Serine Proteases-Like Genes in the Asian Rice Gall Midge Show Differential Expression in Compatible and Incompatible Interactions with Rice


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
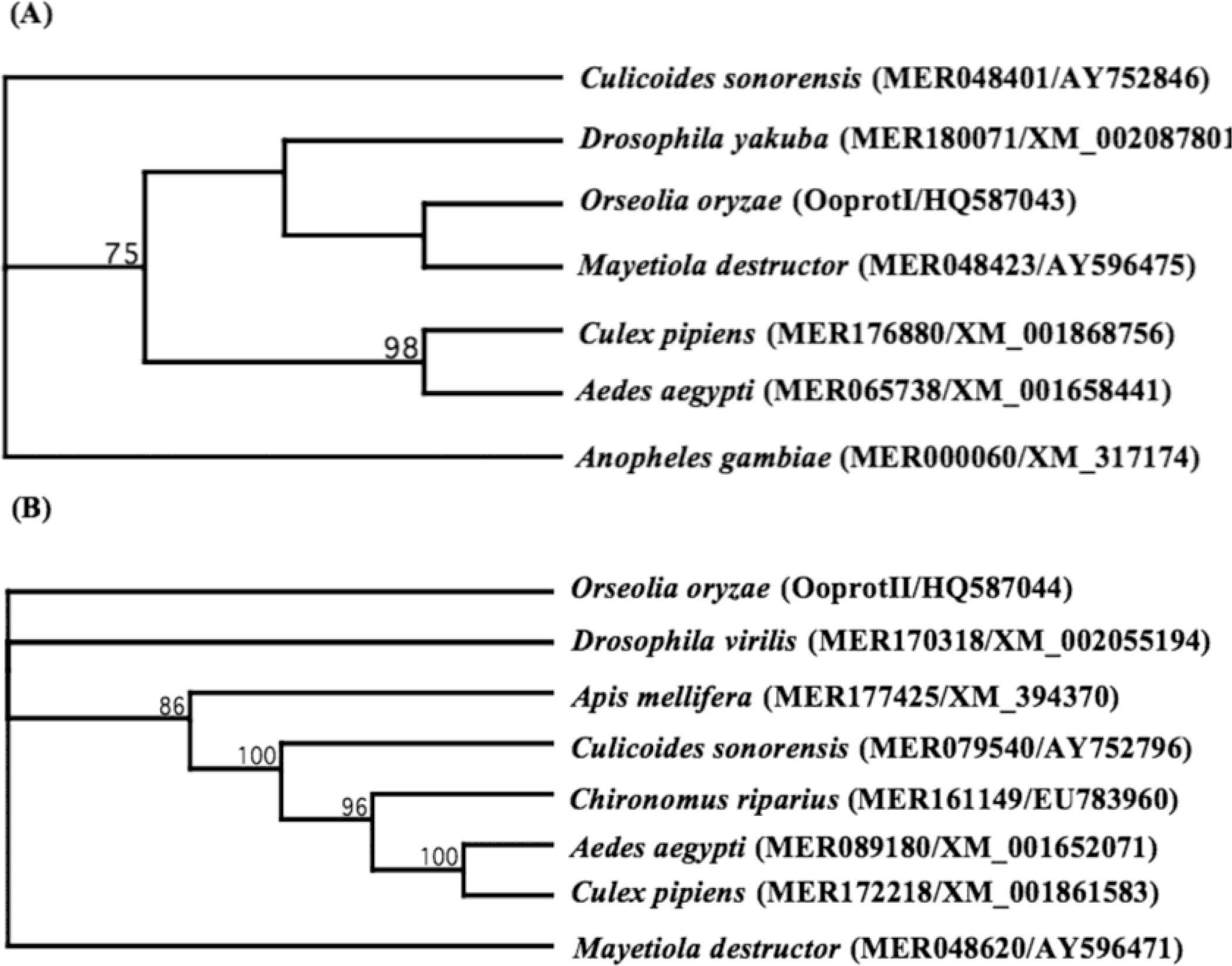
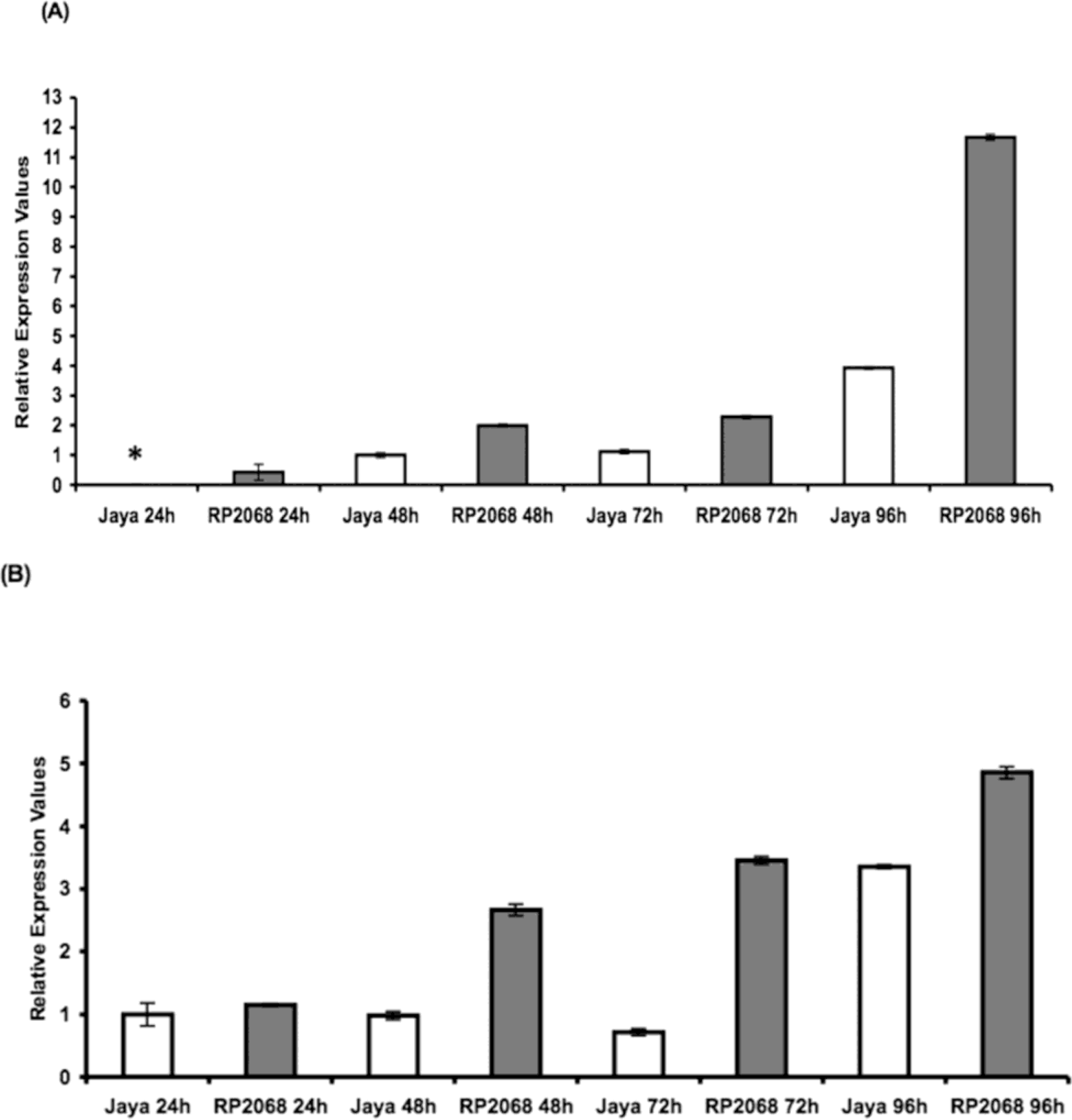
2. Results and Discussion
3. Experimental Section
3.1. Insect Material
3.2. Collection of Larvae and RNA Isolation
3.3. cDNA Library Preparation
3.4. Sequence and Phylogenetic Analysis
3.5. RT-PCR and Statistical Analyses
4. Conclusions
Acknowledgments
References
- Bentur, JS; Amudhan, S; Pasalu, IC; Sarma, NP; Prasad, RU. Insect-host plant interactions in gall midge–rice. In New Approaches to Gall Midge Resistance in Rice; International Rice Research Institute: Los Baños, Philippines, 2004; pp. 23–29. [Google Scholar]
- Khush, GS; Panda, N. Host Plant Resistance in Rice; International Rice Research Institute: Manila, Philippines, 1995; p. 240. [Google Scholar]
- Vijaya Lakshmi, P; Amudhan, S; Himabindu, K; Cheralu, C; Bentur, JS. A new biotype of the Asian rice gall midge Orseolia oryzae (Diptera: Cecidomyiidae) characterized from the Warangal population in Andhra Pradesh, India. Int. J. Trop. Insect Sci 2006, 26, 207–211. [Google Scholar]
- Sardesai, N; Rajyashri, KR; Behura, SK; Nair, S; Mohan, M. Genetic, physiological and molecular interactions of rice and its major dipteran pest, gall midge. Plant Cell Tissue Organ Cult 2001, 64, 115–131. [Google Scholar]
- Rawat, N; Sinha, DK; Rajendrakumar, P; Shrivastava, P; Neeraja, CN; Sundaram, RM; Nair, S; Bentur, JS. Role of pathogenesis-related genes in rice–gall midge interactions. Curr. Sci 2010, 99, 1361–1368. [Google Scholar]
- Andow, DA; Bentur, JS. Pedigreed crosses to estimate recessive virulence allele frequencies in natural populations of gall midges. Entomol. Exp. Appl 2010, 135, 18–36. [Google Scholar]
- Harris, MO; Stuart, JJ; Mohan, M; Nair, S; Lamb, RJ; Rohfritsch, O. Grasses and gall midges: Plant defense and insect adaptation. Annu. Rev. Entomol 2003, 48, 549–577. [Google Scholar]
