Silibinin Causes Apoptosis and Cell Cycle Arrest in Some Human Pancreatic Cancer Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
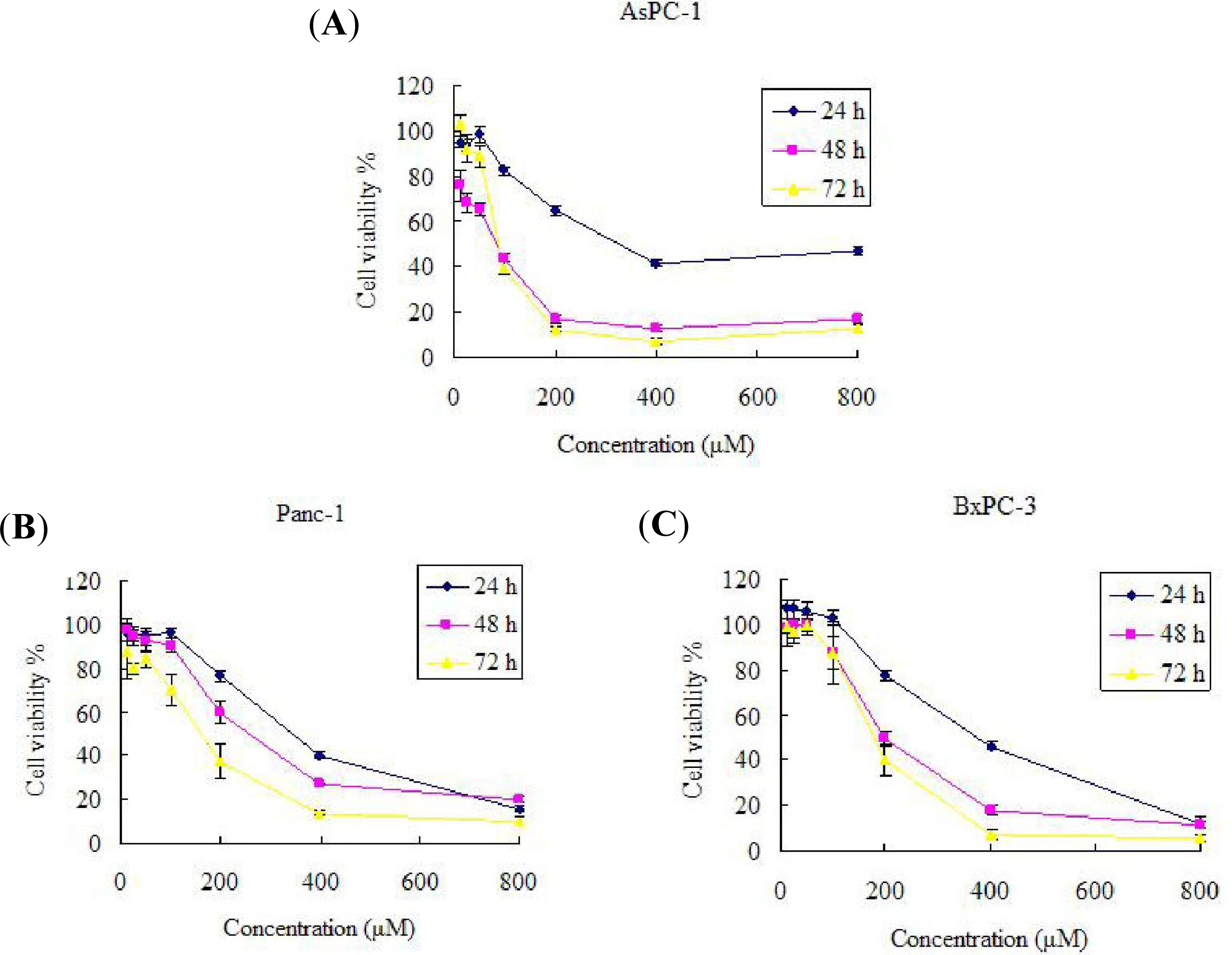
2.1. Effect of Silibinin on AsPC-1, Panc-1 and BxPC-3 Cell Growth Inhibition
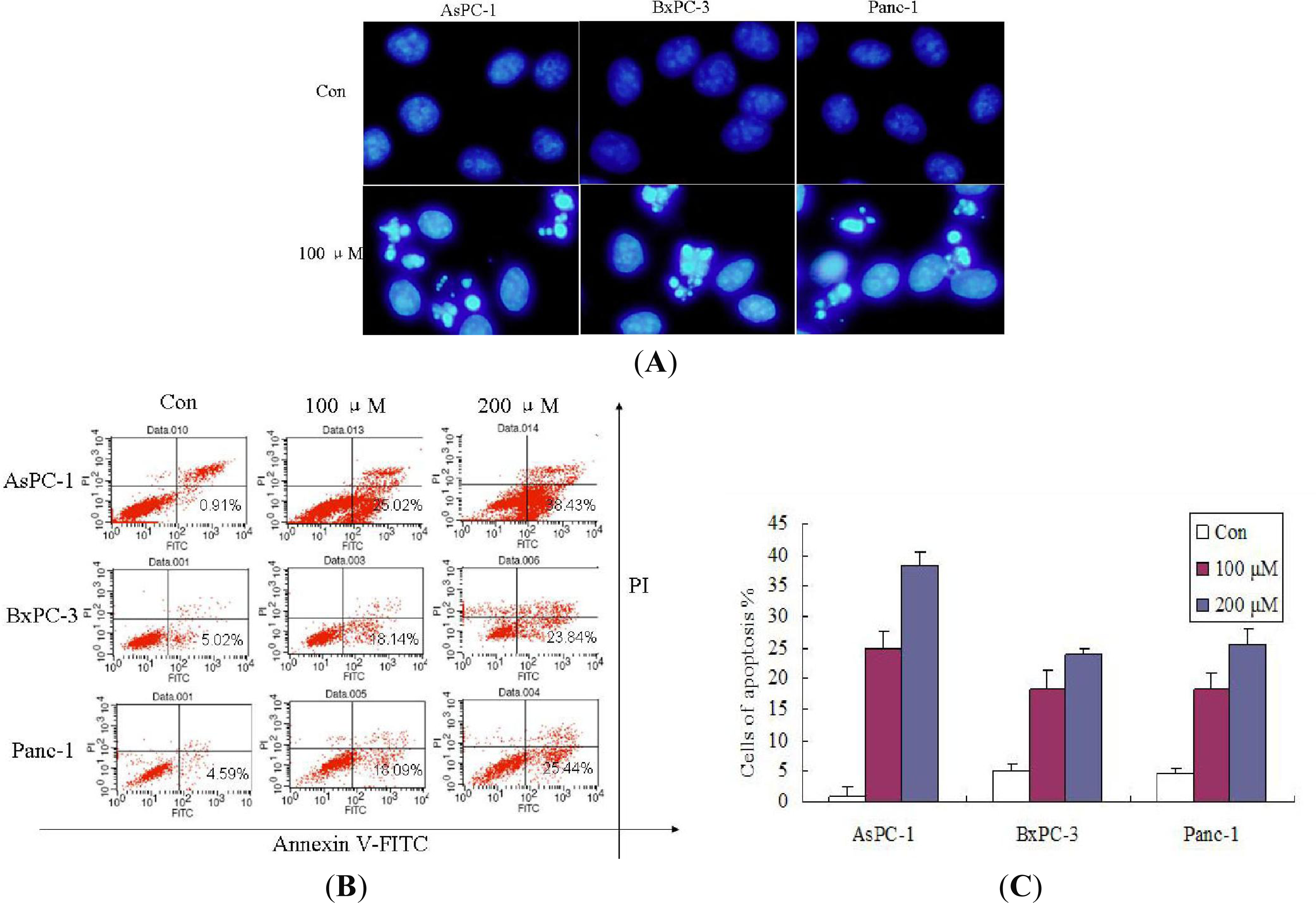
2.2. Effect of Silibinin on the Induction of Apoptosis
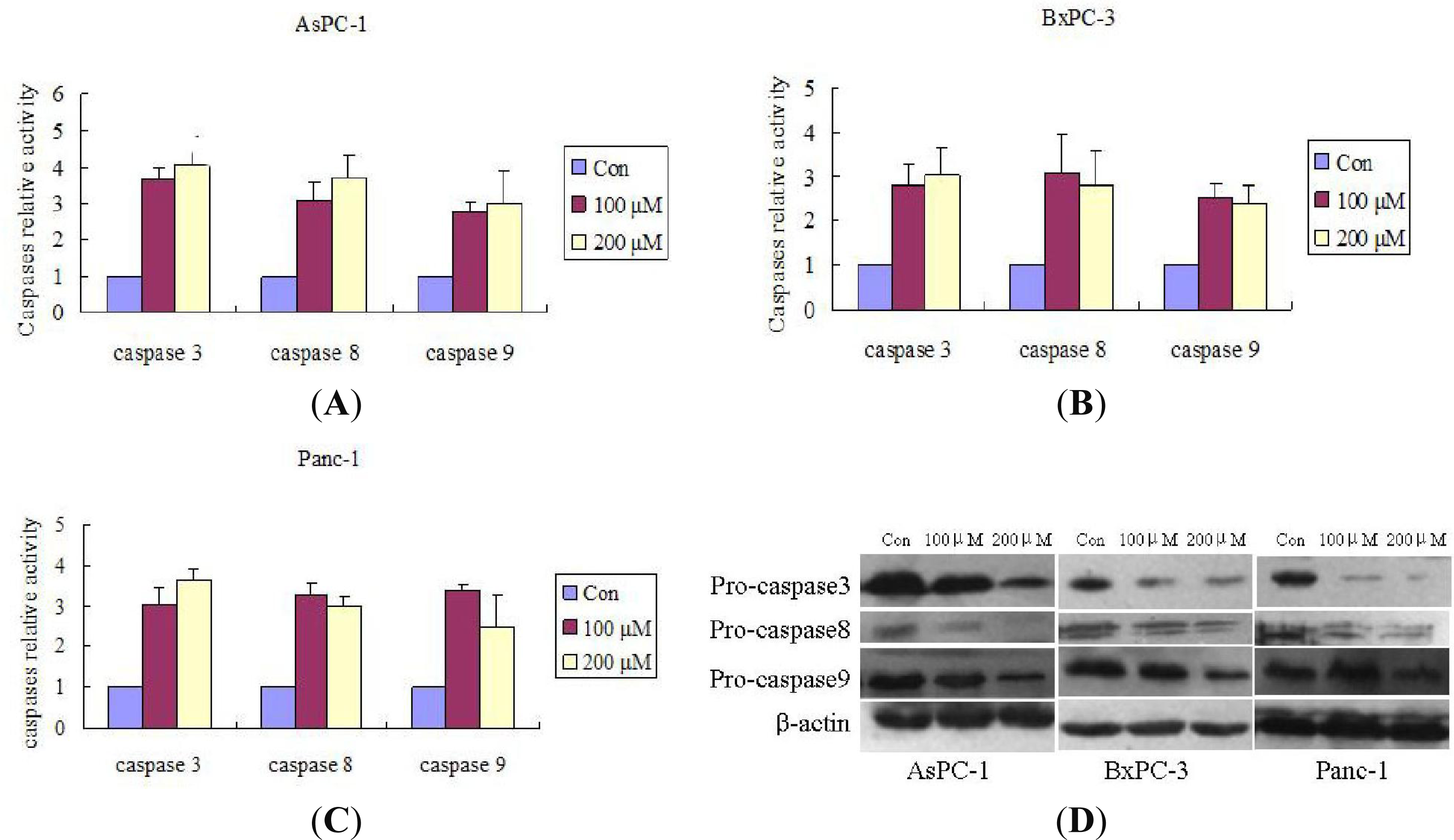
2.3. Silibinin Triggers Apoptosis by Activating the Caspases
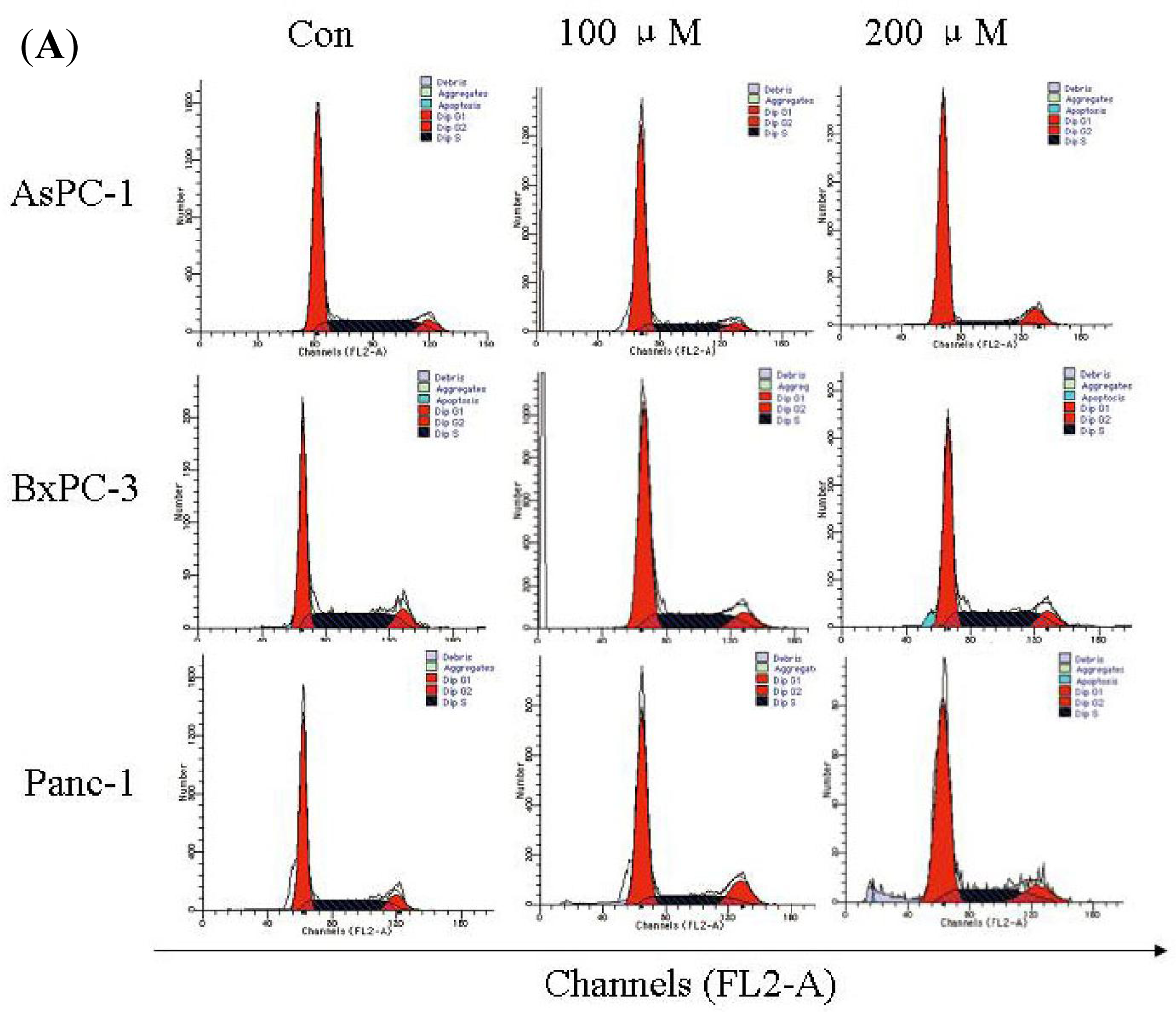
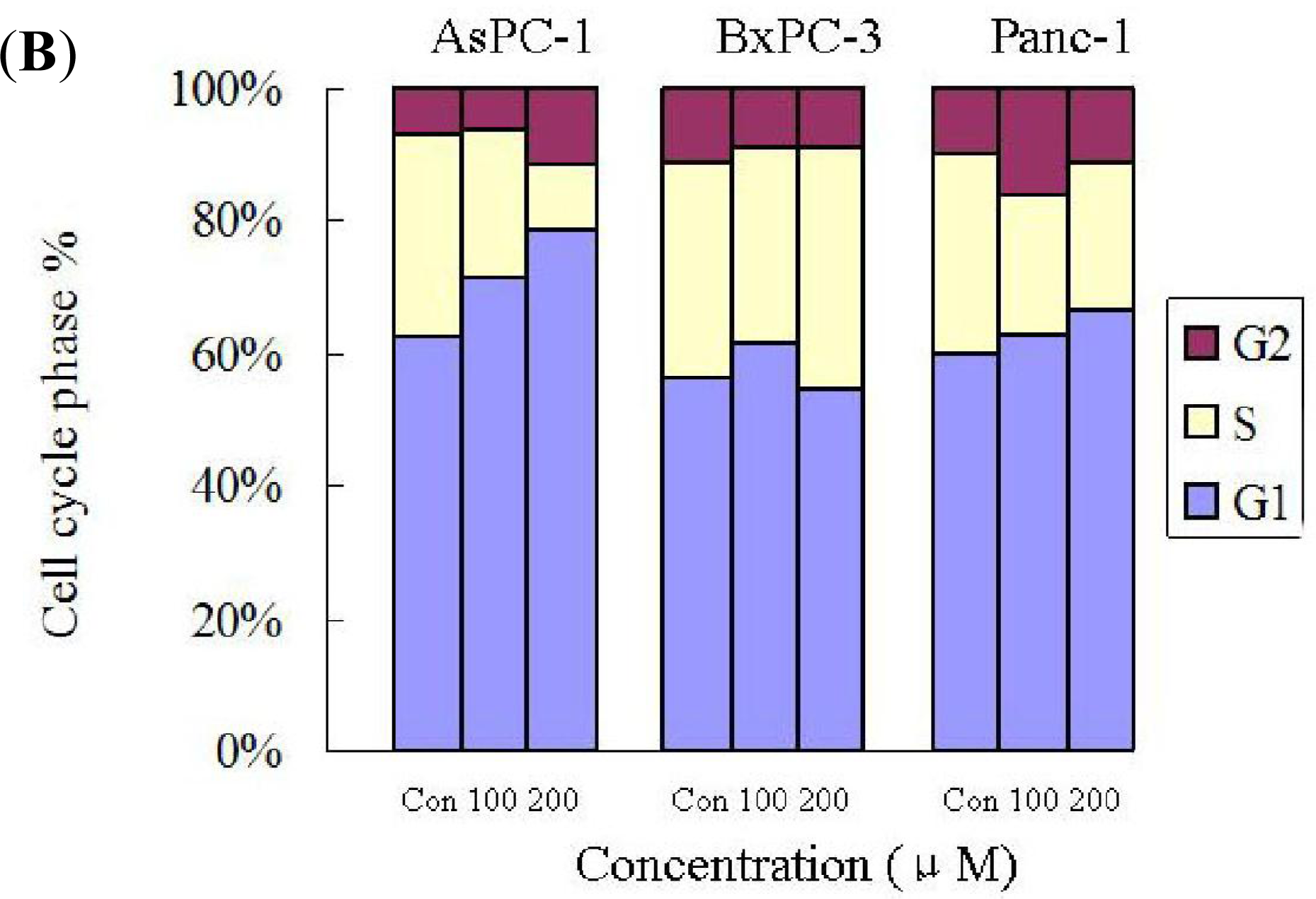
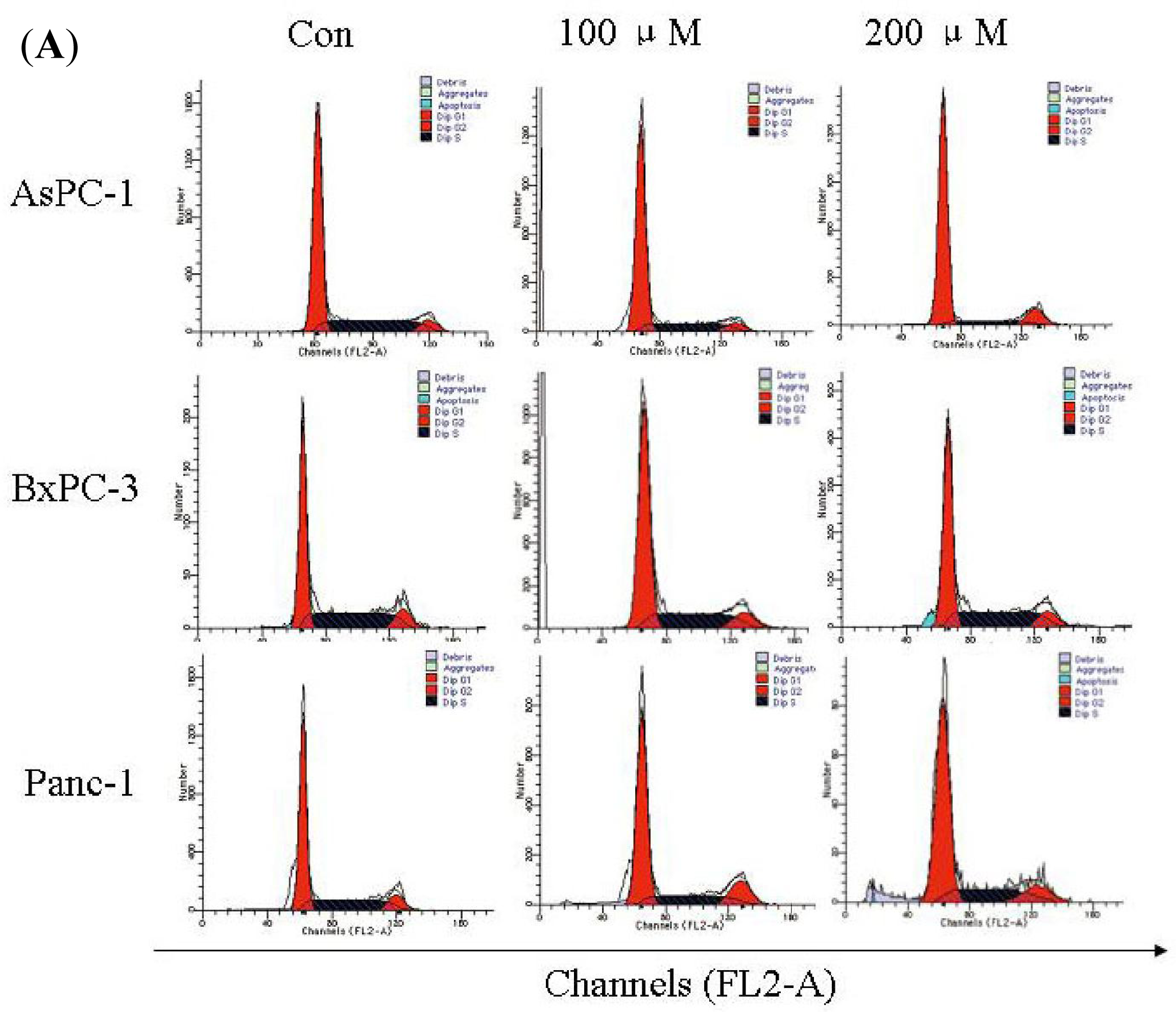
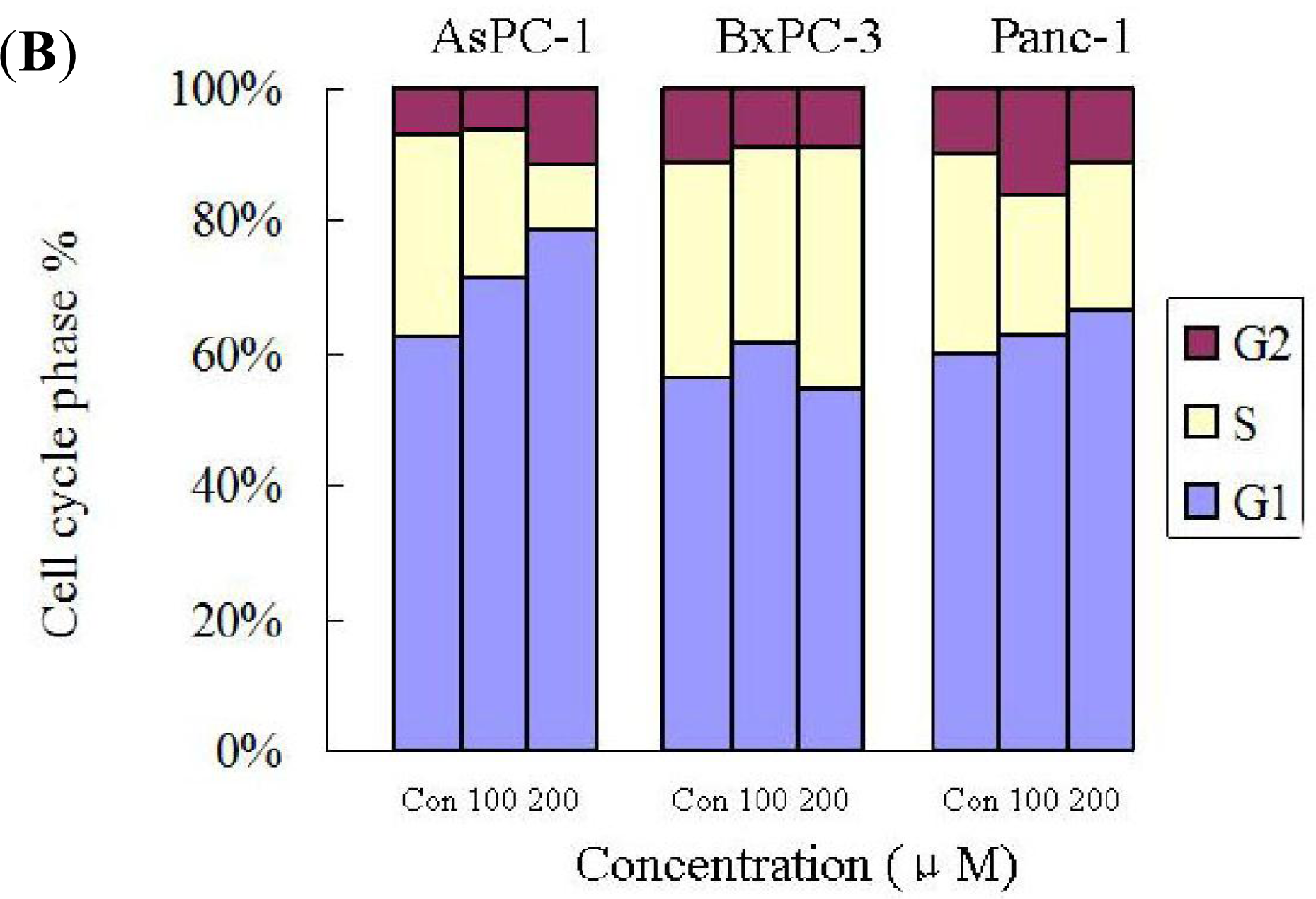
2.4. Silibinin Induces Cell-Cycle Arrest in Human AsPC-1, but not in Panc-1 and BxPC-3 Cells
3. Experimental Section
3.1. Cell Line and Reagents
3.2. Cell Proliferation Assay
3.3. Nuclear Staining with DAPI
3.4. Flow Cytometric Analysis: Cell Cycle and Apoptosis
