An Oligodeoxynucleotide with Promising Modulation Activity for the Proliferation and Activation of Osteoblast

Abstract
:1. Introduction
2. Results and Discussion
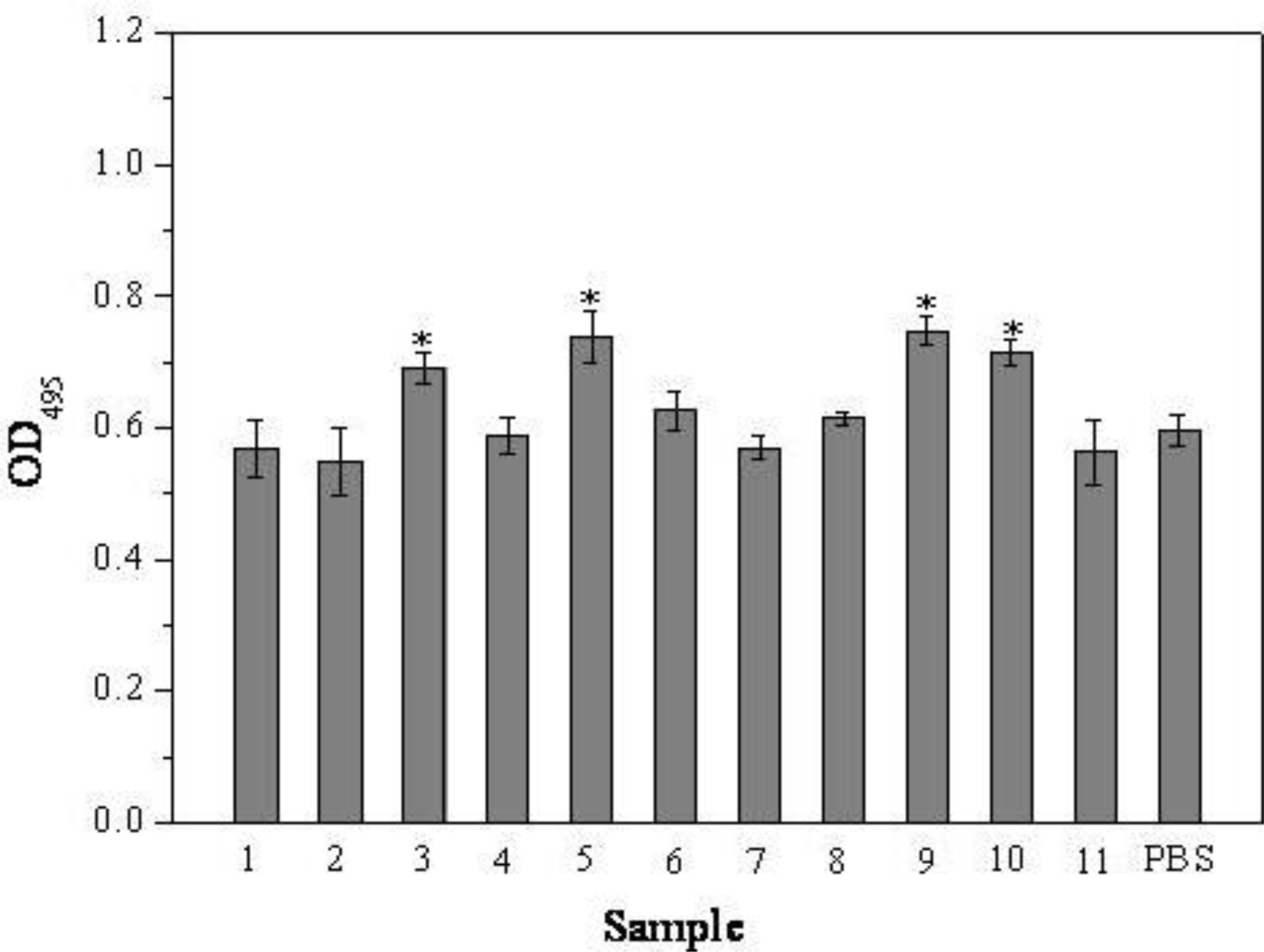
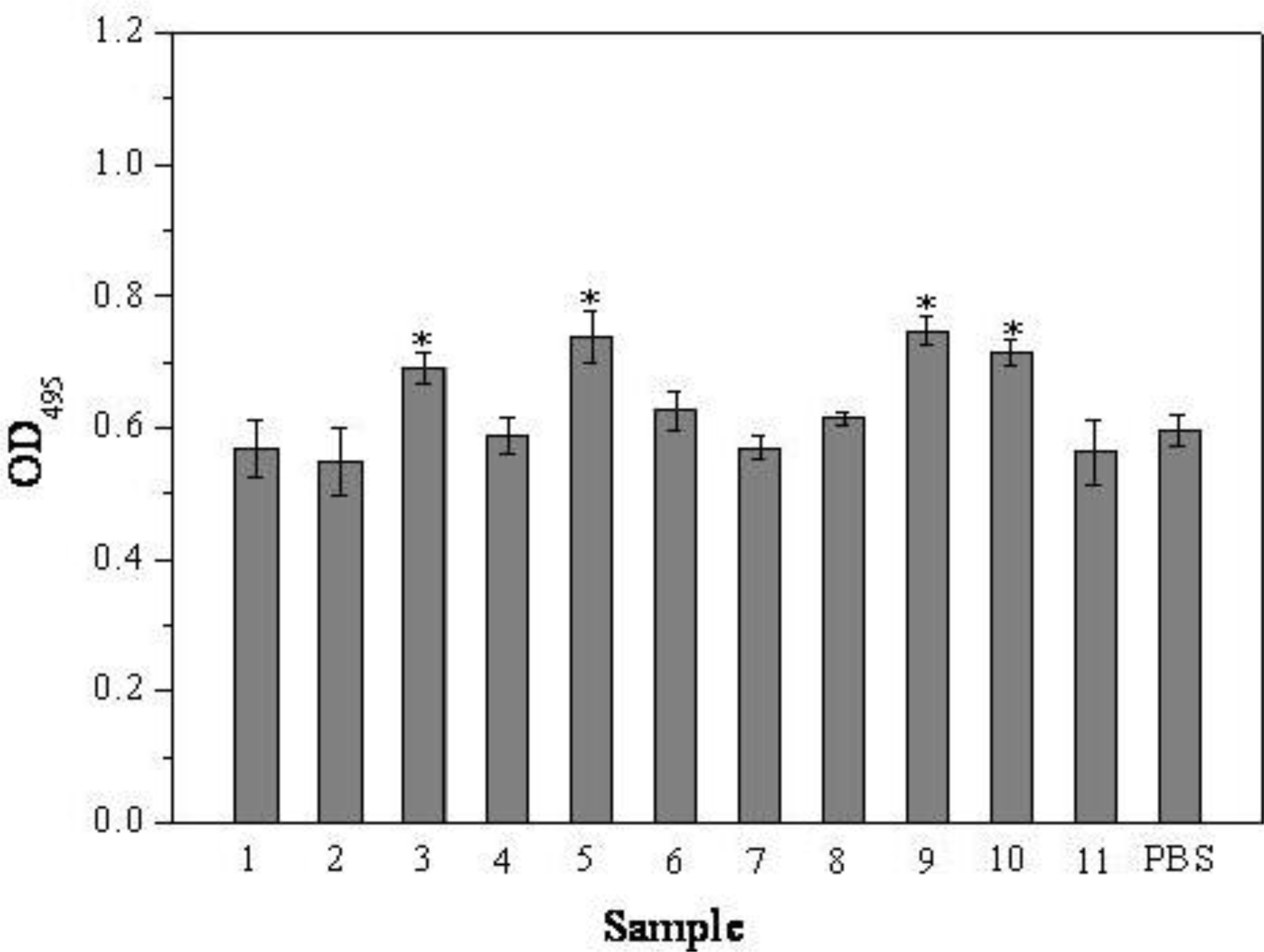
2.1. MTT Assay
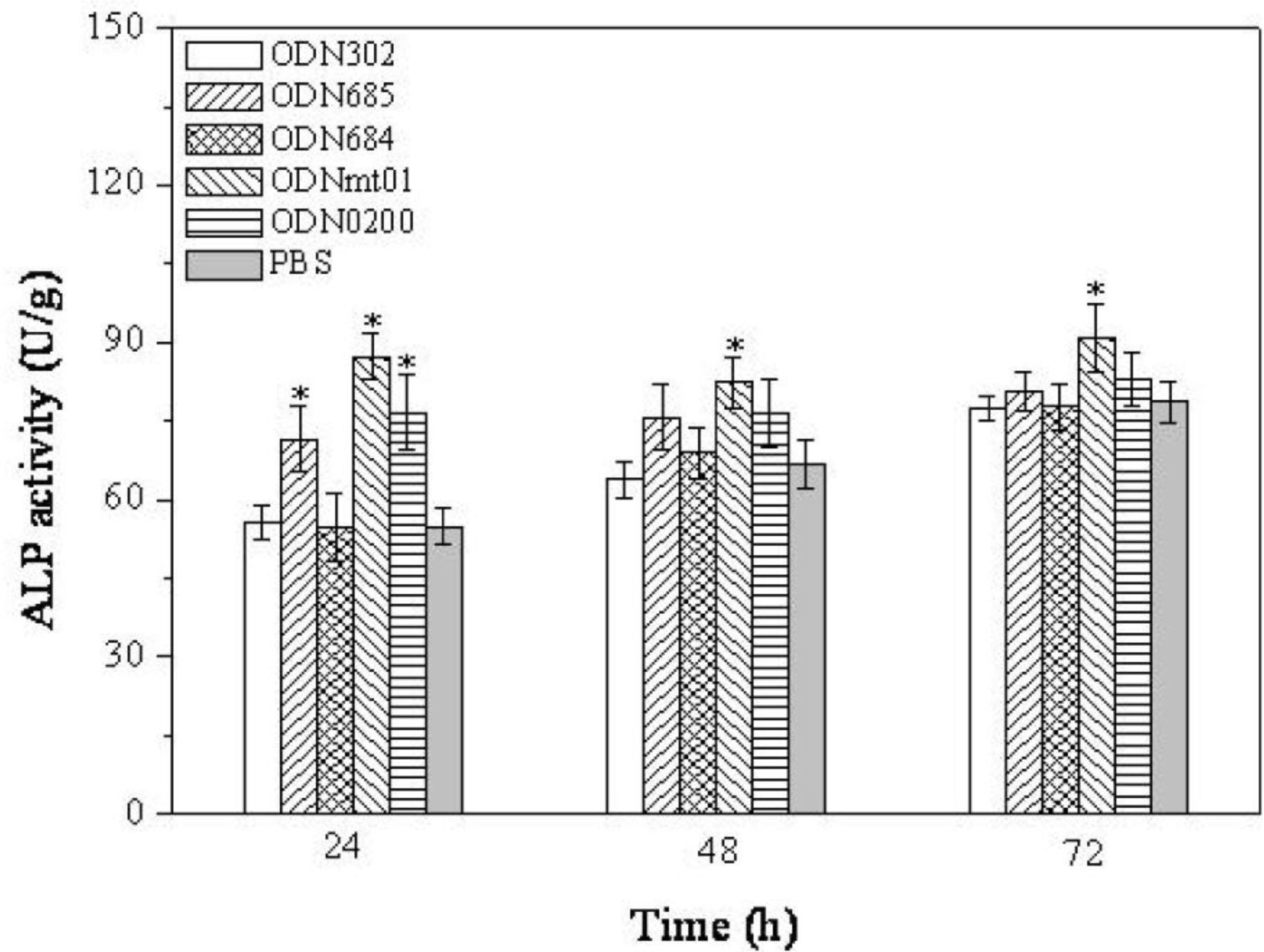
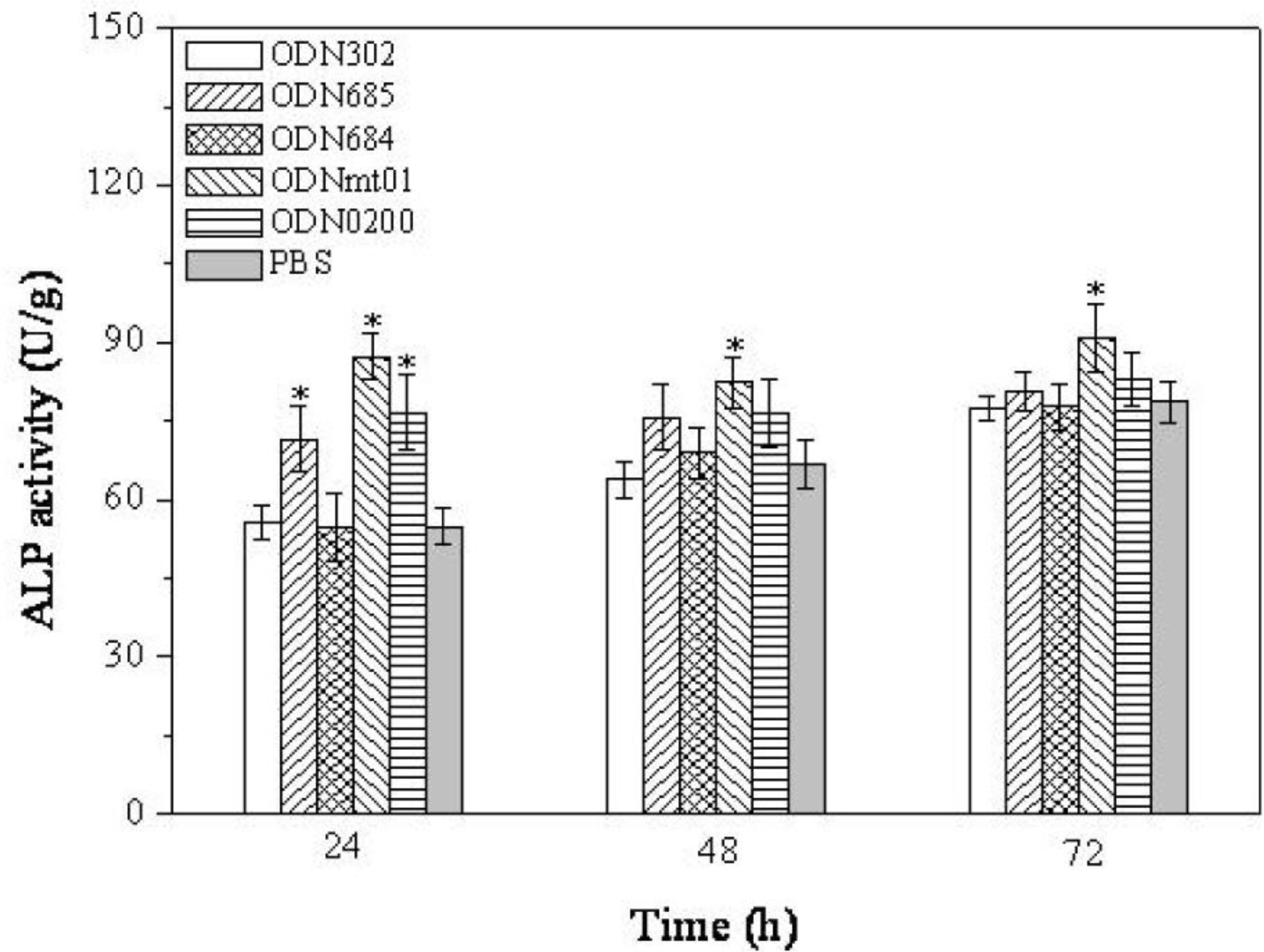
2.2. Effect of ODNs on ALP Activity
2.3. Effect of ODN MT01 on the Morphology of MG 63 Cells
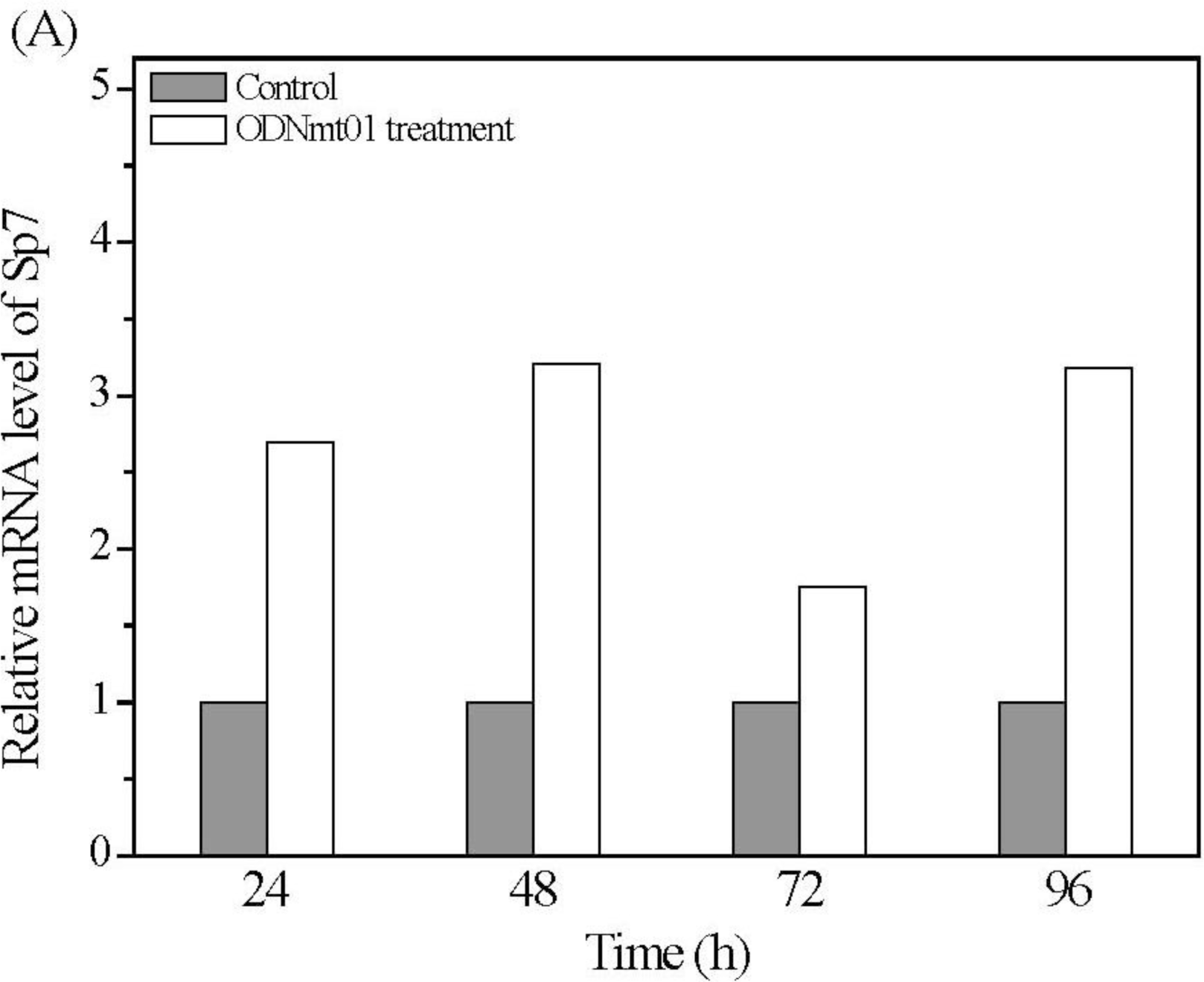
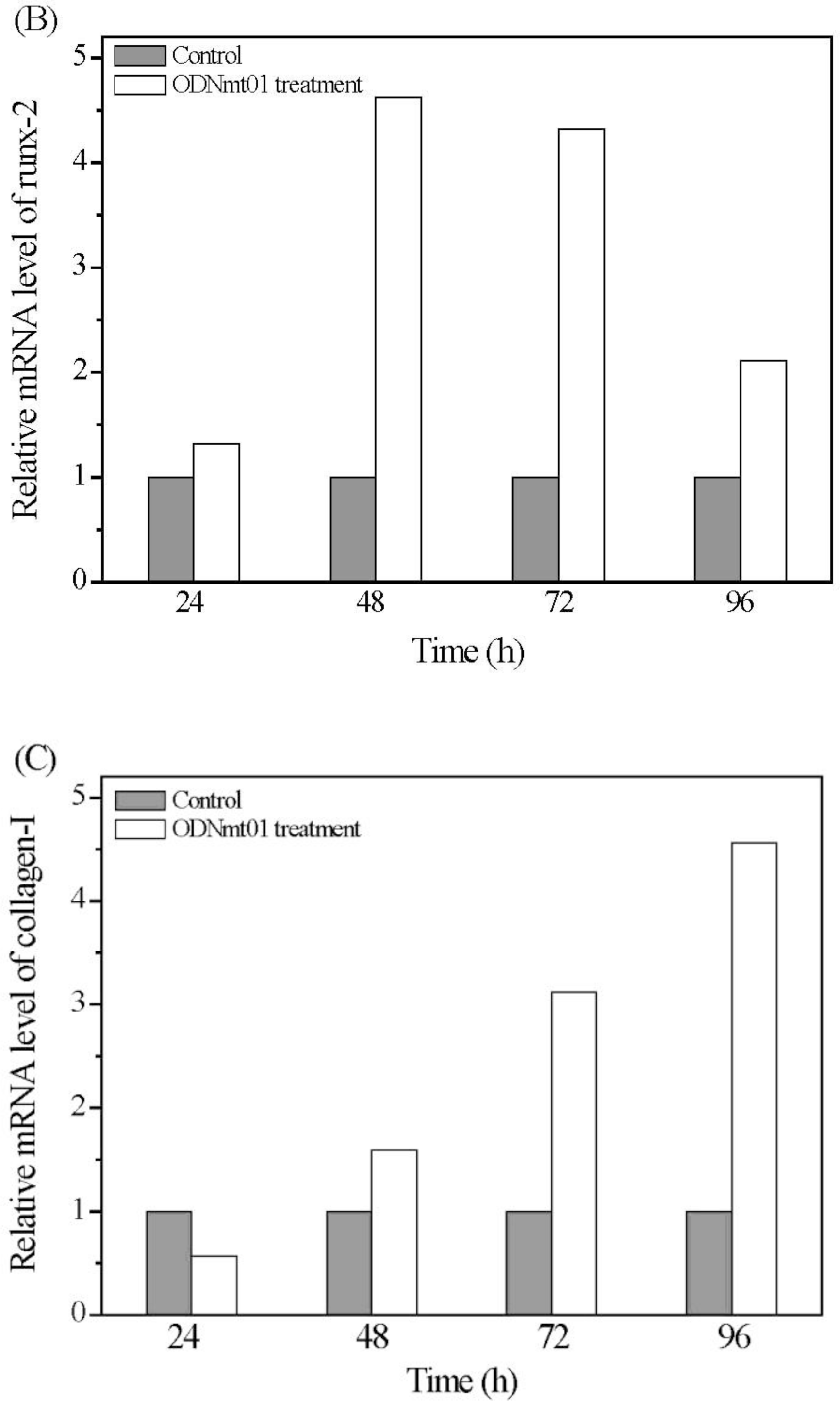
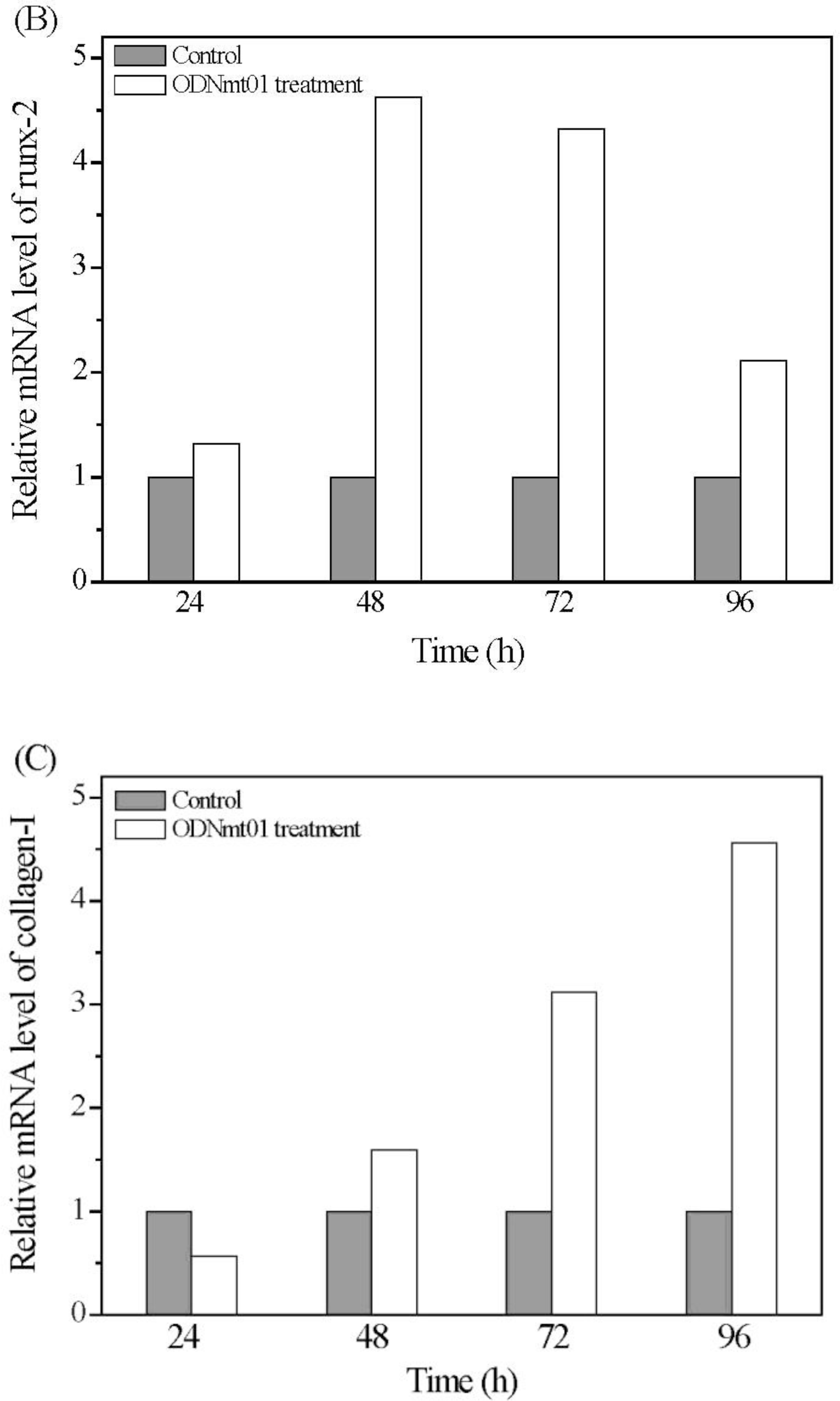
2.4. Effect of ODN MT01 on Gene Expression of MG 63 Cells
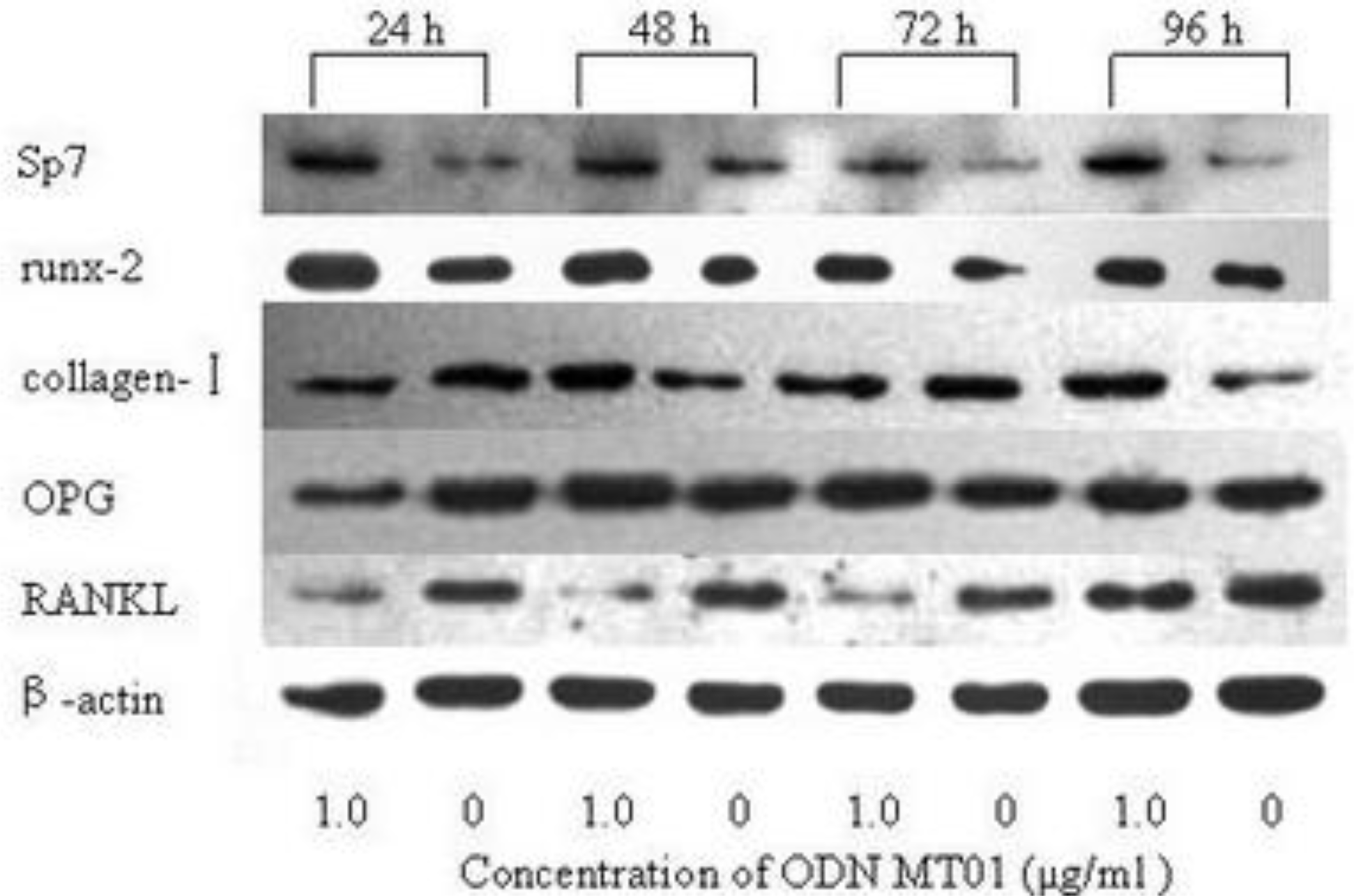
2.5. Western Blotting Analysis
3. Experimental Section
3.1. Materials
3.2. Cell Culture
3.3. MTT Assay
3.4. Measurement of ALP Activity
3.5. Real-Time PCR
3.6. Western Blotting
3.7. Statistical Analysis
4. Conclusions
Acknowledgments
References
- Verna, C; Zaffe, D; Siciliani, G. Histomorphometric study of bone reactions during orthodontic tooth movement in rats. Bone 1999, 24, 371–379. [Google Scholar]
- Graves, DT; Kayal, RA; Oates, T. Osteoimmunology in the oral cavity (periodontal disease, lesions of endodontic origin and orthodontic tooth movement). Osteoimmunology 2011, 15, 411–441. [Google Scholar]
