Enhancement of Salinity Tolerance during Rice Seed Germination by Presoaking with Hemoglobin


Abstract
:1. Introduction
2. Results
2.1. The Inhibition of Seed Germination and Seedling Shoot Growth Were Alleviated by the Pretreatment of Hemoglobin
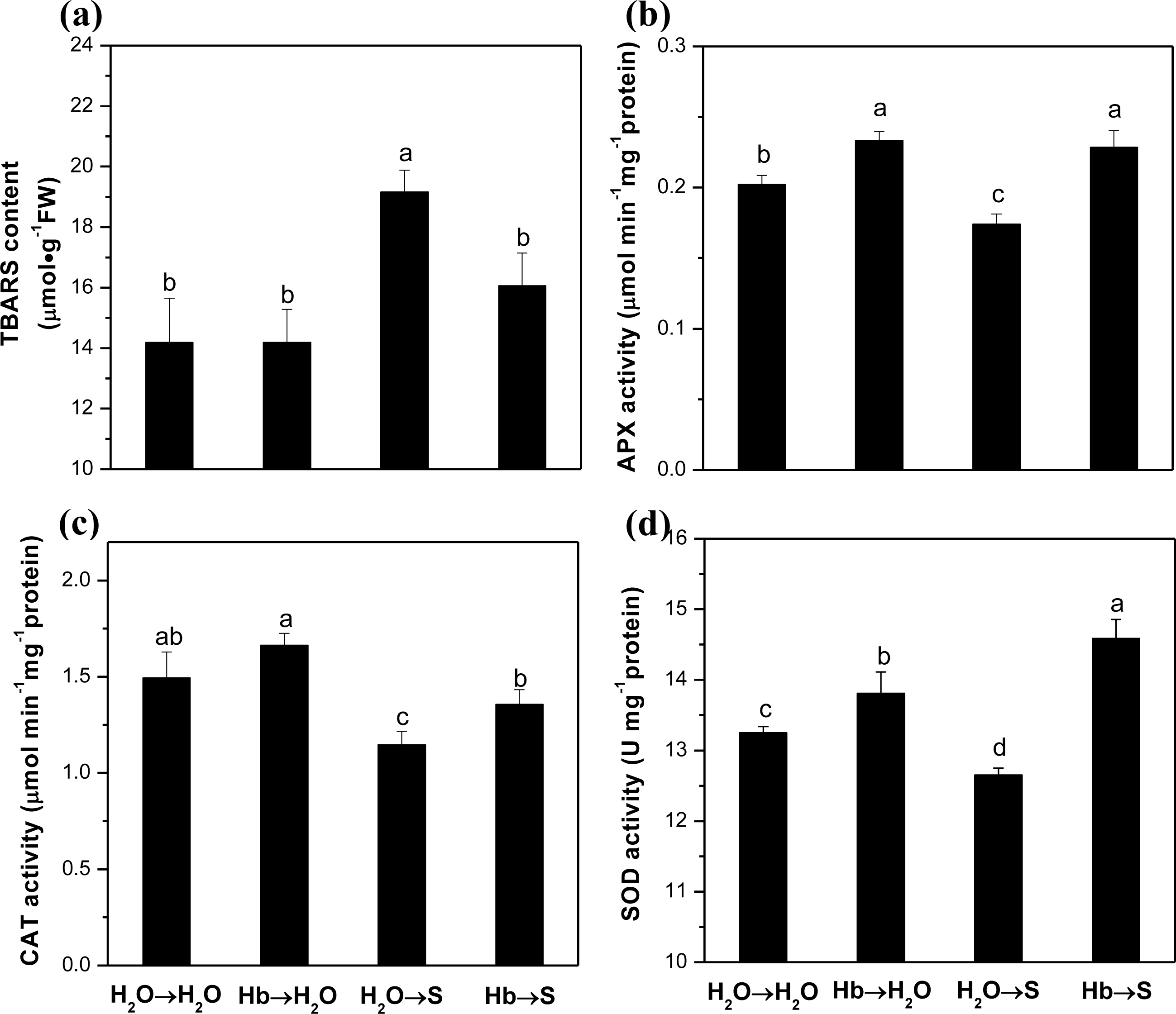
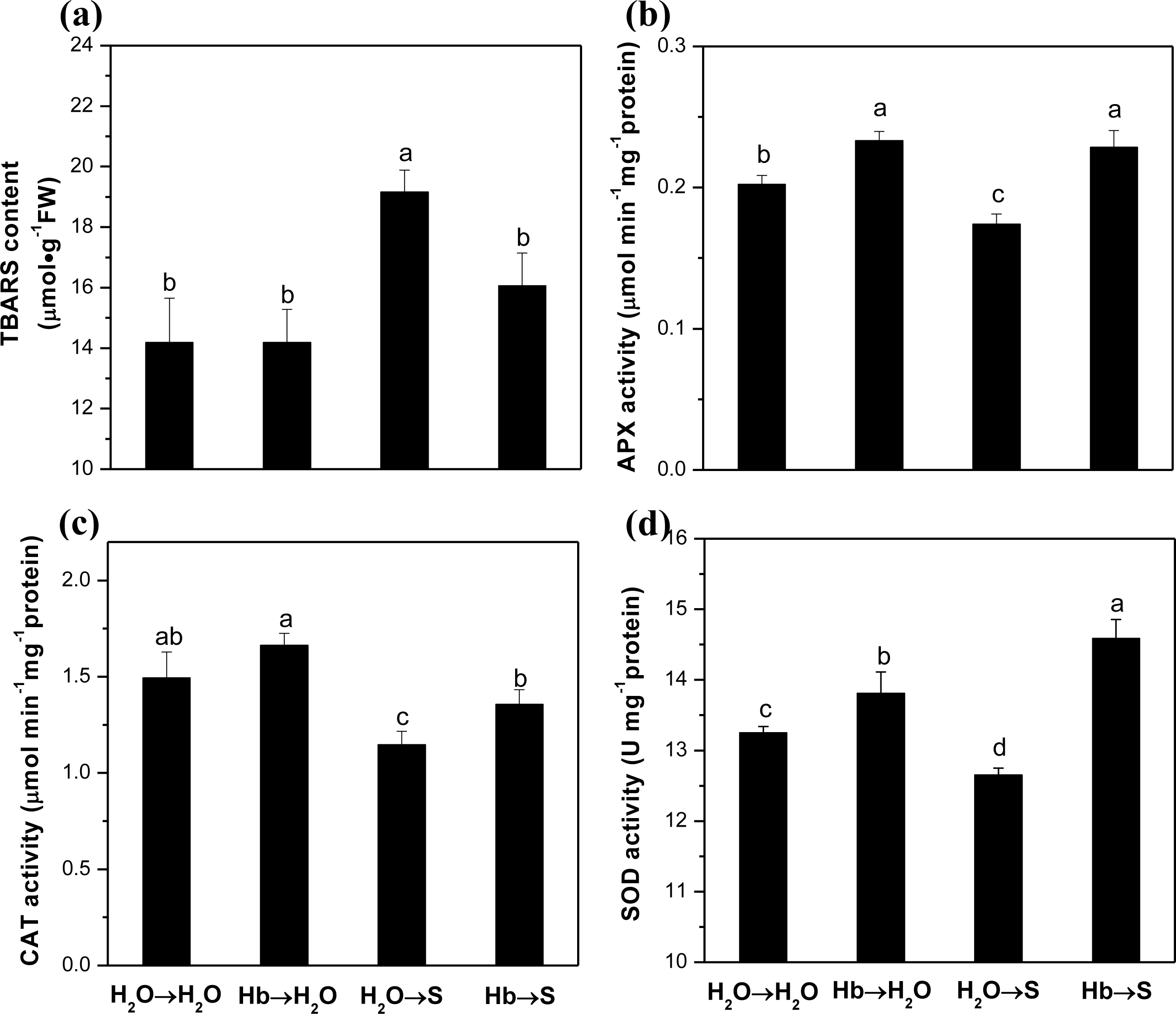
2.2. Lipid Peroxidation and ROS-Scavenging Enzyme Activities
2.3. Antioxidant Enzyme Transcripts
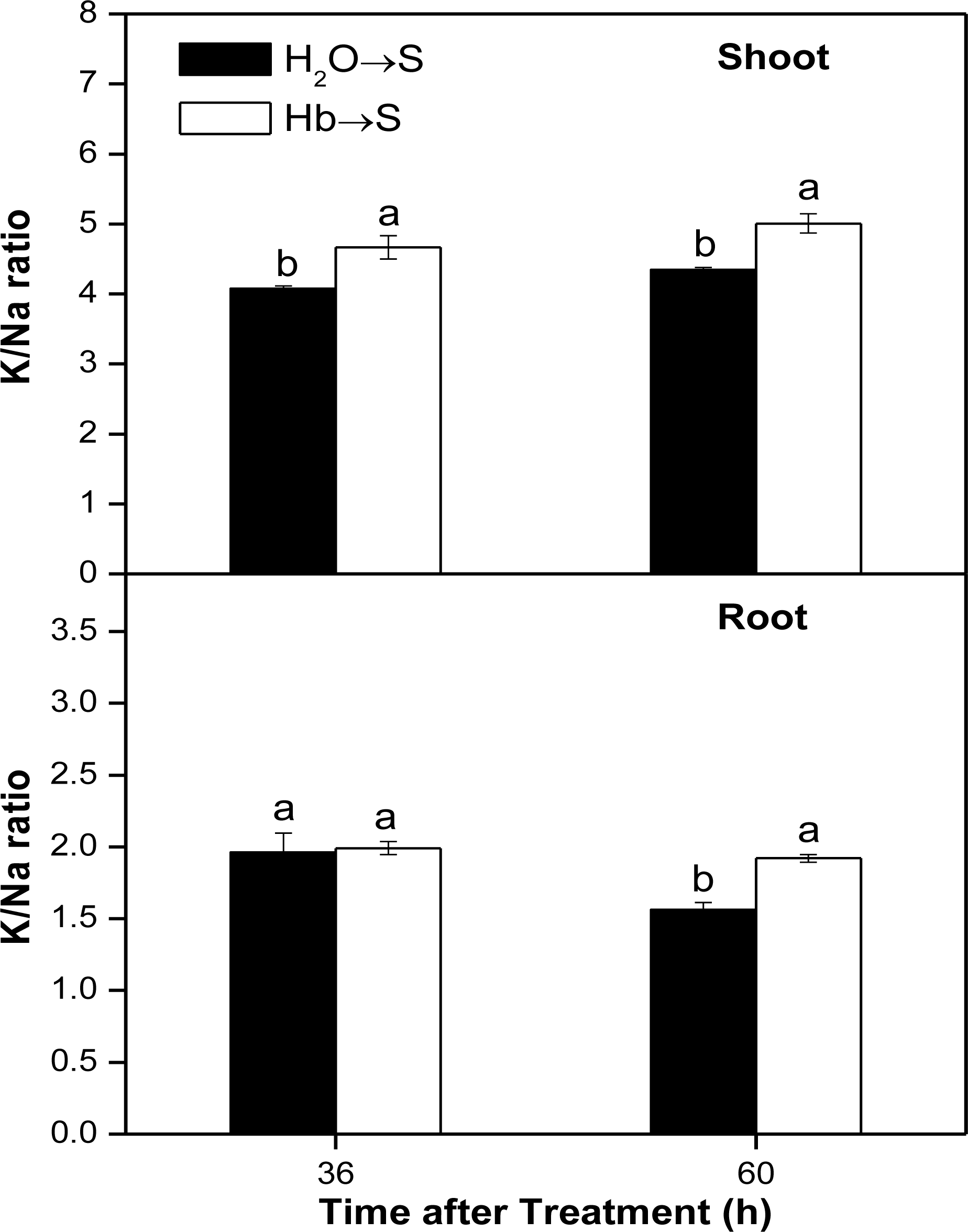
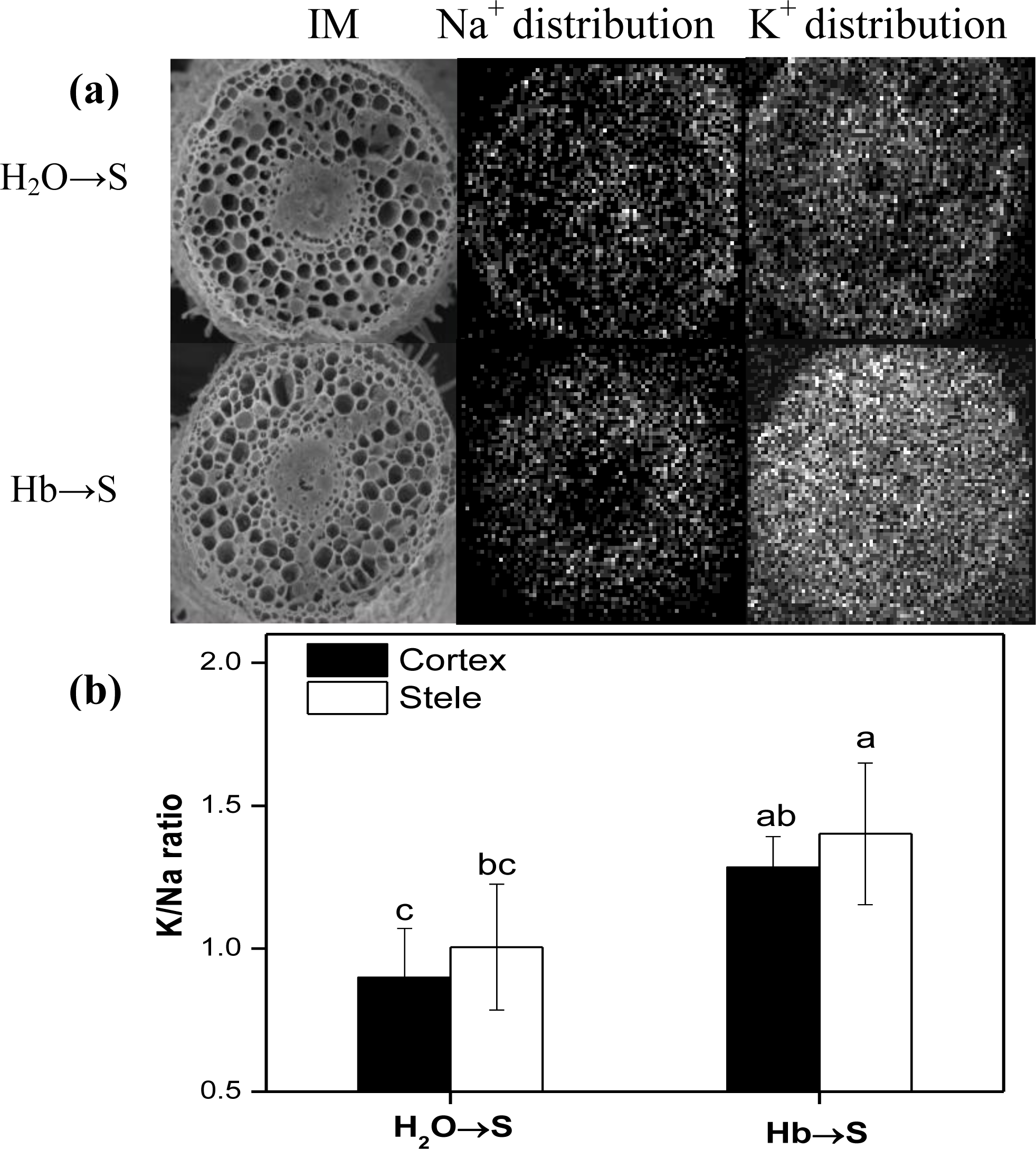
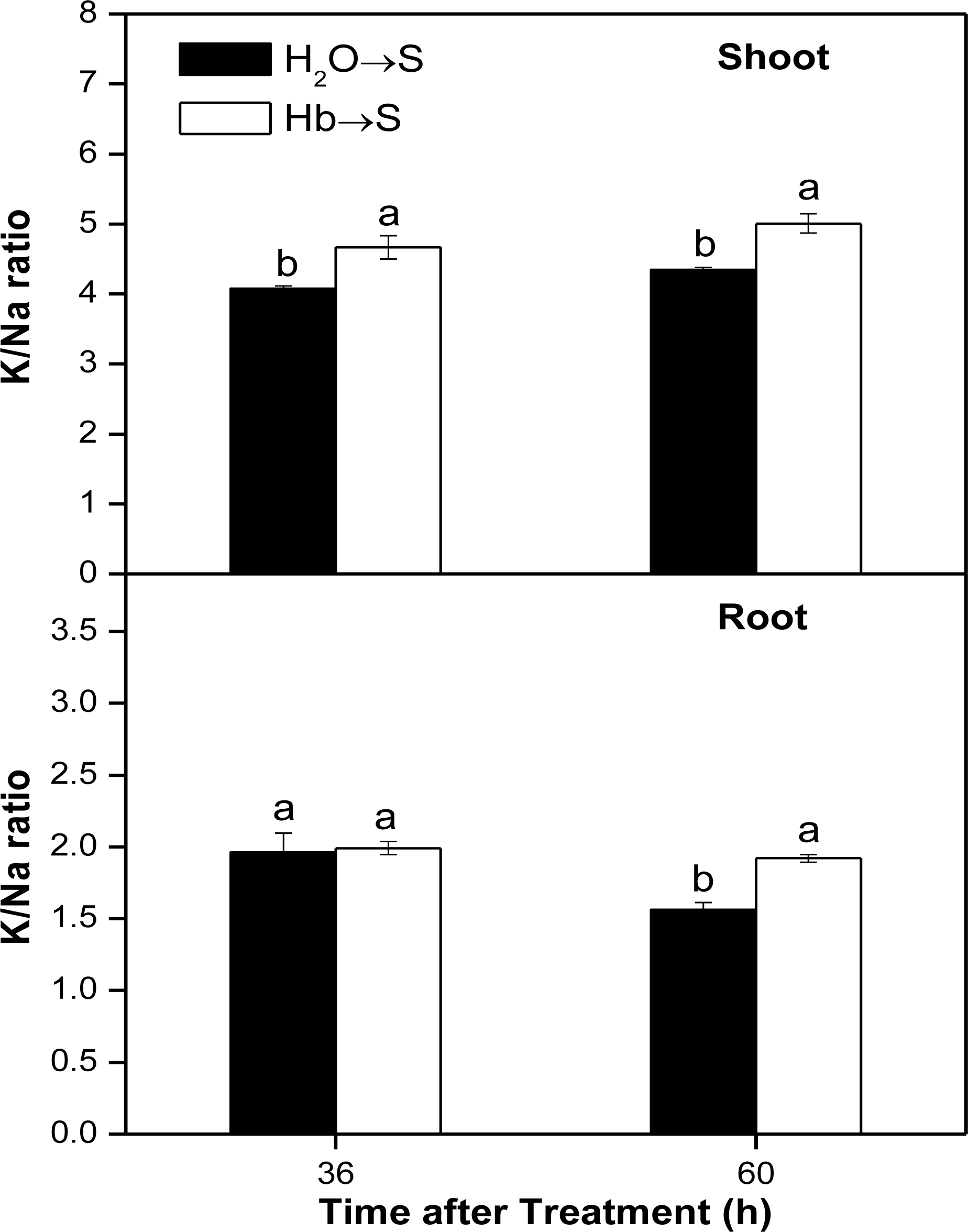
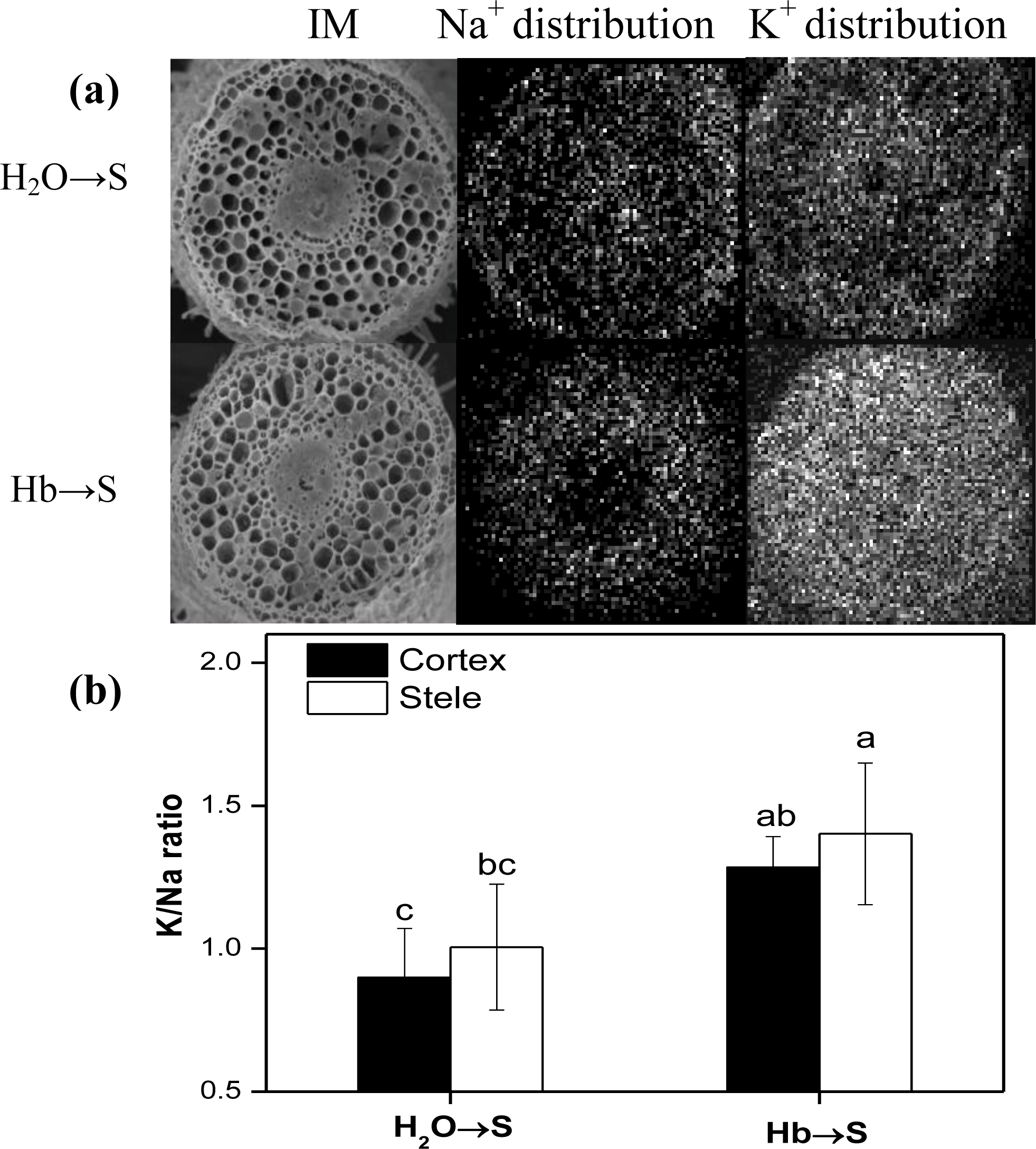
2.4. Changes of K/Na Ratio
3. Discussion
4. Experimental Section
4.1. Chemicals
4.2. Plant Material and Experimental Design
4.3. Germination and Growth Analysis
4.4. TBARS Determination
4.5. Antioxidant Enzyme Assays
4.6. Semi-Quantitative RT-PCR Analysis
4.7. Determination of Ion Contents
4.8. X-ray Microanalysis
4.9. Statistical Analysis
5. Conclusions
Acknowledgments
References
- Chang, C; Wang, B; Shi, L; Li, Y; Duo, L; Zhang, W. Alleviation of salt stress-induced inhibition of seed germination in cucumber (Cucumis sativus L.) by ethylene and glutamate. J. Plant Physiol 2010, 167, 1152–1156. [Google Scholar]