3.5. Caspase Activity Assay
3.6. Western Blotting Analysis
3.7. Statistical Analysis
4. Conclusions
Acknowledgments
References
- Duan, WJ; Li, QS; Xia, MY; Tashiro, S; Onodera, S; Ikejima, T. Silibinin activated p53 and induced autophagic death in human fibrosarcoma HT1080 cells via reactive oxygen species-p38 and c-Jun N-terminal kinase pathways. Biol. Pharm. Bull 2011, 34, 47–53. [Google Scholar]
- Deep, G; Agarwal, R. Antimetastatic efficacy of Silibinin: Molecular mechanisms and therapeutic potential against cancer. Cancer Metast. Rev 2010, 29, 447–463. [Google Scholar]
- Hahn, VG; Lehmann, HD; Kürten, M; Uebel, H; Vogel, G. Pharmacology and toxicology of silymarin antihepatotoxic agent of Silybum marianum (L) gaertn. Arzneim.-Forsch.-Drug Res 1968, 18, 698–704. [Google Scholar]
- Vogel, G; Trost, W; Braatz, R; Odenthal, KP; Brüsewitz, G; Antweiler, H; Seeger, R. Pharmacodynamics, site and mechanism of action of silymarin, the antihepatoxic principle from Silybum mar. (L) Gaertn. 1. Acute toxicology or tolerance, general and specific (liver-) pharmacology. Arzneim.-Forsch.-Drug Res 1975, 25, 82–89. [Google Scholar]
- Flora, K; Hahn, M; Rosen, H; Benner, K. Milk thistle (Silybum marianum) for the therapy of liver disease. Am. J. Gastroenterol 1998, 93, 139–143. [Google Scholar]
- Wellington, K; Jarvis, B. Silymarin: A review of its clinical properties in the management of hepatic disorders. Biodrugs 2001, 15, 465–489. [Google Scholar]
- Kim, HO; Choi, HK; Ha, ES; Lee, KJ; Hur, GY; Jung, KH; Lee, SY; Lee, SY; Kim, JH; Shin, C; et al. Silibinin induces apoptosis by downregulating survivin through inhibition of hypoxia-inducible factor-1 alpha in non-small cell lung cancer cells. J. Thorac. Oncol 2010, 5, S248. [Google Scholar]
