Differential Responses to Blood Pressure and Oxidative Stress in Streptozotocin-Induced Diabetic Wistar-Kyoto Rats and Spontaneously Hypertensive Rats: Effects of Antioxidant (Honey) Treatment

Abstract
:1. Introduction
2. Results and Discussion
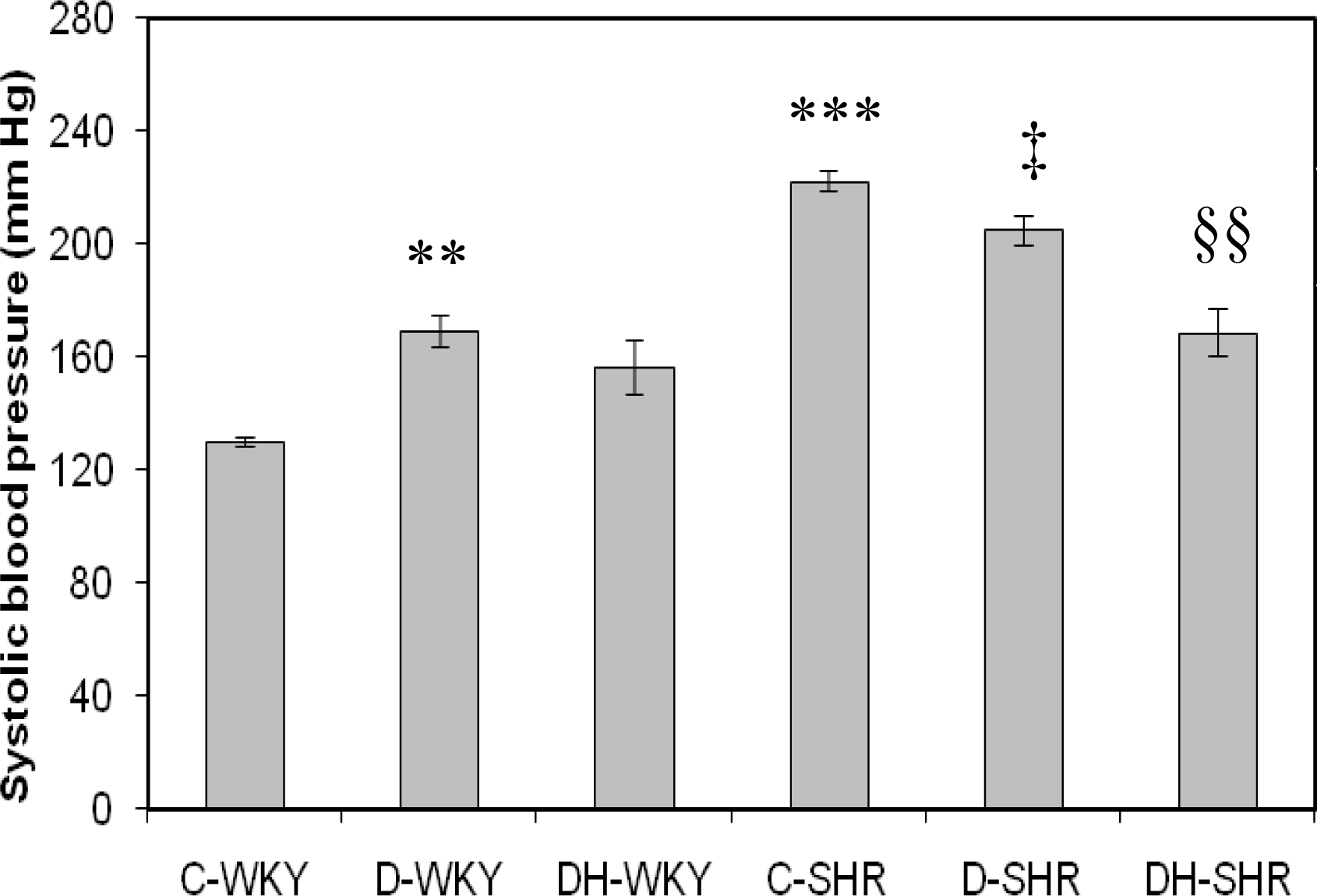
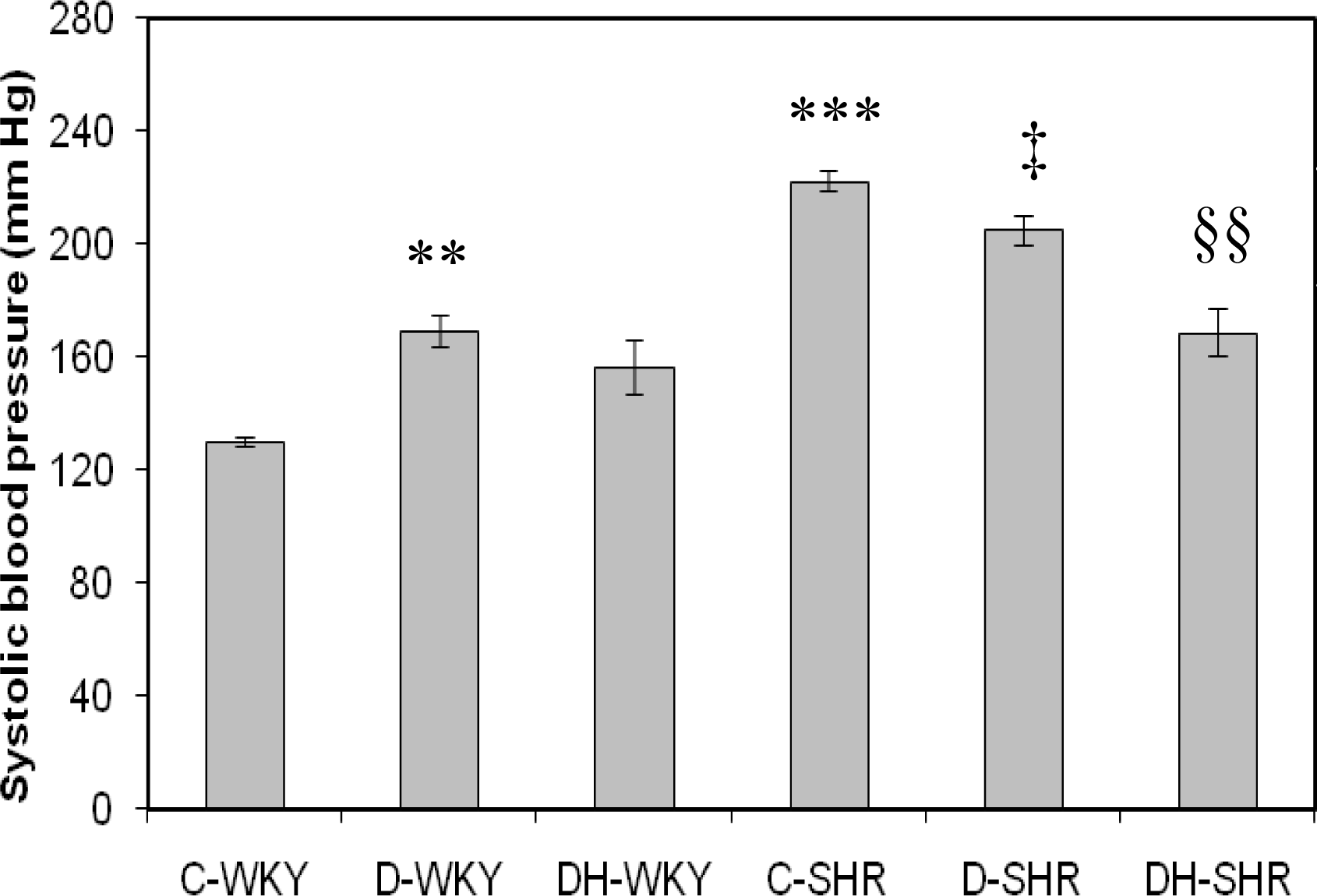