- Khan, AA; Huang, XL. Synergistic enhancement of ethylene production and germination with kinetin and 1-aminocyclopropane-1-carboxylic acid in lettuce seeds exposed to salinity stress. Plant Physiol 1988, 87, 847–852. [Google Scholar]
- Kaur, S; Gupta, AK; Kaur, N. Gibberellin A3 reverses the effect of salt stress in chickpea (Cicer arietinum L.) seedlings by enhancing amylase activity and mobilization of starch in cotyledons. Plant Growth Regul 1998, 26, 85–90. [Google Scholar]
- Çavusoglu, K; Kabar, K. Effects of hydrogen peroxide on the germination and early seedling growth of barley under NaCl and high temperature stresses. EurAsia. J. BioSci 2010, 4, 70–79. [Google Scholar]
- Zheng, CF; Jiang, D; Liu, FL; Dai, TB; Liu, WC; Jing, Q; Cao, WX. Exogenous nitric oxide improves seed germination in wheat against mitochondrial oxidative damage induced by high salinity. Environ. Exp. Bot 2009, 67, 222–227. [Google Scholar]
- Liu, KL; Xu, S; Xuan, W; Ling, TF; Cao, ZY; Huang, BK; Sun, YG; Fang, L; Liu, ZY; Zhao, N; Shen, WB. Carbon monoxide counteracts the inhibition of seed germination and alleviates oxidative damage caused by salt stress in Oryza sativa. Plant Sci 2007, 172, 544–555. [Google Scholar]
