microRNA Response to Listeria monocytogenes Infection in Epithelial Cells

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. Eukaryotic Cell Culture
2.3. LLO Purification
2.4. Infection Assays and LLO Treatment
2.5. RNA Isolation
2.6. miRNA Microarray
2.7. Reverse Transcription Reaction and Quantitative Real-Time PCR Analysis
2.8. Statistical Data Analysis of Infection Experiments
3. Results
3.1. L. monocytogenes Differentially Induces miRNAs Dependent on Cellular and Subcellular Localization
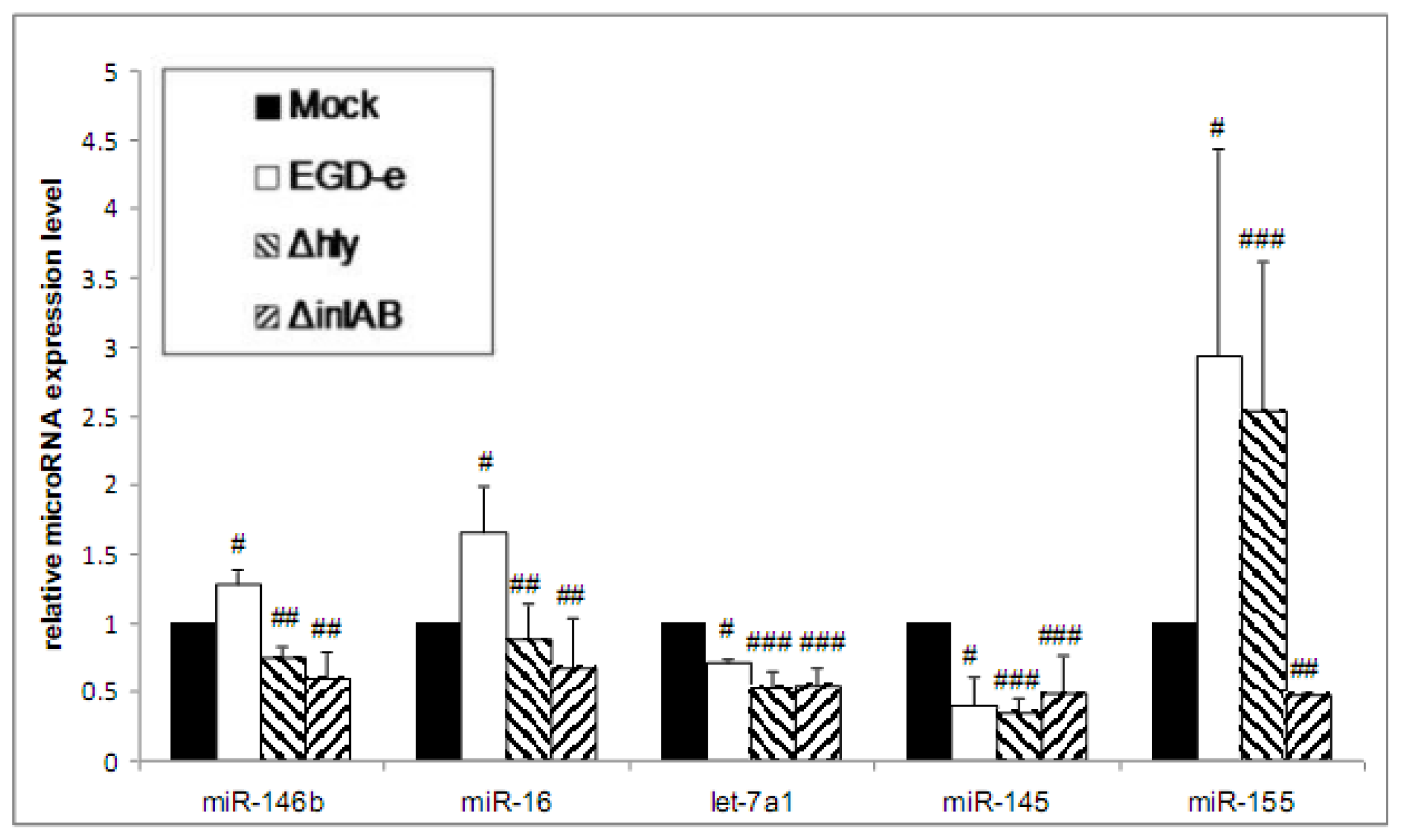
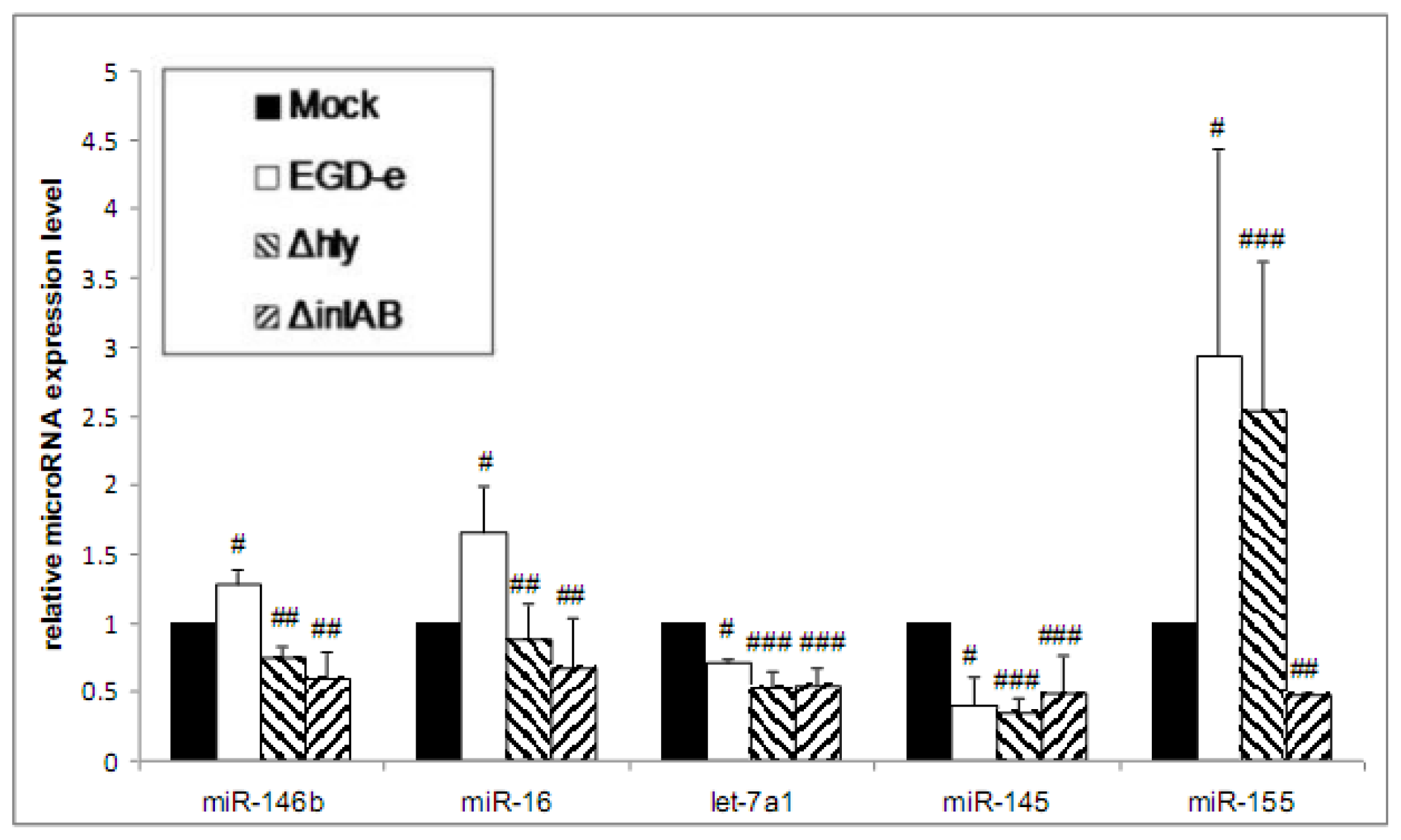
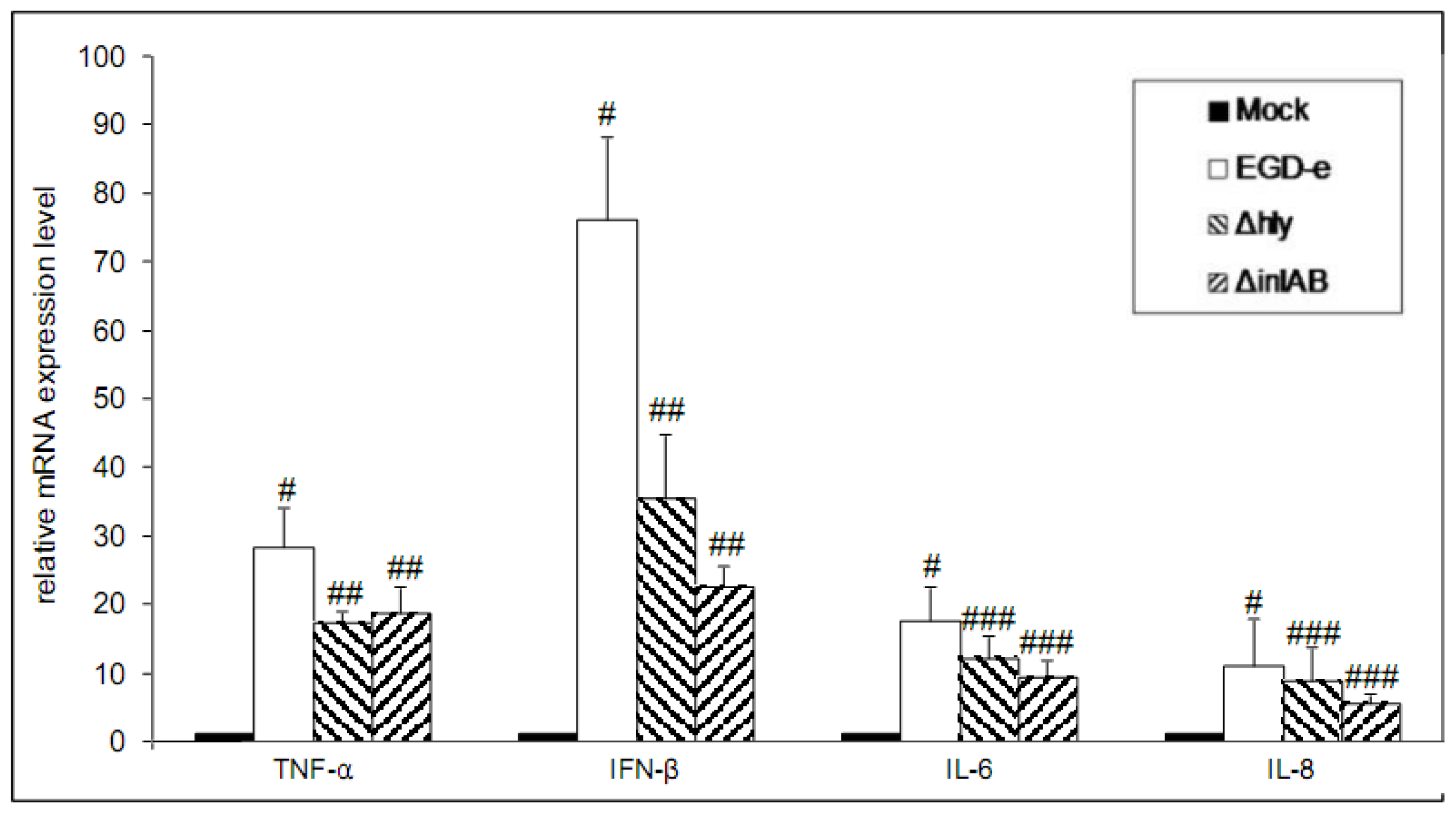
3.2. Wild-Type and LLO-Deficient Bacteria Induce