- Ducy, P; Schinke, T; Karsenty, G. The osteoblast: A sophisticated fibroblast under central surveillance. Science 2000, 289, 1501–1504. [Google Scholar]
- Teitelbaum, SL. Bone resorption by osteoclasts. Science 2000, 289, 1504–1508. [Google Scholar]
- Kim, SH; Yoo, CI; Kim, HT. Activation of peroxisome proliferator-activated receptor-γ (PPARγ) induces cell death through MAPK-dependent mechanism in osteoblastic cells. Toxicol. Appl. Pharmacol 2006, 215, 198–207. [Google Scholar]
- Ripamonti, U; Petit, J-C. Bone morphogenetic proteins, cementogenesis, myoblastic stem cells and the induction of periodontal tissue regeneration. Cytokine Growth Factor Rev 2009, 20, 489–499. [Google Scholar]
- Wikesjö, UME; Huang, YH; Polimeni, G; Qahash, M. Bone Morphogenetic proteins: A realistic alternative to bone grafting for alveolar reconstruction. Oral Maxillofac. Surg. Clin. North Am 2007, 19, 535–551. [Google Scholar]
- Fischer, J; Kolk, A; Wolfart, S; Pautkeb, C; Warnke, PH; Plank, C; Smeets, R. Future of local bone regeneration—Protein versus gene therapy. J. Cranio-Maxillofac. Surg 2011, 39, 54–64. [Google Scholar]
- Nada, RM; Sugar, AW; Wijdeveld, MGMM; Borstlap, WA; Clauser, L; Hoffmeister, B; Kuijpers-Jagtman, AM; The Eurocran Distraction Osteogenesis Group. Current practiceof distraction osteogenesis for craniofacial anomalies in Europe: A web based survey. J. Cranio-Maxillofac. Surg 2010, 38, 83–89. [Google Scholar]