- Zhu, JK. Plant salt tolerance. Trends Plant Sci 2001, 6, 66–71. [Google Scholar]
- Muramoto, T; Tsurui, N; Terry, MJ; Yokota, A; Kohchi, T. Expression and biochemical properties of a ferredoxin-dependent heme oxygenase required for phytochrome chromophore synthesis. Plant Physiol 2002, 130, 1958–1966. [Google Scholar]
- Davis, SJ; Bhoo, SH; Durski, AM; Walker, JM; Vierstra, RD. The heme-oxygenase family required for phytochrome chromophore biosynthesis is necessary for proper photomorphogenesis in higher plants. Plant Physiol 2001, 126, 656–669. [Google Scholar]
- Shekhawat, GS; Verma, K. Haem oxygenase (HO): An overlooked enzyme of plant metabolism and defence. J. Exp. Bot 2010, 61, 2255–2270. [Google Scholar]
- Xu, S; Zhang, B; Cao, ZY; Ling, TF; Shen, WB. Heme oxygenase is involved in cobalt chloride-induced lateral root development in tomato. Biometals 2011, 24, 181–191. [Google Scholar]
- Choi, AM. Heme oxygenase-1 protects the heart. Circ. Res 2001, 89, 105–107. [Google Scholar]
- Marton, LS; Wang, X; Kowalczuk, A; Zhang, ZD; Windmeyer, E; Macdonald, RL. Effects of hemoglobin on heme oxygenase gene expression and viability of cultured smooth muscle cells. Am. J. Physiol. Heart Circ. Physiol 2000, 279, 2405–2413. [Google Scholar]