miR-155, While the ΔinlAB Mutant Strain Suppresses miR-155 Expression
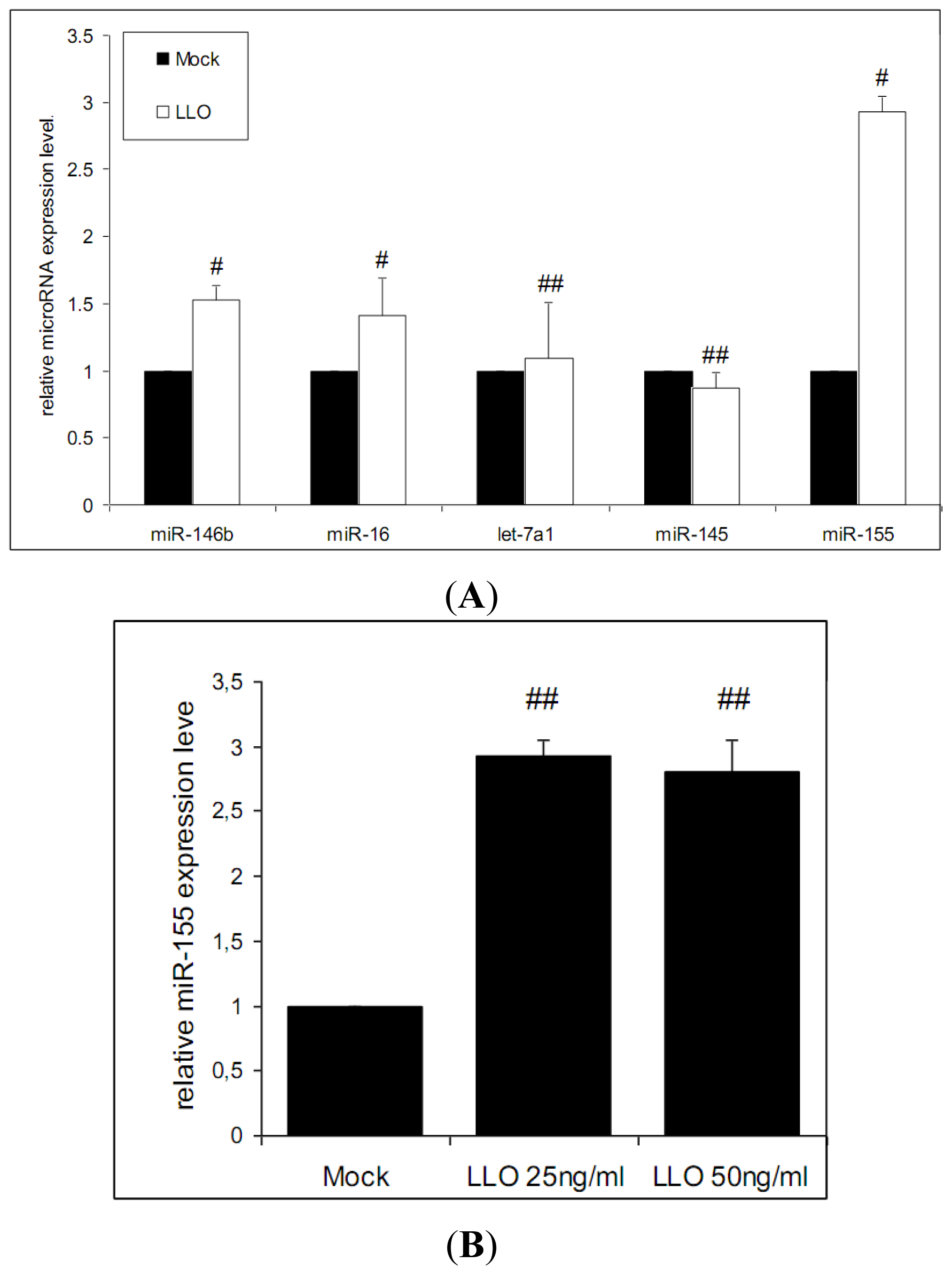
3.3. Purified LLO Induces the Expression of miR-146b, miR-16 and miR-155 in Caco-2 Cells
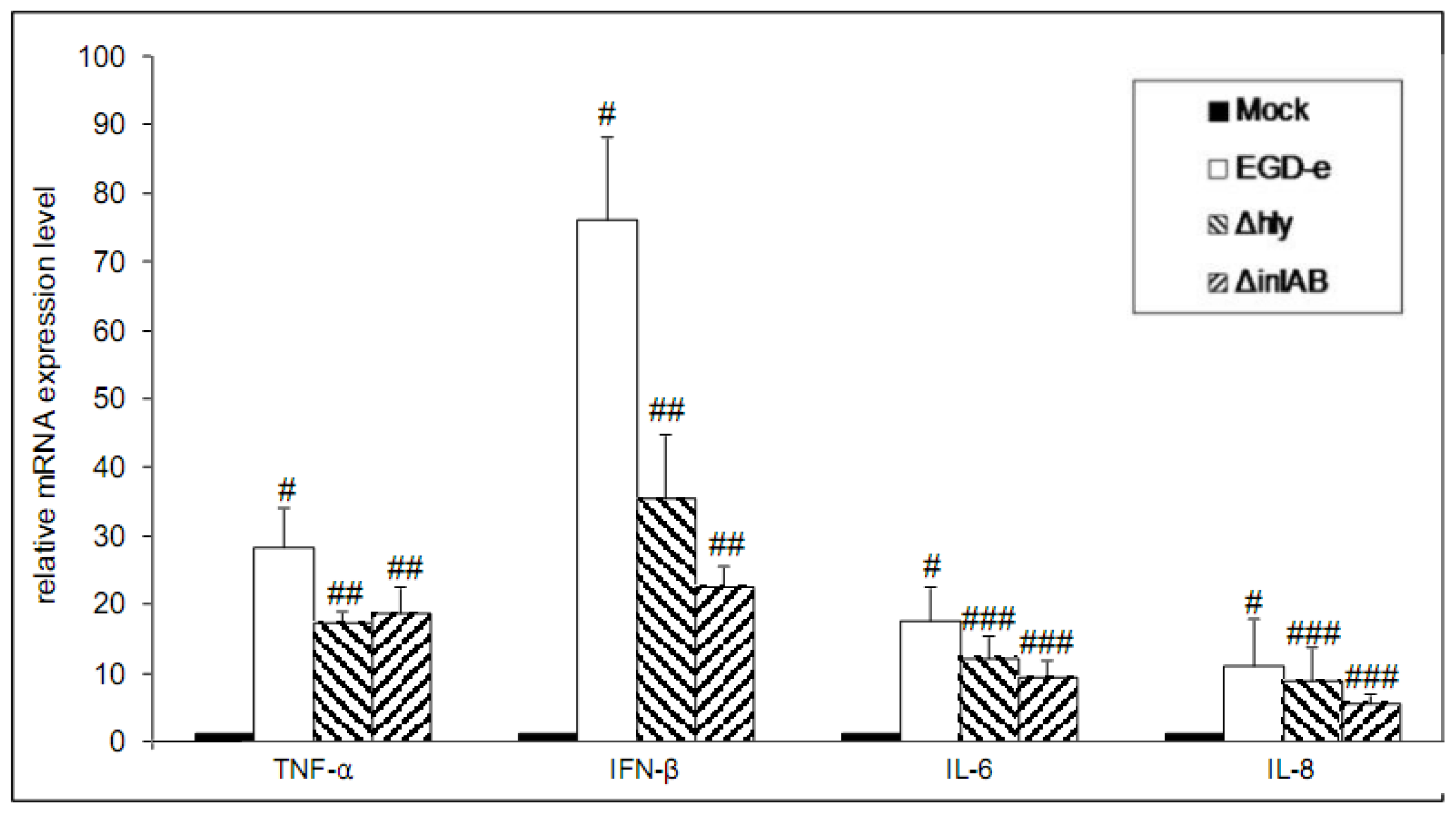
3.4. Deregulation of mRNAs That Are Targeted by miRNAs
4. Discussion
5. Conclusion
Supplementary Information
ijms-13-01173-s001.pdfAcknowledgements
- Conflict of InterestThere is no conflict of interest.
