Difference in TRI13 Gene Sequences between the 3-Acetyldeoxynivalenol Producing Fusarium graminearum Chemotypes from Canada and China




Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Isolation and DNA Extraction
2.2. SCAR Analysis
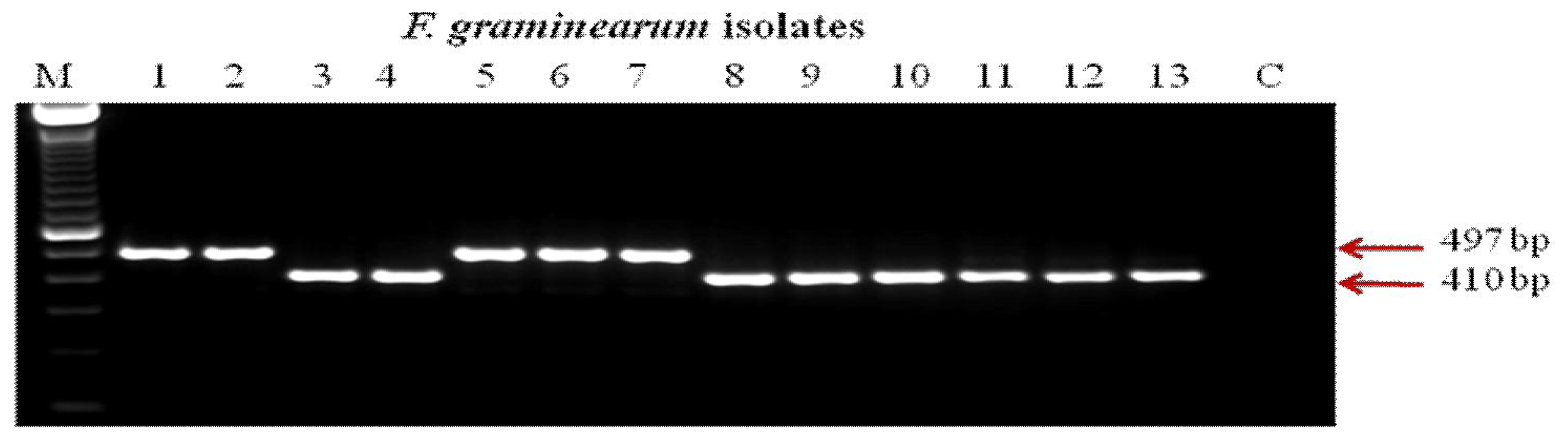
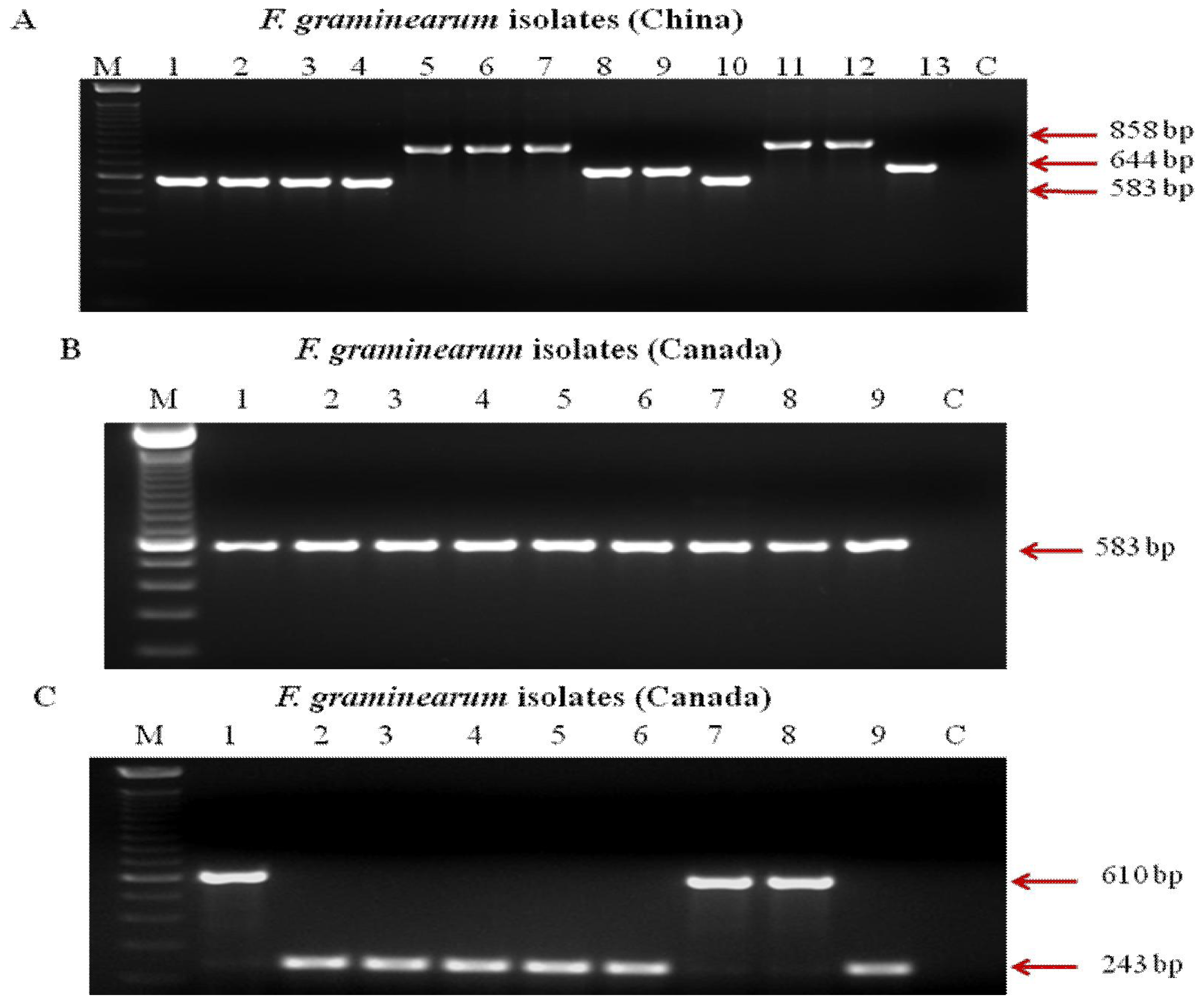
2.3. PCR Assays for Trichothecene Chemotypes
2.4. PCR Conditions
2.5. Confirmation of PCR Products Using Sequencing
3. Results
4. Discussion
Acknowledgements
References
- Charmley, LL; Trenholm, HL; Prelusky, DB; Rosenberg, A. Economic losses and decontamination. Nat Toxins 1995, 3, 199–203. [Google Scholar]
- Proctor, RH; Hohn, TM; McCormick, SP. Reduced virulence of Gibberella zeae caused by disruption of a trichothecene toxin biosynthetic gene. Mol Plant Microbe Interact 1995, 8, 593–601. [Google Scholar]
- Miller, JD; Greenhalgh, R; Wang, YZ; Lu, M. Trichothecene chemotypes of three Fusarium species. Mycologia 1991, 83, 121–130. [Google Scholar]
- Desjardins, AE. Trichothecenes. In Fusarium Mycotoxins-Chemistry, Genetics, and Biology; The American Phytopathological Society Press: St. Paul, MN, USA, 2006; pp. 13–64. [Google Scholar]
- Lee, T; Han, YK; Kim, KH; Yun, SH; Lee, YW. Tri13 and Tri7 determine deoxynivalenoland nivalenol-producing chemotypes of Gibberella zeae. Appl Environ Microbiol 2002, 68, 2148–2154. [Google Scholar]
- Lee, T; Oh, DW; Kim, HS; Lee, J; Kim, YH; Yun, SH; Lee, YW. Identification of deoxynivalenol- and nivalenol- producing chemotypes of Gibberella zeae by using PCR. Appl Environ Microbiol 2001, 67, 2966–2972. [Google Scholar]
- Ward, T; Bielawski, JP; Kistler, CH; Sullivan, E; O’Donnell, K. Ancestral polymorphism and adaptive evolution in the trichothecene mycotoxin gene cluster of phytopathogenic Fusarium. Proc Natl Acad Sci USA 2002, 99, 9278–9283. [Google Scholar]
- Jennings, P; Coates, ME; Turner, JA; Chandler, EA; Nicholson, P. Determination of deoxynivalenol and nivalenol chemotypes of Fusarium graminearum isolates from England and Wales by PCR assay. Plant Pathol 2004, 53, 643–652. [Google Scholar]