- Vega, LG; Bilbao, A. Alveolar distraction osteogenesis for dental implant preparation: An update. Oral Maxillofac. Surg. Clin. North Am 2010, 22, 369–385. [Google Scholar]
- Lee, E-J; Shin, D-S; Kim, HE; Koh, Y-H; Jang, J-H. Membrane of hybrid chitosan-silica xerogel for guided bone regeneration. Biomaterials 2009, 30, 743–750. [Google Scholar]
- Park, KH; Han, DI; Rhee, Y-H; Jeong, S-J; Kim, SH; Park, YG. Protein kinase C βII and δ/θ play critical roles in bone morphogenic protein-4-stimulated osteoblastic differentiation of MC3T3-E1 cells. Biochem. Biophys. Res. Commun 2010, 403, 7–12. [Google Scholar]
- Yang, DH; Guo, J; Divieti, PF; Bringhurst, R. Parathyroid hormone activates PKC-δ and regulates osteoblastic differentiation via a PLC-independent pathway. Bone 2006, 38, 485–496. [Google Scholar]
- Heikenwalder, M; Polymenidou1, M; Junt, T; Sigurdson, C; Wagner, H; Akira, S; Zinkernagel, R; Aguzzi1, A. Lymphoid follicle destruction and immunosuppression after repeated CpG oligodeoxynucleotide administration. Nat. Med 2004, 10, 187–192. [Google Scholar]
- Penolazzi, L; Lambertini, E; Borgatti, M. Decoy oligodeoxynucleotides targeting NF-kappaB transcription factors: induction of apoptosis in human primary osteoclasts. Biochem. Pharmacol 2003, 66, 1189–1198. [Google Scholar]