- Motterlini, R; Foresti, R; Vandegriff, K; Intaglietta, M; Winslow, RM. Oxidative-stress response in vascular endothelial cells exposed to acellular hemoglobin solutions. Am. J. Physiol. Heart Circ. Physiol 1995, 269, 648–655. [Google Scholar]
- Zhou, J; Zhu, X; Zhang, G; Ling, T. Protective effect of hemoglobin-induced heme oxygenase-1 on injured lungs caused by limb ischemia-reperfusion in rats. Chin. J. Traumatol 2002, 5, 86–91. [Google Scholar]
- Azhakanandam, K; Lowe, KC; Power, JB; Davey, MR. Hemoglobin (ErythrogenTM)-enhanced mitotic division and plant regeneration from cultured rice protoplasts (Oryza sativa L.). Enzyme Microb. Tech 1997, 21, 572–577. [Google Scholar]
- Lamar, CA; Mahesh, VB; Brann, DW. Regulation of gonadotrophin-releasing hormone (GnRH) secretion by heme molecules: A regulatory role for carbon monoxide? Endocrinology 1996, 137, 790–793. [Google Scholar]
- Xuan, W; Zhu, FY; Xu, S; Huang, BK; Ling, TF; Qi, JY; Ye, MB; Shen, WB. The heme oxygenase/carbon monoxide system is involved in the auxin-induced cucumber adventitious rooting process. Plant Physiol 2008, 148, 881–893. [Google Scholar]