- Quarta, A; Mita, G; Haidukowski, M; Santino, A; Mule, G; Visconti, A. Multiplex PCR assay for the identification of nivalenol, 3- and 15-acetyldeoxynivalenol chemotypes in Fusarium. FEMS Microbiol Lett 2006, 259, 7–13. [Google Scholar]
- O’Donnell, K; Kistler, HC; Tacke, BK; Casper, HH. Gene genealogies reveal global phylogeographic structure and reproductive isolation among lineages of Fusarium graminearum, the fungus causing wheat scab. Proc Natl Acad Sci USA 2000, 97, 7905–7910. [Google Scholar]
- O’Donnell, K; Ward, TJ; Geiser, DM; Kistler, HC; Aoki, T. Genealogical concordance between the mating type locus and seven other nuclear genes supports formal recognition of nine phylogenetically distinct species within the Fusarium graminearum clade. Fungal Genet Biol 2004, 41, 600–623. [Google Scholar]
- Starkey, DE; Ward, TJ; Aoki, T; Gale, LR; Kistler, HC; Geiser, DM; Suga, H; Tóth, B; Varga, J; O’Donnell, K. Global molecular surveillance reveals novel Fusarium head blight species and trichothecene toxin diversity. Fungal Genet Biol 2007, 44, 1191–1204. [Google Scholar]
- O’Donnell, K; Ward, TJ; Aberra, D; Kistler, HC; Aoki, T; Orwig, N; Kimura, M; Bj⊘rnstad, A; Klemsdal, SS. Multilocus genotyping and molecular phylogenetics resolve a novel head blight pathogen within the Fusarium graminearum species complex from Ethiopia. Fungal Genet Biol 2008, 45, 1514–1522. [Google Scholar]
- Yli-Mattila, T; Gagkaeva, T; Ward, TJ; Aoki, T; Kistler, HC; O’Donnell, K. A novel Asian clade within the Fusarium graminearum species complex includes a newly discovered cereal head blight pathogen from the Russian Far East. Mycologia 2009, 101, 841–852. [Google Scholar]
- Goswami, RS; Kistler, HC. Pathogenicity and in planta mycotoxin accumulation among members of the Fusarium graminearum species complex on wheat and rice. Phytopathology 2005, 95, 1397–1404. [Google Scholar]
- Zhang, JB; Li, HP; Dang, FJ; Qu, B; Xu, YB; Zhao, CS; Liao, YC. Determination of the trichothecene mycotoxin chemotypes and associated geographical distribution and phylogenetic species of the Fusarium graminearum clade from China. Mycol Res 2007, 111, 967–975. [Google Scholar]