- Amcheslavsky, A; Zou, W; Bar-Shavit, Z. Toll-like receptor 9 regulates tumor necrosis factor-α expression by different mechanisms implications for osteoclastogenesis. J. Biol. Chem 2004, 279, 54039–54045. [Google Scholar]
- Chang, J-H; Chang, E-J; Kim, H-H; Kim, S-K. Enhanced inhibitory effects of a novel CpG motif on osteoclast differentiation via TREM-2 down-regulation. Biochem. Biophys. Res. Commun 2009, 389, 28–33. [Google Scholar]
- Jacobson, A; Johansson, S; Branting, M; Melhus, H. Vitamin A differentially regulates RANKL and OPG expression in human osteoblasts. Biochem. Biophys. Res. Commun 2004, 322, 162–167. [Google Scholar]
- Zou, W; Amcheslavsky, A; Bar-Shavit, Z. CpG Oligodeoxynucleotides Modulate the Osteoclastogenic Activity of Osteoblasts via Toll-like Receptor 9. J. Biol. Chem 2003, 278, 16732–16740. [Google Scholar]
- Amcheslavsky, A; Hemmi, H; Akira, S; Bar-Shavit, Z. Differential contribution of osteoclast- and osteoblast-lineage cells to CpG-oligodeoxynucleotide (CpG-ODN) modulation of osteoclastogenesis. J. Bone Miner. Res 2005, 20, 1692–1699. [Google Scholar]
- Nørgaard, NN; Holien, T; Jonsson, S; Hella, H; Espevik, T; Sundan, A; Standal, T. CpG-Oligodeoxynucleotide inhibits Smad-dependent bone morphogenetic protein signaling: Effects on myeloma cell apoptosis and in vitro osteoblastogenesis. J. Immunol 2010, 185, 3131–3139. [Google Scholar]