References
- Ambros, V. The functions of animal microRNAs. Nature 2004, 431, 350–355. [Google Scholar]
- Huang, Y.; Shen, X.J.; Zou, Q.; Wang, S.P.; Tang, S.M.; Zhang, G.Z. Biological functions of microRNAs: A review. J. Physiol. Biochem 2011, 67, 129–139. [Google Scholar]
- Cullen, B.R. Viruses and microRNAs: RISCy interactions with serious consequences. Genes Dev 2011, 25, 1881–1894. [Google Scholar]
- Oertli, M.; Engler, D.B.; Kohler, E.; Koch, M.; Meyer, T.F.; Muller, A. MicroRNA-155 is essential for the T cell-mediated control of Helicobacter pylori infection and for the induction of chronic Gastritis and Colitis. J. Immunol 2011, 187, 3578–3586. [Google Scholar]
- Schulte, L.N.; Eulalio, A.; Mollenkopf, H.J.; Reinhardt, R.; Vogel, J. Analysis of the host microRNA response to Salmonella uncovers the control of major cytokines by the let-7 family. EMBO J 2011, 30, 1977–1989. [Google Scholar]
- Russo, A.; Potenza, N. Antiviral effects of human microRNAs and conservation of their target sites. FEBS Lett 2011, 585, 2551–2555. [Google Scholar]
- Plaisance-Bonstaff, K.; Renne, R. Viral miRNAs. Methods Mol. Biol 2011, 721, 43–66. [Google Scholar]
- Navarro, L.; Jay, F.; Nomura, K.; He, S.Y.; Voinnet, O. Suppression of the microRNA pathway by bacterial effector proteins. Science 2008, 321, 964–967. [Google Scholar]
- Rao, J.R.; Nelson, D.; Moore, J.E.; Millar, B.C.; Goldsmith, C.E.; Rendall, J.; Elborn, J.S. Non-coding small (micro) RNAs of Pseudomonas aeruginosa isolated from clinical isolates from adult patients with cystic fibrosis. Br. J. Biomed. Sci 2010, 67, 126–132. [Google Scholar]
- Xiao, B.; Liu, Z.; Li, B.S.; Tang, B.; Li, W.; Guo, G.; Shi, Y.; Wang, F.; Wu, Y.; Tong, W.D.; et al. Induction of microRNA-155 during Helicobacter pylori infection and its negative regulatory role in the inflammatory response. J. Infect. Dis 2009, 200, 916–925. [Google Scholar]
- Glaser, P.; Frangeul, L.; Buchrieser, C.; Rusniok, C.; Amend, A.; Baquero, F.; Berche, P.; Bloecker, H.; Brandt, P.; Chakraborty, T.; et al. Comparative genomics of Listeria species. Science 2001, 294, 849–852. [Google Scholar]
- Guzman, C.A.; Rohde, M.; Chakraborty, T.; Domann, E.; Hudel, M.; Wehland, J.; Timmis, K.N. Interaction of Listeria monocytogenes with mouse dendritic cells. Infect. Immun 1995, 63, 3665–3673. [Google Scholar]
- Parida, S.K.; Domann, E.; Rohde, M.; Muller, S.; Darji, A.; Hain, T.; Wehland, J.; Chakraborty, T. Internalin B is essential for adhesion and mediates the invasion of Listeria monocytogenes into human endothelial cells. Mol. Microbiol 1998, 28, 81–93. [Google Scholar]
- Rose, F.; Zeller, S.A.; Chakraborty, T.; Domann, E.; Machleidt, T.; Kronke, M.; Seeger, W.; Grimminger, F.; Sibelius, U. Human endothelial cell activation and mediator release in response to Listeria monocytogenes virulence factors. Infect. Immun 2001, 69, 897–905. [Google Scholar]