- Tyagi, A; Singh, RP; Agarwal, C; Agarwal, R. Silibinin activates p53-caspase 2 pathway and causes caspase-mediated cleavage of Cip1/p21 in apoptosis induction in bladder transitional-cell papilloma RT4 cells: Evidence for a regulatory loop between p53 and caspase 2. Carcinogenesis 2006, 27, 2269–2280. [Google Scholar]
- Agarwal, C; Singh, RP; Dhanalakshmi, S; Tyagi, AK; Tecklenburg, M; Sclafani, RA; Agarwal, R. Silibinin upregulates the expression of cyclin-dependent kinase inhibitors and causes cell cycle arrest and apoptosis in human colon carcinoma HT-29 cells. Oncogene 2003, 22, 8271–8282. [Google Scholar]
- Jemal, A; Siegel, R; Xu, JQ; Ward, E. Cancer Statistics, 2010. CA Cancer J. Clin 2010, 60, 277–300. [Google Scholar]
- Park, YJ; Wen, J; Bang, S; Park, SW; Song, SY. [6]-Gingerol induces cell cycle arrest and cell death of mutant p53-expressing pancreatic cancer cells. Yonsei Med. J 2006, 47, 688–697. [Google Scholar]
- Lowe, SW; Lin, AW. Apoptosis in cancer. Carcinogenesis 2000, 21, 485–495. [Google Scholar]
- Petak, I; Houghton, JA. Shared pathways: Death receptors and cytotoxic drugs in cancer therapy. Pathol. Oncol. Res 2001, 7, 95–106. [Google Scholar]
- Debatin, KM; Poncet, D; Kroemer, G. Chemotherapy: Targeting the mitochondrial cell death pathway. Oncogene 2002, 21, 8786–8803. [Google Scholar]
- Irene, M; Thomas, E; Alex, A. Targeting apoptosis Pathways in Cancer Therapy. CA Cancer J. Clin 2005, 55, 178–194. [Google Scholar]
- Tyagi, A; Agarwal, C; Harrison, G; Glode, LM; Agarwal, R. Silibinin causes cell cycle arrest and apoptosis in human bladder transitional cell carcinoma cells by regulating CDKI-CDK-cyclin cascade, and caspase 3 and PARP cleavages. Carcinogenesis 2004, 25, 1711–1720. [Google Scholar]