- Yoshimatsu, M; Kitaura, H; Fujimura, YJ; Eguchi, T; Kohara, H; Morita, Y; Yoshidaa, N. IL-12 inhibits TNF-α induced osteoclastogenesis via a T cell-independent mechanism in vivo. Bone 2009, 45, 1010–1016. [Google Scholar]
- Choi, JY; Pratap, J; Javed, A; Zaidi, SK; Xing, LP; Eva Balint, E; Dalamangas, S; Boyce, B; Van Wijnen, AJ; et al. Subnuclear targeting of Runx/Cbfa/AML factors is essential for tissue-specific differentiation during embryonic development. Proc. Natl. Acad. Sci. USA 2001, 98, 8650–8655. [Google Scholar]
- Pratap, J; Pratap, J; Galindo, M; Zaidi, SK; Vradii, D; Bhat, BM; Robinson, JA; Choi, J-Y; Komori, T; Stein, JL; Lian, JB; et al. Cell growth regulatory role of Runx2 during proliferative expansion of preosteoblasts. Cancer Res 2003, 63, 5357–5362. [Google Scholar]
- Nakashima, K; Zhou, X; Kunkel, G; Zhang, Z; Deng, JM; Behringer, RR; de Crombrugghe, B. The novel zinc fingercontaining transcription factor osterix is required for osteoblast differentiation and bone formation. Cell 2002, 108, 17–29. [Google Scholar]
- Kaback, LA; Soung, DY; Naik, A; Smith, N; Schwarz, EM; O’Keefe, RJ; Drissi, H. Osterix/Sp7 regulates mesenchymal stem cell mediated endochondral ossification. J. Cell. Physiol 2008, 214, 173–182. [Google Scholar]
