300. [Google Scholar]
- Drobna, Z.; Stein, U.; Walther, W.; Barancik, M.; Breier, A. Pentoxifylline influences drug transport activity of P-glycoprotein and decreases mdrl gene expression in multidrug resistant mouse leukemic L1210/VCR cells. Gen. Physiol. Biophys 2002, 21, 103–109. [Google Scholar]
- Stefankova, Z.; Barancik, M.; Breier, A. Overcoming of P-glycoprotein mediated vincristine resistance of L1210/VCR mouse leukemic cells could be induced by pentoxifyline but not by theophylline and caffeine. Neoplasma 1996, 43, 11–15. [Google Scholar]
- Gibalova, L.; Sedlak, J.; Labudova, M.; Barancik, M.; Rehakova, A.; Breier, A.; Sulova, Z. Multidrug resistant P-glycoprotein positive L1210/VCR cells are also cross-resistant to cisplatin via a mechanism distinct from P-glycoprotein-mediated drug efflux activity. Gen. Physiol. Biophys 2009, 28, 391–403. [Google Scholar]
- Bravo-Cuellar, A.; Ortiz-Lazareno, P.C.; Lerma-Diaz, J.M.; Dominguez-Rodriguez, J.R.; Jave-Suarez, L.F.; Aguilar-Lemarroy, A.; del Toro-Arreola, S.; de Celis-Carrillo, R.; Sahagun-Flores, J.E.; de Alba-Garcia, J.E.; et al. Sensitization of cervix cancer cells to Adriamycin by Pentoxifylline induces an increase in apoptosis and decrease senescence. Mol. Cancer 2010, 9, 114. [Google Scholar]
- Barancik, M.; Bohacova, V.; Sedlak, J.; Sulova, Z.; Breier, A. LY294,002, a specific inhibitor of PI3K/Akt kinase pathway, antagonizes P-glycoprotein-mediated multidrug resistance. Eur. J. Pharm. Sci 2006, 29, 426–434. [Google Scholar]
- Zhang, J.; Ling, Y.; Tang, L.; Luo, B.; Chacko, B.K.; Patel, R.P.; Fallon, M.B. Pentoxifylline attenuation of experimental hepatopulmonary syndrome. J. Appl. Physiol 2007, 102, 949–955. [Google Scholar]
- Dua, P.; Gude, R.P. Antiproliferative and antiproteolytic activity of pentoxifylline in cultures of B16F10 melanoma cells. Cancer Chemother. Pharmacol 2006, 58, 195–202. [Google Scholar]
- Liang, Y.; O’Driscoll, L.; McDonnell, S.; Doolan, P.; Oglesby, I.; Duffy, K.; O’Connor, R.; Clynes, M. Enhanced in vitro invasiveness and drug resistance with altered gene expression patterns in a human lung carcinoma cell line after pulse selection with anticancer drugs. Int. J. Cancer 2004, 111, 484–493. [Google Scholar]
- Polekova, L.; Barancik, M.; Mrazova, T.; Pirker, R.; Wallner, J.; Sulova, Z.; Breier, A. Adaptation of mouse leukemia cells L1210 to vincristine. Evidence for expression of P-glycoprotein. Neoplasma 1992, 39, 73–77. [Google Scholar]
- Breier, A.; Barancik, M.; Stefankova, Z.; Uhrik, B.; Tribulova, N. Effect of pentoxifylline on P-glycoprotein mediated vincristine resistance of L1210 mouse leukemic cell line. Neoplasma 1994, 41, 297–303. [Google Scholar]
- Kupsakova, I.; Rybar, A.; Docolomansky, P.; Drobna, Z.; Stein, U.; Walther, W.; Barancik, M.; Breier, A. Reversal of P-glycoprotein mediated vincristine resistance of L1210/VCR cells by analogues of pentoxifylline. A QSAR study. Eur. J. Pharm. Sci 2004, 21, 283–293. [Google Scholar]
- Chua, B.T.; Gallego-Ortega, D.; de Molina, A.R.; Ullrich, A.; Lacal, J.C.; Downward, J. Regulation of Akt(ser473) phosphorylation by choline kinase in breast carcinoma cells. Mol. Cancer 2009, 8, 131. [Google Scholar]
- Viladkar, A.; Chitnis, M. In vitro effects of pentoxifylline and doxorubicin on cell survival and DNA damage in sensitive and MDR-P388 leukemia cells. Cancer Biother 1994, 9, 143–151. [Google Scholar]
- Viladkar, A.; Juvekar, A.; Chitnis, M.; Advani, S. Amelioration of doxorubicin resistance by pentoxifylline in human chronic myeloid leukemia cells in vitro. Sel. Cancer Ther 1991, 7, 119–126. [Google Scholar]
- Fantin, M.; Quintieri, L.; Kusz, E.; Kis, E.; Glavinas, H.; Floreani, M.; Padrini, R.; Duda, E.; Vizler, C. Pentoxifylline and its major oxidative metabolites exhibit different pharmacological properties. Eur. J. Pharmacol 2006, 535, 301–309. [Google Scholar]