- Uchida, A; Jagendorf, AT; Hibino, T; Takabe, T; Takabe, T. Effects of hydrogen peroxide and nitric oxide on both salt and heat stress tolerance in rice. Plant Sci 2002, 63, 515–523. [Google Scholar]
- Garratt, LC; Janagoudar, BS; Anthony, P; Davey, MR; Power, JB; Lowe, KC. Hemoglobin-stimulated growth and antioxidant activities in cultured cotton cells. Free Radic. Biol. Med 2001, 31, 1156–1162. [Google Scholar]
- Xie, YJ; Ling, TF; Han, Y; Liu, KL; Zheng, QS; Huang, LQ; Yuan, XX; He, ZY; Hu, B; Fang, L; Shen, ZG; Yang, Q; Shen, WB. Carbon monoxide enhances salt tolerance by nitric oxide-mediated maintenance of ion homeostasis and up-regulation of antioxidant defense in wheat seedling roots. Plant Cell Environ 2008, 31, 1864–1881. [Google Scholar]
- Huang, BK; Xu, S; Xuan, W; Li, M; Cao, ZY; Liu, KL; Ling, TF; Shen, WB. Carbon monoxide alleviates salt-induced oxidative damage in wheat seedling leaves. J. Integr. Plant Biol 2006, 48, 249–254. [Google Scholar]
- Liu, YH; Xu, S; Ling, TF; Xu, LL; Shen, WB. Heme oxygenase/carbon monoxide system participates in regulating wheat seed germination under osmotic stress involving the nitric oxide pathway. J. Plant Physiol 2010, 167, 1371–1379. [Google Scholar]