2.1. Systolic Blood Pressure
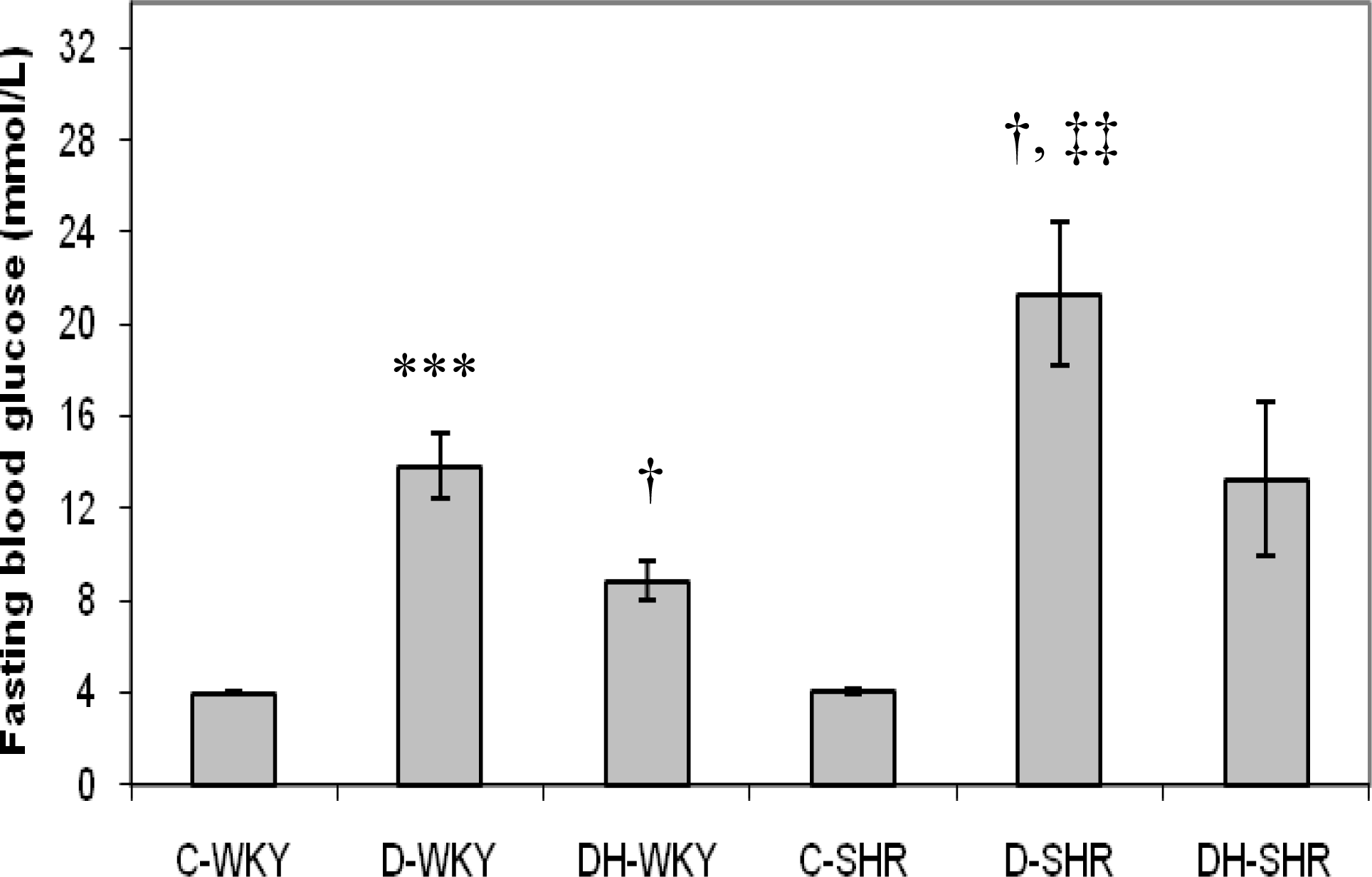