- Lee, MH; Kwon, TG; Park, HS; Wozney, JM; Ryoo, HM. BMP-2-induced Osterix expression is mediated by Dlx5 but is independent of Runx2. Biochem. Biophys. Res. Commun 2003, 309, 689–694. [Google Scholar]
- Kim, Y-J; Kim, H-N; Park, E-K; Lee, B-H; Ryoo, H-M; Kim, S-Y; Kim, I-S; Stein, JL; Lian, JB; Stein, GS; et al. The bone-related Zn finger transcription factor Osterix promotes proliferation of mesenchymal cells. Gene 2006, 366, 145–151. [Google Scholar]
- Khosla, S. Minireview: the OPG/RANKL/RANK system. Endocrinology 2001, 142, 5050–5055. [Google Scholar]
- Tat, SK; Padrines, M; Theoleyre, S; Heymann, D; Fortun, Y. IL-6, RANKL, TNF-alpha/IL-1: Interrelations in bone resorption pathophysiology. Cytokine Growth Factor Rev 2004, 15, 49–60. [Google Scholar]
- Bao, M; Zhang, Y; Wan, M; Dai, L; Hu, XP; Wu, XL; Wang, L; Deng, P; Wang, JZ; Chen, JZ; et al. Anti-SARS-CoV immunity induced by a novel CpG oligodeoxynucleotide. Clin. Immunol 2006, 118, 180–187. [Google Scholar]
- Wang, X; Bao, M; Wan, M; Wei, HF; Wang, L; Yu, HT; Zhang, XS; Yu, YL; Wang, LY. A CpG oligodeoxynucleotide acts as a potent adjuvant for inactivated rabies virus vaccine. Vaccine 2008, 26, 1893–1901. [Google Scholar]
- Yang, L; Sun, LG; Wu, XL; Wang, L; Wei, HF; Wan, M; Zhang, PY; Yu, YL; Wang, LY. Therapeutic injection of C-class CpG ODN in draining lymph node area induces potent activation of immune cells and rejection of established breast cancer in mice. Clin. Immunol 2009, 131, 426–437. [Google Scholar]