- Wu, MZ; Huang, JJ; Xu, S; Ling, TF; Xie, YJ; Shen, WB. Haem oxygenase delays programmed cell death in wheat aleurone layers by modulation of hydrogen peroxide metabolism. J. Exp. Bot 2011, 62, 235–248. [Google Scholar]
- Sjorstrand, T. Endogenous formation of carbon monoxide in man under normal and pathological conditions. Scand. J. Clin. Lab. Invest 1949, 1, 201–214. [Google Scholar]
- Ruan, HH; Shen, WB; Ye, MB; Xu, LL. Protective effects of nitric oxide on salt stress-induced oxidative damages to wheat (Triticum aestivum) leaves. Chin. Sci. Bull 2002, 47, 677–681. [Google Scholar]
- Nakano, Y; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol 1981, 22, 867–880. [Google Scholar]
- Beauchamp, C; Fridovich, I. Superoxide dismutase: Improved assays and an assay applicable to acrylamide gels. Anal. Biochem 1971, 44, 276–287. [Google Scholar]
- Bradford, MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem 1976, 72, 248–254. [Google Scholar]
- Vázquez, MD; Poschenrieder, C; Corrales, I; Barceló, J. Change in apoplastic aluminum during the initial growth response to aluminum by roots of a tolerant maize variety. Plant Physiol 1999, 119, 435–444. [Google Scholar]