- Liu, X; Fellers, JP; Wilde, GE; Stuart, JJ; Chen, MS. Characterization of two genes expressed in the salivary glands of the Hessian fly, Mayetiola destructor (Say). Insect Biochem. Mol. Biol 2004, 34, 229–237. [Google Scholar]
- Baker, B; Zambryski, P; Staskawicz, B; Dinesh-Kumar, SP. Signalling in plant-microbe interactions. Science 1997, 276, 726–733. [Google Scholar]
- Harris, MO; Freeman, TP; Moore, JA; Anderson, KG; Payne, SA; Anderson, KM; Rohfritsch, O. H-Gene-mediated resistance to Hessian fly exhibits features of penetration resistance to fungi. Phytopathology 2010, 100, 279–289. [Google Scholar]
- Zhang, S; Shukle, R; Mittapalli, O; Zhu, YC; Reese, JC; Wang, H; Hua, BZ; Chen, MS. The gut transcriptome of a gall midge, Mayetiola destructor. J. Insect Physiol 2010, 56, 1198–1206. [Google Scholar]
- Broadway, RM. Resistance of plants to herbivorous insects: Can this resistance fail? Can. J. Plant Pathol 1996, 18, 476–481. [Google Scholar]
- Krem, MM; Rose, T; Di Cera, E. The C-terminal sequence encodes function in serine proteases. J. Biol. Chem 1999, 274, 28063–28066. [Google Scholar]
- Page, MJ; Di Cera, E. Serine peptidases: Classification, structure and function. Cell. Mol. Life Sci 2008, 65, 1220–1236. [Google Scholar]
- Yang, L; Fang, Z; Dicke, M; van Loon, JJ; Jongsma, MA. The diamondback moth, Plutella xylostella, specifically inactivates Mustard Trypsin inhibitor 2 (MTI2) to overcome host plant defense. Insect Biochem. Mol. Boil 2009, 39, 55–61. [Google Scholar]
- Jongsma, MA; Bakker, PL; Peters, J; Bosch, D; Stiekema, WJ. Adaptation of Spodoptera exigua larvae to plant proteinase inhibitors by induction of gut proteinase activity insensitive to inhibition. Proc. Natl. Acad. Sci. USA 1995, 92, 8041–8045. [Google Scholar]
- Brioschi, D; Nadalini, LD; Bengtson, MH; Sogayar, MC; Moura, DS; Silva-Filho, MC. General upregulation of Spodoptera frugiperda trypsins and chymotrypsins allows its adaptation to soybean proteinase inhibitor. Insect Biochem. Mol. Biol 2007, 37, 1283–1290. [Google Scholar]