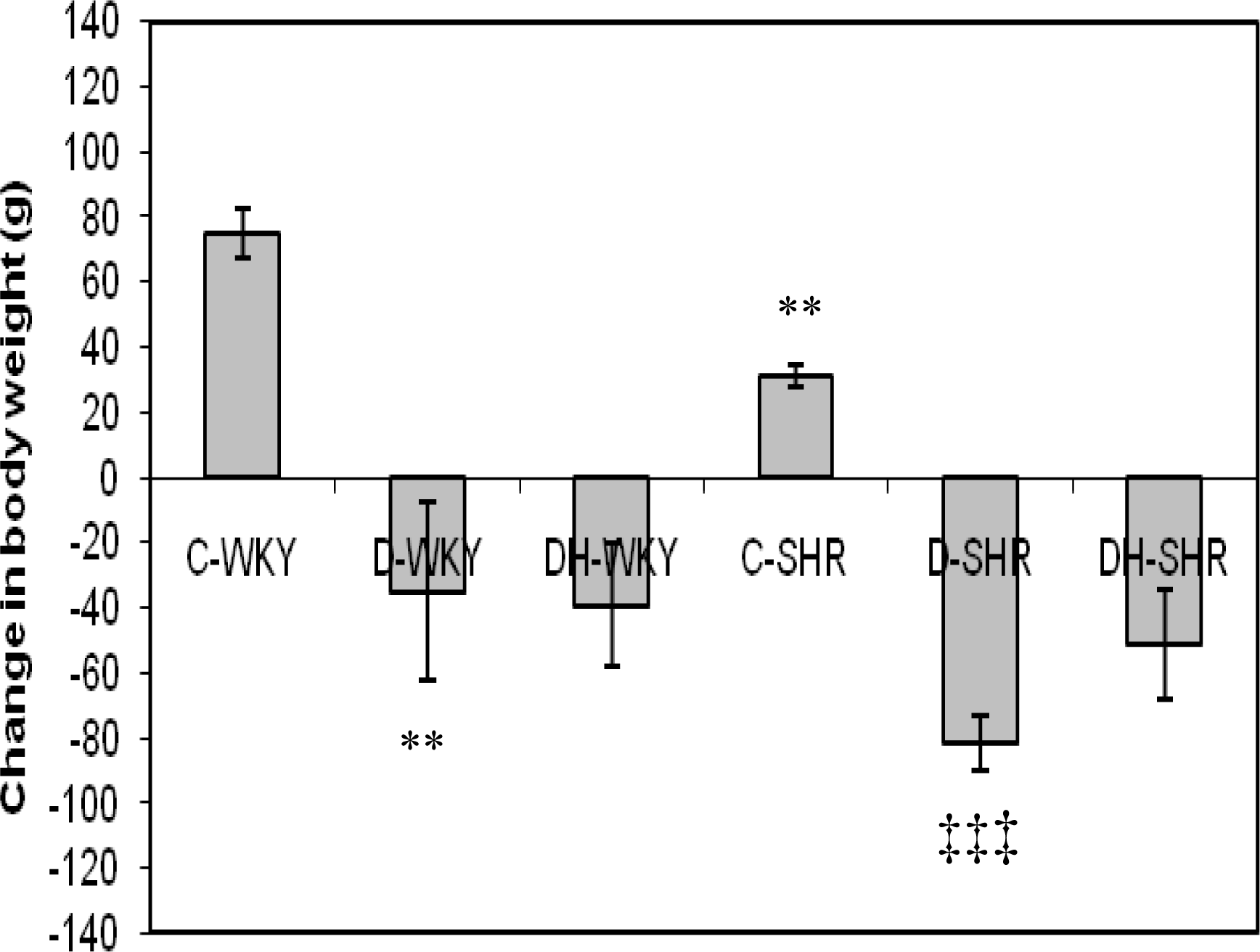
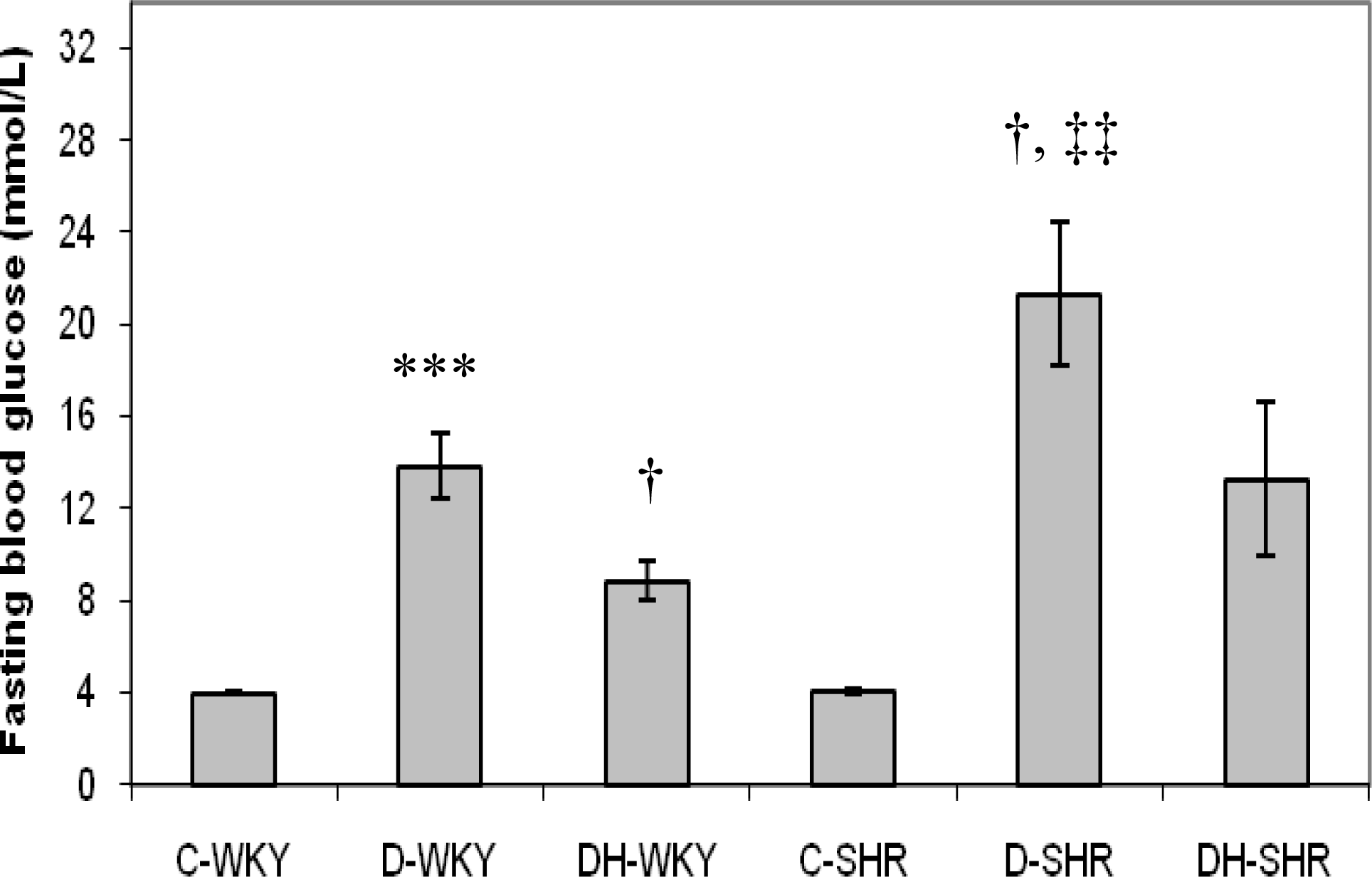
2.2. Blood Glucose and Body Weight