- Zhao, L; Zhang, F; Guo, J; Yang, Y; Li, B; Zhang, L. Nitric oxide functions as a signal in salt resistance in the calluses from two ecotypes of reed. Plant Physiol 2007, 144, 206–217. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Germination rate (%) | Shoot length (cm) | Root length (cm) |
|---|---|---|---|
| H2O→H2O | 89.55 ± 2.52a | 1.34 ± 0.10a | 3.44 ± 0.05a |
| Hb0.01→H2O | 88.32 ± 2.84a | 1.37 ± 0.11a | 3.39 ± 0.25a |
| Hb0.05→H2O | 90.51 ± 2.05a | 1.35 ± 0.08a | 3.35 ± 0.07a |
| Hb0.2→H2O | 95.44 ± 0.69a | 1.38 ± 0.11a | 3.34 ± 0.04a |
| Hb1.0→H2O | 96.34 ± 2.69a | 1.44 ± 0.10a | 3.27 ± 0.22a |
| Hb5.0→H2O | 94.43 ± 2.11a | 1.45 ± 0.05a | 3.05 ± 0.20b |
| H2O→S | 45.47 ± 7.26c | 0.70 ± 0.05c | 2.55 ± 0.25bc |
| Hb0.01→S | 51.48 ± 3.44c | 0.76 ± 0.10bc | 2.80 ± 0.48bc |
| Hb0.05→S | 52.86 ± 2.69c | 0.76 ± 0.10bc | 2.57 ± 0.51bc |
| Hb0.2→S | 61.05 ± 9.67b | 0.87 ± 0.10b | 2.50 ± 0.29c |
| Hb1.0→S | 67.31 ± 3.92b | 0.93 ± 0.09b | 2.47 ± 0.01c |
| Hb5.0→S | 62.98 ± 5.47b | 0.90 ± 0.03b | 2.38 ± 0.24c |
| Treatment | Germination rate (%) | Shoot length (cm) | Root length (cm) |
|---|---|---|---|
| H2O→H2O | 90.30 ± 2.38a | 1.33 ± 0.20a | 3.46 ± 0.14a |
| Hb1.0→H2O | 94.47 ± 2.49a | 1.46 ± 0.16a | 3.30 ± 0.23a |