- Collison, A.; Mattes, J.; Plank, M.; Foster, P.S. Inhibition of house dust mite-induced allergic airways disease by antagonism of microRNA-145 is comparable to glucocorticoid treatment. J. Allergy Clin. Immunol 2011, 128, 160–167. [Google Scholar]
- Bhaumik, D.; Scott, G.K.; Schokrpur, S.; Patil, C.K.; Orjalo, A.V.; Rodier, F.; Lithgow, G.J.; Campisi, J. MicroRNAs miR-146a/b negatively modulate the senescence-associated inflammatory mediators IL-6 and IL-8. Aging (Albany NY) 2009, 1, 402–411. [Google Scholar]
- Jing, Q.; Huang, S.; Guth, S.; Zarubin, T.; Motoyama, A.; Chen, J.; Di Padova, F.; Lin, S.C.; Gram, H.; Han, J. Involvement of microRNA in AU-rich element-mediated mRNA instability. Cell 2005, 120, 623–634. [Google Scholar]
- Witwer, K.W.; Sisk, J.M.; Gama, L.; Clements, J.E. MicroRNA regulation of IFN-beta protein expression: Rapid and sensitive modulation of the innate immune response. J. Immunol 2010, 184, 2369–2376. [Google Scholar]
- Tili, E.; Michaille, J.J.; Cimino, A.; Costinean, S.; Dumitru, C.D.; Adair, B.; Fabbri, M.; Alder, H.; Liu, C.G.; Calin, G.A.; et al. Modulation of miR-155 and miR-125b levels following lipopolysaccharide/TNF-alpha stimulation and their possible roles in regulating the response to endotoxin shock. J. Immunol 2007, 179, 5082–5089. [Google Scholar]
- O’Connell, R.M.; Taganov, K.D.; Boldin, M.P.; Cheng, G.; Baltimore, D. MicroRNA-155 is induced during the macrophage inflammatory response. Proc. Natl. Acad. Sci. USA 2007, 104, 1604–1609. [Google Scholar]
- Zhu, J.Y.; Strehle, M.; Frohn, A.; Kremmer, E.; Hofig, K.P.; Meister, G.; Adler, H. Identification and analysis of expression of novel microRNAs of murine gammaherpesvirus 68. J. Virol 2010, 84, 10266–10275. [Google Scholar]
- Zhou, R.; Hu, G.; Liu, J.; Gong, A.Y.; Drescher, K.M.; Chen, X.M. NF-kappaB p65-dependent transactivation of miRNA genes following Cryptosporidium parvum infection stimulates epithelial cell immune responses. PLoS Pathog 2009, 5, e1000681. [Google Scholar]
- Landgraf, P.; Rusu, M.; Sheridan, R.; Sewer, A.; Iovino, N.; Aravin, A.; Pfeffer, S.; Rice, A.; Kamphorst, A.O.; Landthaler, M.; et al. A mammalian microRNA expression atlas based on small RNA library sequencing. Cell 2007, 129, 1401–1414. [Google Scholar]
- Linsley, P.S.; Schelter, J.; Burchard, J.; Kibukawa, M.; Martin, M.M.; Bartz, S.R.; Johnson, J.M.; Cummins, J.M.; Raymond, C.K.; Dai, H.; et al. Transcripts targeted by the microRNA-16 family cooperatively regulate cell cycle progression. Mol. Cell Biol 2007, 27, 2240–2252. [Google Scholar]
- Taganov, K.D.; Boldin, M.P.; Chang, K.J.; Baltimore, D. NF-kappaB-dependent induction of microRNA miR-146, an inhibitor targeted to signaling proteins of innate immune responses. Proc. Natl. Acad. Sci. USA 2006, 103, 12481–12486. [Google Scholar]