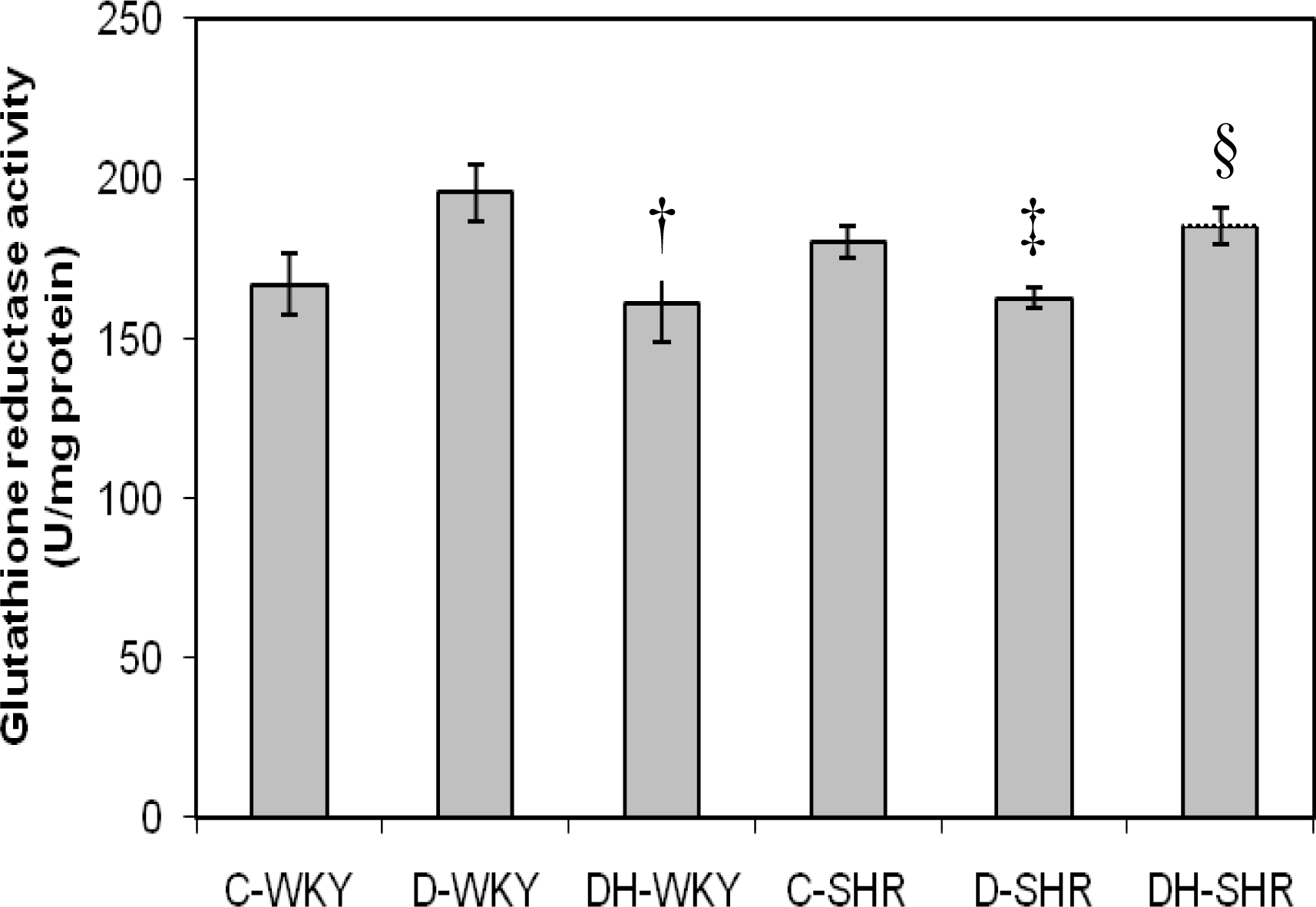
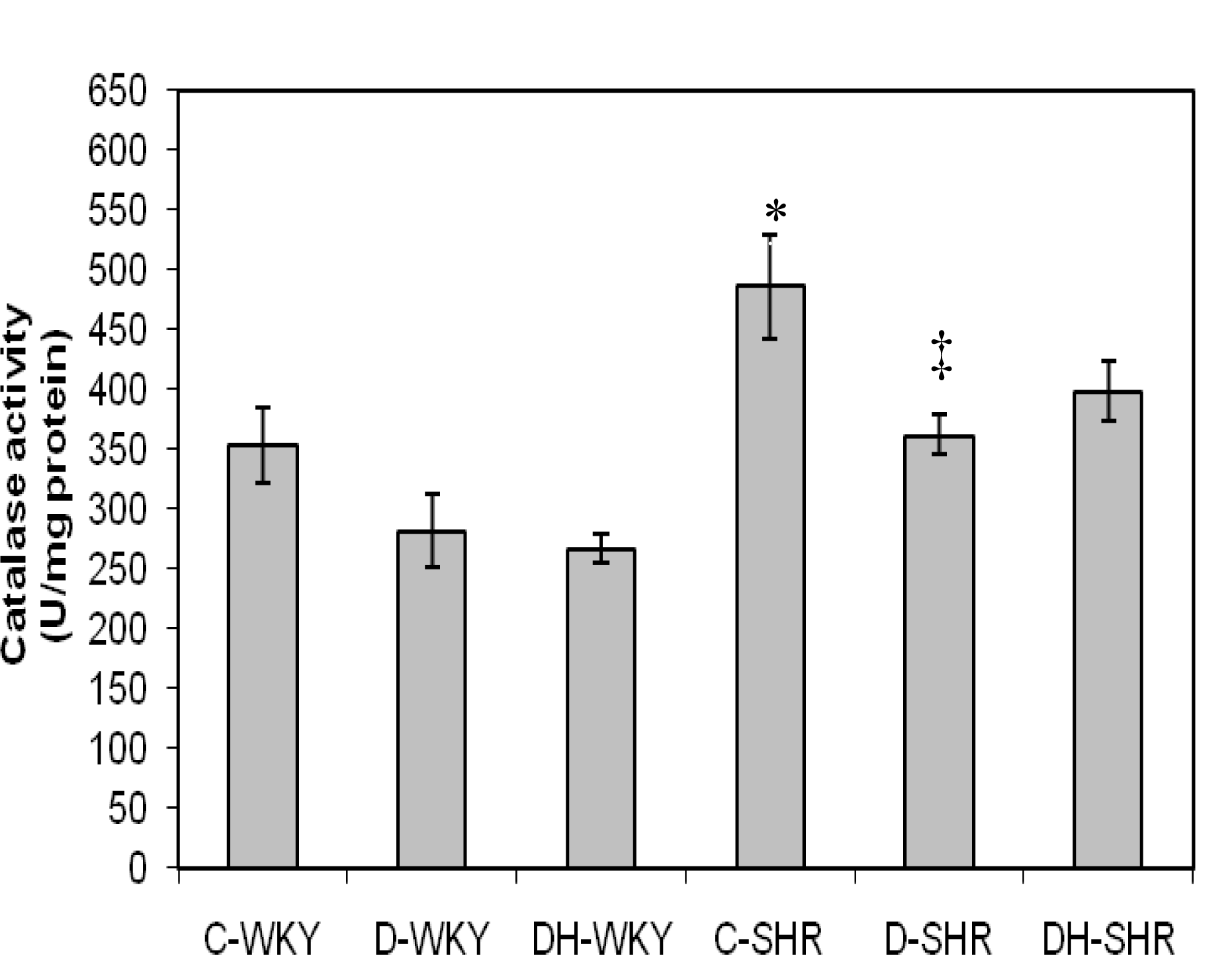
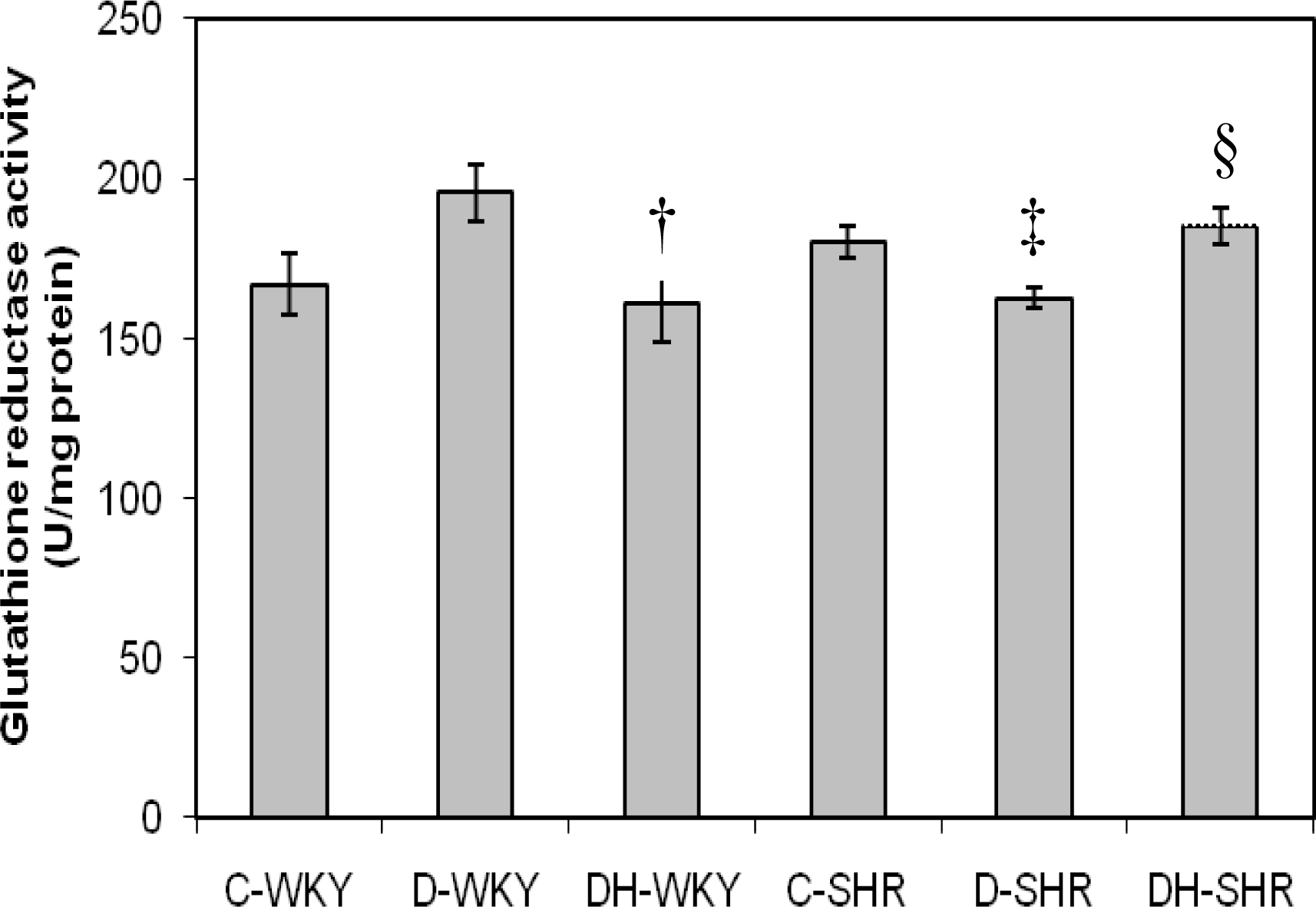