| H2O→ZnPPIX | 87.55 ± 2.08ab | 1.15 ± 0.24ab | 2.90 ± 0.10ab |
| Hb1.0→ZnPPIX | 84.54 ± 5.05ab | 1.18 ± 0.25ab | 3.04 ± 0.30ab |
| H2O→S | 46.32 ± 6.59d | 0.70 ± 0.08c | 2.58 ± 0.33b |
| Hb1.0→S | 64.88 ± 3.77c | 0.90 ± 0.11b | 2.47 ± 0.10b |
| H2O→S + ZnPPIX | 44.19 ± 5.04d | 0.66 ± 0.07c | 2.14 ± 0.25bc |
| Hb1.0→S + ZnPPIX | 45.70 ± 2.58d | 0.79 ± 0.09bc | 1.95 ± 0.21c |
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Xu, S.; Hu, B.; He, Z.; Ma, F.; Feng, J.; Shen, W.; Yang, J. Enhancement of Salinity Tolerance during Rice Seed Germination by Presoaking with Hemoglobin. Int. J. Mol. Sci. 2011, 12, 2488-2501. https://doi.org/10.3390/ijms12042488
Xu S, Hu B, He Z, Ma F, Feng J, Shen W, Yang J. Enhancement of Salinity Tolerance during Rice Seed Germination by Presoaking with Hemoglobin. International Journal of Molecular Sciences. 2011; 12(4):2488-2501. https://doi.org/10.3390/ijms12042488
Chicago/Turabian StyleXu, Sheng, Bing Hu, Ziyi He, Fei Ma, Jianfei Feng, Wenbiao Shen, and Jie Yang. 2011. "Enhancement of Salinity Tolerance during Rice Seed Germination by Presoaking with Hemoglobin" International Journal of Molecular Sciences 12, no. 4: 2488-2501. https://doi.org/10.3390/ijms12042488