- Quinn, S.R.; O’Neill, L.A. A trio of microRNAs that control Toll-like receptor signalling. Int. Immunol 2011, 23, 421–425. [Google Scholar]
- Liu, Z.; Xiao, B.; Tang, B.; Li, B.; Li, N.; Zhu, E.; Guo, G.; Gu, J.; Zhuang, Y.; Liu, X.; et al. Up-regulated microRNA-146a negatively modulate Helicobacter pylori-induced inflammatory response in human gastric epithelial cells. Microbes Infect 2010, 12, 854–863. [Google Scholar]
- Vora, P.; Youdim, A.; Thomas, L.S.; Fukata, M.; Tesfay, S.Y.; Lukasek, K.; Michelsen, K.S.; Wada, A.; Hirayama, T.; Arditi, M.; et al. Beta-defensin-2 expression is regulated by TLR signaling in intestinal epithelial cells. J. Immunol 2004, 173, 5398–5405. [Google Scholar]
- Leber, J.H.; Crimmins, G.T.; Raghavan, S.; Meyer-Morse, N.P.; Cox, J.S.; Portnoy, D.A. Distinct TLR- and NLR-mediated transcriptional responses to an intracellular pathogen. PLoS Pathog 2008, 4. [Google Scholar] [CrossRef]
- Pekow, J.R.; Dougherty, U.; Mustafi, R.; Zhu, H.; Kocherginsky, M.; Rubin, D.T.; Hanauer, S.B.; Hart, J.; Chang, E.B.; Fichera, A.; et al. miR-143 and miR-145 are downregulated in ulcerative colitis: Putative regulators of inflammation and protooncogenes. Inflamm. Bowel Dis 2012, 12, 94–100. [Google Scholar]
- Linden, S.K.; Bierne, H.; Sabet, C.; Png, C.W.; Florin, T.H.; McGuckin, M.A.; Cossart, P. Listeria monocytogenes internalins bind to the human intestinal mucin MUC2. Arch. Microbiol 2008, 190, 101–104. [Google Scholar]
- Sachdeva, M.; Mo, Y.Y. MicroRNA-145 suppresses cell invasion and metastasis by directly targeting mucin 1. Cancer Res 2010, 70, 378–387. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
| microRNA | FC microarray | FC qRT-PCR | target mRNA | Reference |
|---|---|---|---|---|
| miR-146b | 1.43 | 1.28 | IL-8, IL-6 | [16] |
| miR-16 | 0.64 | 1.65 | TNF-α, IL-6, IL-8 | [17] |
| let-7a1 | 0.63 | 0.72 | IL-10, IL-6 | [5] |
| miR-145 | 0.39 | 0.39 | IFN-β | [18] |
| miR-155 | 1.783 | 2.92 | TNF-α, IFN-β | [19,20] |
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Izar, B.; Mannala, G.K.; Mraheil, M.A.; Chakraborty, T.; Hain, T. microRNA Response to Listeria monocytogenes Infection in Epithelial Cells. Int. J. Mol. Sci. 2012, 13, 1173-1185. https://doi.org/10.3390/ijms13011173
Izar B, Mannala GK, Mraheil MA, Chakraborty T, Hain T. microRNA Response to Listeria monocytogenes Infection in Epithelial Cells. International Journal of Molecular Sciences. 2012; 13(1):1173-1185. https://doi.org/10.3390/ijms13011173
Chicago/Turabian StyleIzar, Benjamin, Gopala Krishna Mannala, Mobarak Abu Mraheil, Trinad Chakraborty, and Torsten Hain. 2012. "microRNA Response to Listeria monocytogenes Infection in Epithelial Cells" International Journal of Molecular Sciences 13, no. 1: 1173-1185. https://doi.org/10.3390/ijms13011173