- Hogan, FS; Krishnegowda, NK; Mikhailova, M; Kahlenberg, MS. Flavonoid, silibinin, inhibits proliferation and promotes cell-cycle arrest of human colon cancer. J. Surg. Res 2007, 143, 58–65. [Google Scholar]
- Deep, G; Singh, RP; Agarwal, C; Kroll, DJ; Agarwal, R. Silymarin and silibinin cause G1 and G2-M cell cycle arrest via distinct circuitries in human prostate cancer PC3 cells: A comparison of flavanone silibinin with flavanolignan mixture silymarin. Oncogene 2006, 25, 1053–1069. [Google Scholar]
- Mateen, S; Tyagi, A; 1, Agarwal, C; Singh, RP; Agarwal, R. Silibinin inhibits human nonsmall cell lung cancer cell growth through cell-cycle arrest by modulating expression and function of key cell-cycle regulators. Mol. Carcinog 2010, 49, 247–258. [Google Scholar]
- Agarwal, C; Singh, RP; Dhanalakshmi, S; Tyagi, AK; Tecklenburg, M; Sclafani, RA; Agarwal, R. Silibinin upregulates the expression of cyclin-dependent kinase inhibitors and causes cell cycle arrest and apoptosis in human colon carcinoma HT-29 cells. Oncogene 2003, 22, 8271–8282. [Google Scholar]
- Li, L; Gao, Y; Zhang, L; Zeng, J; He, D; Sun, Y. Silibinin inhibits cell growth and induces apoptosis by caspase activation, down-regulating survivin and blocking EGFR-ERK activation in renal cell carcinoma. Cancer Lett 2008, 272, 61–69. [Google Scholar]
- Wesierska-Gadek, J; Gueorguieva, M; Horky, M. Dual action of cyclin-dependent kinase inhibitors: induction of cell cycle arrest and apoptosis. A comparison of the effects exerted by roscovitine and cisplatin. Pol. J. Pharmacol 2003, 55, 895–902. [Google Scholar]

© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Ge, Y.; Zhang, Y.; Chen, Y.; Li, Q.; Chen, J.; Dong, Y.; Shi, W. Silibinin Causes Apoptosis and Cell Cycle Arrest in Some Human Pancreatic Cancer Cells. Int. J. Mol. Sci. 2011, 12, 4861-4871. https://doi.org/10.3390/ijms12084861
Ge Y, Zhang Y, Chen Y, Li Q, Chen J, Dong Y, Shi W. Silibinin Causes Apoptosis and Cell Cycle Arrest in Some Human Pancreatic Cancer Cells. International Journal of Molecular Sciences. 2011; 12(8):4861-4871. https://doi.org/10.3390/ijms12084861
Chicago/Turabian StyleGe, Yakun, Yuanxin Zhang, Yunpeng Chen, Quanshun Li, Jun Chen, Ying Dong, and Wei Shi. 2011. "Silibinin Causes Apoptosis and Cell Cycle Arrest in Some Human Pancreatic Cancer Cells" International Journal of Molecular Sciences 12, no. 8: 4861-4871. https://doi.org/10.3390/ijms12084861