- Cumagun, CJR; Bowden, RL; Jurgenson, JE; Leslie, JF; Miedaner, T. Genetic mapping of pathogenicity and aggressiveness of Gibberella zeae (Fusarium graminearum) toward wheat. Phytopathology 2004, 94, 520–526. [Google Scholar]
- Qu, B; Li, HP; Zhang, JB; Xu, YB; Huang, T; Wu, AB; Zhao, CS; Carter, J; Nicholson, P; Liao, YC. Geographic distribution and genetic diversity of Fusarium graminearum and F. asiaticum on wheat spikes throughout China. Plant Pathol 2008, 57, 15–24. [Google Scholar]
- Bai, GH; Shaner, G. Variation in Fusarium graminearum and cultivar resistance to wheat scab. Plant Dis 1996, 80, 975–979. [Google Scholar]
- Brasier, CM; Cooke, DEL; Duncan, JM. Origin of a new Phytophthora pathogen through interspecific hybridization. Proc Natl Acad Sci USA 1999, 96, 5878–5883. [Google Scholar]
- Wang, J; Li, H; Qu, B; Zhang, J; Huang, T; Chen, F; Liao, Y. Development of a generic PCR detection of 3-acetyldeoxynivalenol, 15-acetyldeoxynivalenol- and nivalenol-chemotypes of Fusarium graminearum Clade. Int J Mol Sci 2008, 9, 2495–2504. [Google Scholar]
- Guo, XW; Fernando, WGD; Seow-Brock, HY. Population structure, chemotype diversity, and potential chemotype shifting of Fusarium graminearum in wheat fields of Manitoba. Plant Dis 2008, 92, 756–762. [Google Scholar]
- Fernando, WGD; Zhang, JK; Dusabenyagasani, M; Guo, X; Ahmed, H; McCallum, B. Genetic diversity of Gibberella zea isolates from Manitoba. Plant Dis 2006, 90, 1337–1342. [Google Scholar]
- Carter, JP; Razanoor, HN; Desjardins, AE; Nicholson, P. Variation in Fusarium graminearum isolates from Nepal associated with their host of origin. Plant Pathol 2000, 49, 1–10. [Google Scholar]
- Ward, TJ; Clear, RM; Rooney, AP; O’Donnell, K; Gaba, D; Patrick, S; Starkey, D; Gilbert, J; Geiser, D; Nowicki, T. An adaptive evolutionary shift in Fusarium head blight pathogen populations is driving the rapid spread of more toxigenic Fusarium graminearum in North America. Fungal Genet Biol 2008, 25, 473–484. [Google Scholar]
- Li, HP; Wu, AB; Zhao, CS; Scholten, O; Löffler, H; Liao, YC. Development of a generic PCR detection of deoxynivalenol- and nivalenol-chemotypes of Fusarium graminearum. FEMS Microbiol Lett 2005, 243, 505–511. [Google Scholar]