- Kumar, A; Jain, A; Sahu, RK; Shrivastava, MN; Nair, S; Mohan, M. Genetic analysis of resistance genes for the rice gall midge in two rice genotypes. Crop Sci 2005, 45, 1631–1635. [Google Scholar]
- Kraut, J. Serine proteases: Structure and mechanisms of catalysis. Annu. Rev. Biochem 1997, 8, 331–358. [Google Scholar]
- Saikia, M; Singh, YT; Bhattacharya, A; Mazumdar-Leighton, S. Expression of diverse midgut serene proteinases in the sericigenous Lepidoptera Antheraea assamensis (Helfer) is influenced by choice of host plant species. Insect Mol. Biol 2010, 20, 1–13. [Google Scholar]
- Zhu-Salzman, K; Bi, JL; Liu, TX. Molecular strategies of plant defense and insect counter-defense. Insect Sci 2005, 12, 3–15. [Google Scholar]
- Vandesompele, J; De Preter, K; Pattyn, F; Poppe, B; van Roy, N; De Paepe, A; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol 2002. 3, research0034.1–research0034.11.. [Google Scholar]



© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Sinha, D.K.; Lakshmi, M.; Anuradha, G.; Rahman, S.J.; Siddiq, E.A.; Bentur, J.S.; Nair, S. Serine Proteases-Like Genes in the Asian Rice Gall Midge Show Differential Expression in Compatible and Incompatible Interactions with Rice. Int. J. Mol. Sci. 2011, 12, 2842-2852. https://doi.org/10.3390/ijms12052842
Sinha DK, Lakshmi M, Anuradha G, Rahman SJ, Siddiq EA, Bentur JS, Nair S. Serine Proteases-Like Genes in the Asian Rice Gall Midge Show Differential Expression in Compatible and Incompatible Interactions with Rice. International Journal of Molecular Sciences. 2011; 12(5):2842-2852. https://doi.org/10.3390/ijms12052842
Chicago/Turabian StyleSinha, Deepak Kumar, Mulagondla Lakshmi, Ghanta Anuradha, Shaik J. Rahman, Ebrahimali A. Siddiq, Jagadish S. Bentur, and Suresh Nair. 2011. "Serine Proteases-Like Genes in the Asian Rice Gall Midge Show Differential Expression in Compatible and Incompatible Interactions with Rice" International Journal of Molecular Sciences 12, no. 5: 2842-2852. https://doi.org/10.3390/ijms12052842