- Sun, R; Sun, LG; Bao, MS; Zhang, YS; Wang, L; Wu, XL; Hu, DL; Liu, YJ; Yu, YL; Wang, LY. A human microsatellite DNA-mimicking oligodeoxynucleotide with CCT repeats negatively regulates TLR7/9-mediated innate immune responses via selected TLR pathways. Clin. Immunol 2010, 134, 262–276. [Google Scholar]
- Yang, G; Wan, M; Zhang, YS; Sun, LG; Sun, R; Hu, DL; Zhou, XJ; Wang, L; Wu, XL; Wang, LY; Yu, YL. Inhibition of a C-rich oligodeoxynucleotide on activation of immune cells in vitro and enhancement of antibody response in mice. Immunology 2010, 131, 501–512. [Google Scholar]
- Schmittgen, TD; Livak, KJ. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc 2008, 3, 1101–1108. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number | Name | Sequence |
|---|---|---|
| 1 | SAT05f | 5′-CCTCCTCCTCCTCCTCCTCCTCCT-3′ |
| 2 | MS19 | 5′-AAAGAAAGAAAGAAAGAAAGAAAG-3′ |
| 3 | BW001 | 5′-TCGTCGGGTGCGACGTCGCAGGGGGG-3′ |
| 4 | FC001 | 5′-TCGGGGACGATCGTCGGGGAC-3′ |
| 5 | FC002 | 5′-TCGTCGACGTCGTCGTTCTC-3′ |
| 6 | BW006 | 5′-TCGACGTTCGTCGTTCGTCGTTC-3′ |
| 7 | YW002 | 5′-TCGCGAACGTTCGCCGCGTTCGAACGCGG-3′ |
| 8 | FC004 | 5′-TCGCGTTCGATCGCGATCGACGGTA-3′ |
| 9 | MT01 | 5′-ACCCCCTCTACCCCCTCTACCCCCTCT-3′ |
| 10 | YW001 | 5′-TCGCGACGTTCGCCCGACGTTCGGTA-3′ |