- Brown, DW; McCormick, SP; Alexander, NJ; Proctor, RH; Desjardins, AE. Inactivation of a cytochrome P-450 is a determinant of trichothecene diversity in Fusarium species. Fungal Genet Biol 2002, 36, 224–233. [Google Scholar]
- Brown, DW; McCormick, SP; Alexander, NJ; Proctor, RH; Desjardins, AE. A genetic and biochemical approach to study trichothecene diversity in Fusarium sporotrichioides and Fusarium graminearum. Fungal Genet Biol 2001, 32, 121–133. [Google Scholar]
- Chandler, EA; Simpson, DR; Thomsett, MA; Nicholson, P. Development of PCR assays to Tri7 and Tri13 trichothecene biosynthetic genes, and characterisation of chemotypes of Fusarium graminearum, Fusarium culmorum and Fusarium cerealis. Physiol Mol Plant Pathol 2003, 62, 355–367. [Google Scholar]
- McCormick, SP; Hohn, TM; Desjardins, AE. Isolation and characterization of Tri3, a gene encoding 15-O-acetyltransferase from Fusarium sporotrichioides. Appl Environ Microbiol 1996, 62, 353–359. [Google Scholar]
- Mirocha, CJ; Abbas, HK; Windels, CE; Xie, W. Variation in deoxynivalenol, 15-acetyldeoxynivalenol, 3-acetyldeoxynivalenol, and zearalenone production by Fusarium graminearum isolates. Appl Environ Microbiol 1989, 55, 1315–1316. [Google Scholar]
- Gale, LR; Harrison, SA; Ward, TJ; O’Donnell, K; Milus, EA; Gale, SW; Kistler, HC. Nivalenol-type populations of Fusarium graminearum and F. asiaticum are prevalent on wheat in southern Louisiana. Phytopathology 2010, 101, 124–134. [Google Scholar]
- Bowden, RL; Leslie, JF. Sexual recombination in Gibberella zeae. Phytopathology 1999, 89, 182–188. [Google Scholar]
- Bowden, RL; Leslie, JF; Lee, J; Lee, YW. Cross Fertility of Lineages in Fusarium graminearum (Gibberella zeae. In The Global Fusarium Initiative for International Collaboration: A Strategic Planning Workshop; Ban, T, Lewis, JM, Phipps, EE, Eds.; CIMMYT: El Batan, Mexico, 2006; pp. 54–60. [Google Scholar]

 - represent insertions in the TRI13 gene.
- represent insertions in the TRI13 gene.
- represent insertions in the TRI13 gene.