2.3. Antioxidant Enzymes
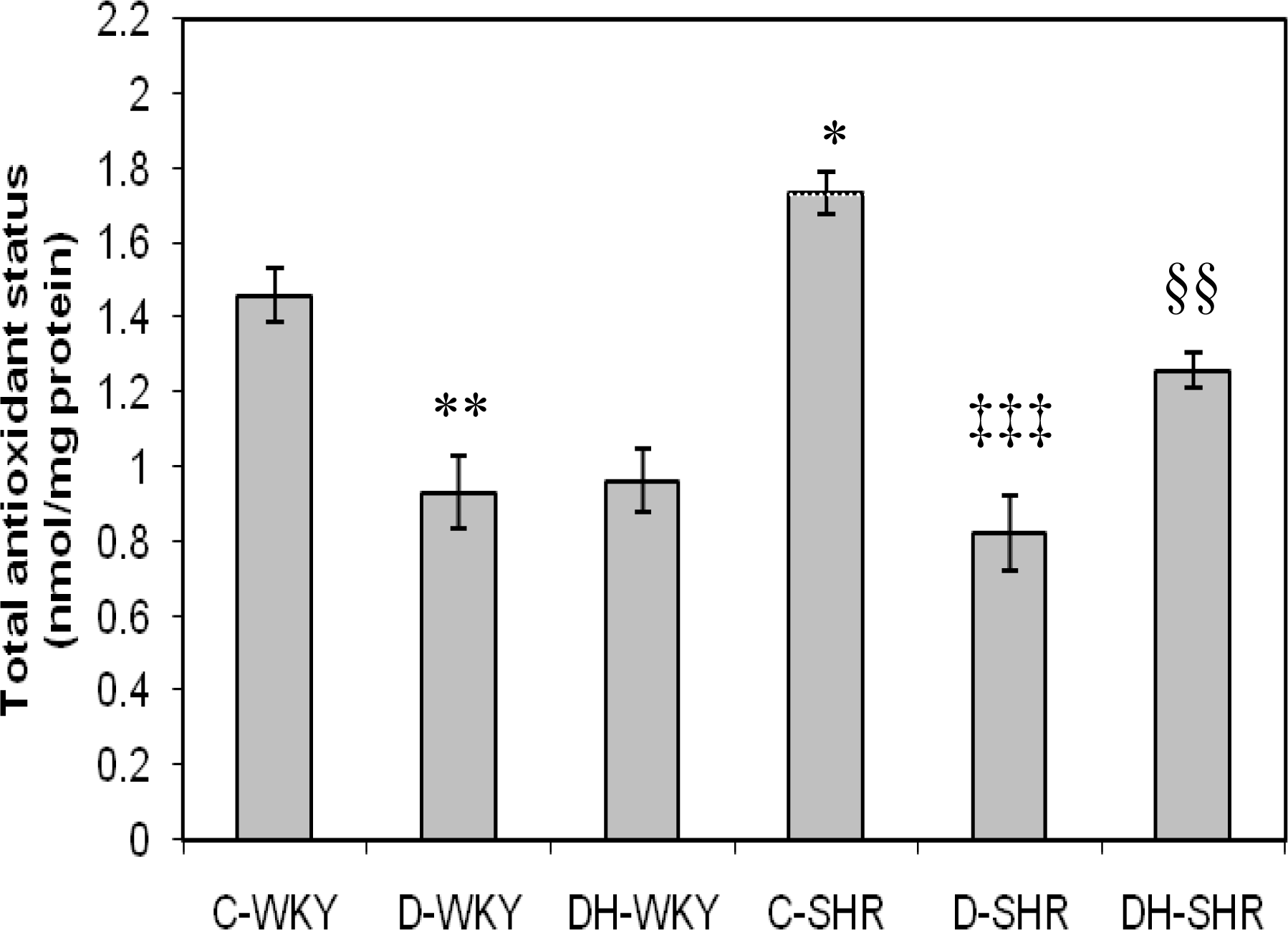