- Bu, R.; Uddin, S.; Bavi, P.; Hussain, A.R.; Al-Dayel, F.; Ghourab, S.; Ahmed, M.; Al-Kuraya, K.S. HGF/c-Met pathway has a prominent role in mediating antiapoptotic signals through AKT in epithelial ovarian carcinoma. Lab. Invest 2010, 91, 124–137. [Google Scholar]
- Rishi, L.; Gahlot, S.; Kathania, M.; Majumdar, S. Pentoxifylline induces apoptosis in vitro in cutaneous T cell lymphoma (HuT-78) and enhances FasL mediated killing by upregulating Fas expression. Biochem. Pharmacol 2009, 77, 30–45. [Google Scholar]
- Campos, L.; Oriol, P.; Sabido, O.; Guyotat, D. Simultaneous expression of P-glycoprotein and BCL-2 in acute myeloid leukemia blast cells. Leuk. Lymphoma 1997, 27, 119–125. [Google Scholar]
- del Principe, M.I.; del Poeta, G.; Maurillo, L.; Buccisano, F.; Venditti, A.; Tamburini, A.; Bruno, A.; Cox, M.C.; Suppo, G.; Tendas, A.; et al. P-glycoprotein and BCL-2 levels predict outcome in adult acute lymphoblastic leukaemia. Br. J. Haematol 2003, 121, 730–738. [Google Scholar]
- Stamenkovic, I. Matrix metalloproteinases in tumor invasion and metastasis. Semin. Cancer Biol 2000, 10, 415–433. [Google Scholar]
- Song, J.H.; Kim, S.H.; Cho, D.; Lee, I.K.; Kim, H.J.; Kim, T.S. Enhanced invasiveness of drug-resistant acute myeloid leukemia cells through increased expression of matrix metalloproteinase-2. Int. J. Cancer 2009, 125, 1074–1081. [Google Scholar]
- Pereda, M.P.; Ledda, M.F.; Goldberg, V.; Chervin, A.; Carrizo, G.; Molina, H.; Muller, A.; Renner, U.; Podhajcer, O.; Arzt, E.; et al. High levels of matrix metalloproteinases regulate proliferation and hormone secretion in pituitary cells. J. Clin. Endocrinol. Metab 2000, 85, 263–269. [Google Scholar]
- Ohsaki, Y.; Ishida, S.; Fujikane, T.; Kikuchi, K. Pentoxifylline potentiates the antitumor effect of cisplatin and etoposide on human lung cancer cell lines. Oncology 1996, 53, 327–333. [Google Scholar]
- Magnusson, M.; Hoglund, P.; Johansson, K.; Jonsson, C.; Killander, F.; Malmstrom, P.; Weddig, A.; Kjellen, E. Pentoxifylline and vitamin E treatment for prevention of radiation-induced side-effects in women with breast cancer: A phase two, double-blind, placebo-controlled randomised clinical trial (Ptx-5). Eur. J. Cancer 2009, 45, 2488–2495. [Google Scholar]
- Seres, M.; Ditte, P.; Breier, A.; Sulova, Z. Effect of thapsigargin on P-glycoprotein-negative and P-glycoprotein-positive L1210 mouse leukaemia cells. Gen. Physiol. Biophys 2010, 29, 396–401. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem 1976, 72, 248–254. [Google Scholar]
- Peters, W.H.; Fleuren-Jakobs, A.M.; Kamps, K.M.; de Pont, J.J.; Bonting, S.L. Lowry protein determination on membrane preparations: Need for standardization by amino acid analysis. Anal. Biochem 1982, 124, 349–352. [Google Scholar]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar]
- Towbin, H.; Staehelin, T.; Gordon, J. Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: Procedure and some applications. Proc. Natl. Acad. Sci. USA 1979, 76, 4350–4354. [Google Scholar]


© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Barancik, M.; Bohacova, V.; Gibalova, L.; Sedlak, J.; Sulova, Z.; Breier, A. Potentiation of Anticancer Drugs: Effects of Pentoxifylline on Neoplastic Cells. Int. J. Mol. Sci. 2012, 13, 369-382. https://doi.org/10.3390/ijms13010369
Barancik M, Bohacova V, Gibalova L, Sedlak J, Sulova Z, Breier A. Potentiation of Anticancer Drugs: Effects of Pentoxifylline on Neoplastic Cells. International Journal of Molecular Sciences. 2012; 13(1):369-382. https://doi.org/10.3390/ijms13010369
Chicago/Turabian StyleBarancik, Miroslav, Viera Bohacova, Lenka Gibalova, Jan Sedlak, Zdena Sulova, and Albert Breier. 2012. "Potentiation of Anticancer Drugs: Effects of Pentoxifylline on Neoplastic Cells" International Journal of Molecular Sciences 13, no. 1: 369-382. https://doi.org/10.3390/ijms13010369
APA StyleBarancik, M., Bohacova, V., Gibalova, L., Sedlak, J., Sulova, Z., & Breier, A. (2012). Potentiation of Anticancer Drugs: Effects of Pentoxifylline on Neoplastic Cells. International Journal of Molecular Sciences, 13(1), 369-382. https://doi.org/10.3390/ijms13010369