- represent insertions in the TRI13 gene.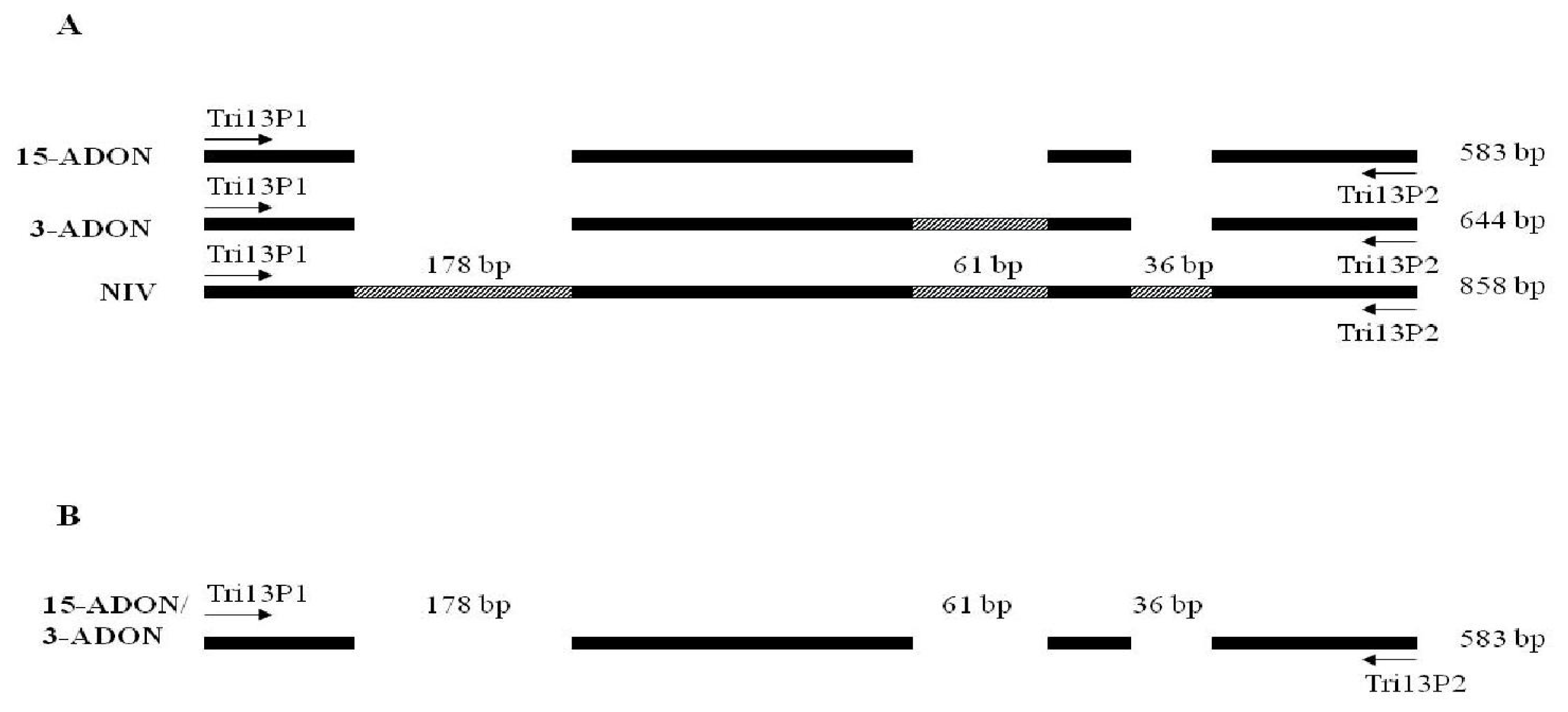

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location | Host | Number of Isolates |
|---|---|---|
| Manitoba, Canada | Wheat | 93 |
| Saskatchewan, Canada | Wheat | 15 |
| Nova Scotia, Canada | Wheat | 3 |
| Alberta, Canada | Wheat | 4 |
| Prince Edward Island, Canada | Wheat | 2 |
| Quebec, Canada | Wheat | 3 |
| China | Wheat | 39 |
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Amarasinghe, C.; Wang, J.-H.; Liao, Y.-C.; Fernando, W.G.D. Difference in TRI13 Gene Sequences between the 3-Acetyldeoxynivalenol Producing Fusarium graminearum Chemotypes from Canada and China. Int. J. Mol. Sci. 2011, 12, 6164-6175. https://doi.org/10.3390/ijms12096164
Amarasinghe C, Wang J-H, Liao Y-C, Fernando WGD. Difference in TRI13 Gene Sequences between the 3-Acetyldeoxynivalenol Producing Fusarium graminearum Chemotypes from Canada and China. International Journal of Molecular Sciences. 2011; 12(9):6164-6175. https://doi.org/10.3390/ijms12096164
Chicago/Turabian StyleAmarasinghe, Chami, Jian-Hua Wang, Yu-Cai Liao, and W.G. Dilantha Fernando. 2011. "Difference in TRI13 Gene Sequences between the 3-Acetyldeoxynivalenol Producing Fusarium graminearum Chemotypes from Canada and China" International Journal of Molecular Sciences 12, no. 9: 6164-6175. https://doi.org/10.3390/ijms12096164