| 11 | FC003 | 5′-TCTCTCTCTCTCTCTCTCTCTCTC-3′ |
| Gene Name | Oligonucleotide UP (5′-3′) | Oligonucleotide DW (5′-3′) |
|---|---|---|
| GAPDH | ATG GGG AAG GTG AAG GTC | TAA AAG CAG CCC TGG TGA CC |
| SP7 | CAC AGC TCT TCT GAC TGT CTG CTG | GTG AAA TGC CTG CAT GGA T |
| RUNX-2 | GAG ATC ATC GCC GAC CAC | TAC CTC TCC GAG GGC TAC C |
| COLLAGEN-I | AGG GCC AAG ACG AAG ACA TC | AGA TCA CGT CAT CGC ACA ACA |
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Feng, Z.; Shen, Y.; Wang, L.; Cheng, L.; Wang, J.; Li, Q.; Shi, W.; Sun, X. An Oligodeoxynucleotide with Promising Modulation Activity for the Proliferation and Activation of Osteoblast. Int. J. Mol. Sci. 2011, 12, 2543-2555. https://doi.org/10.3390/ijms12042543
Feng Z, Shen Y, Wang L, Cheng L, Wang J, Li Q, Shi W, Sun X. An Oligodeoxynucleotide with Promising Modulation Activity for the Proliferation and Activation of Osteoblast. International Journal of Molecular Sciences. 2011; 12(4):2543-2555. https://doi.org/10.3390/ijms12042543
Chicago/Turabian StyleFeng, Zhiyuan, Yuqin Shen, Liying Wang, Lin Cheng, Jing Wang, Quanshun Li, Wei Shi, and Xinhua Sun. 2011. "An Oligodeoxynucleotide with Promising Modulation Activity for the Proliferation and Activation of Osteoblast" International Journal of Molecular Sciences 12, no. 4: 2543-2555. https://doi.org/10.3390/ijms12042543