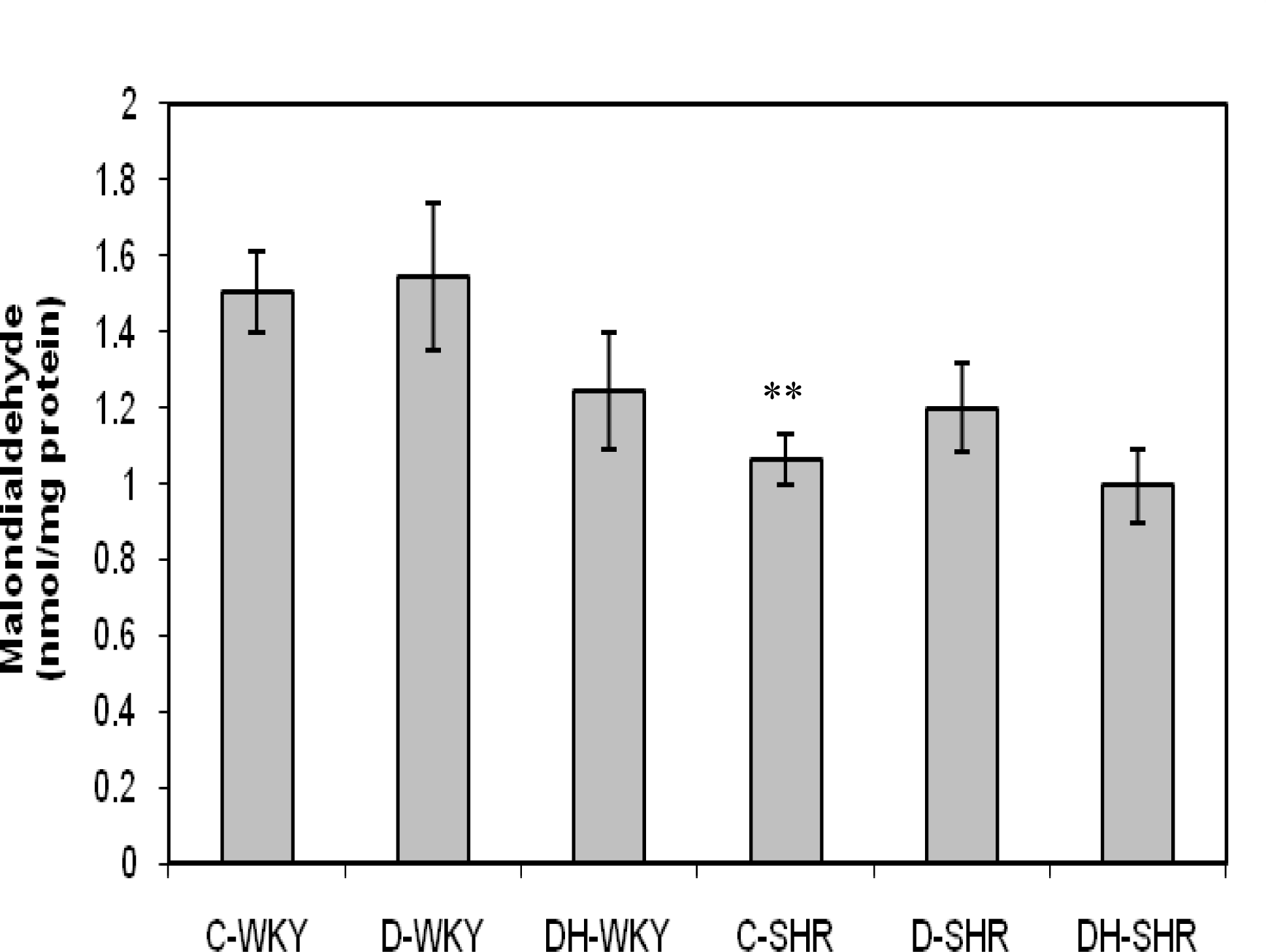
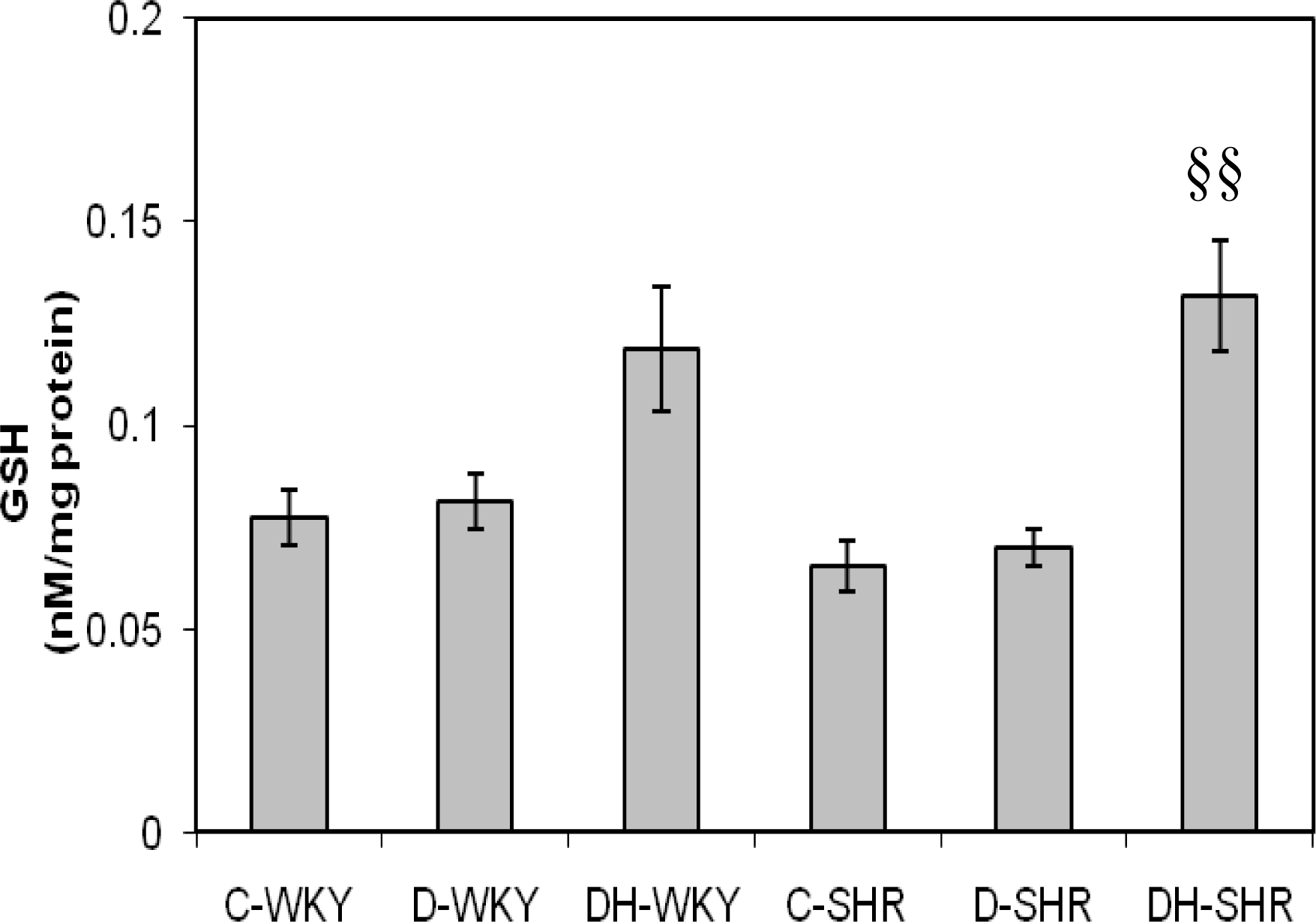
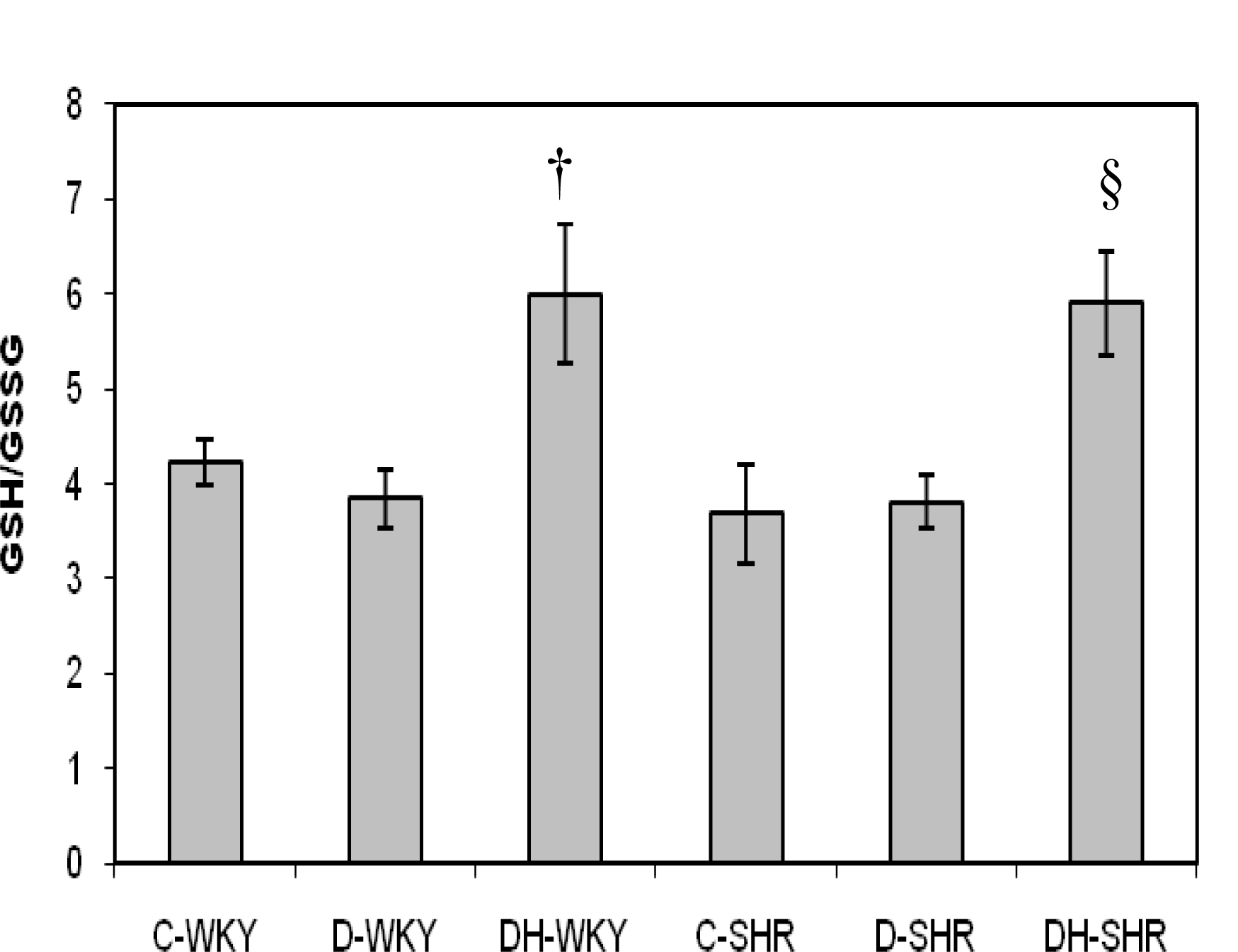
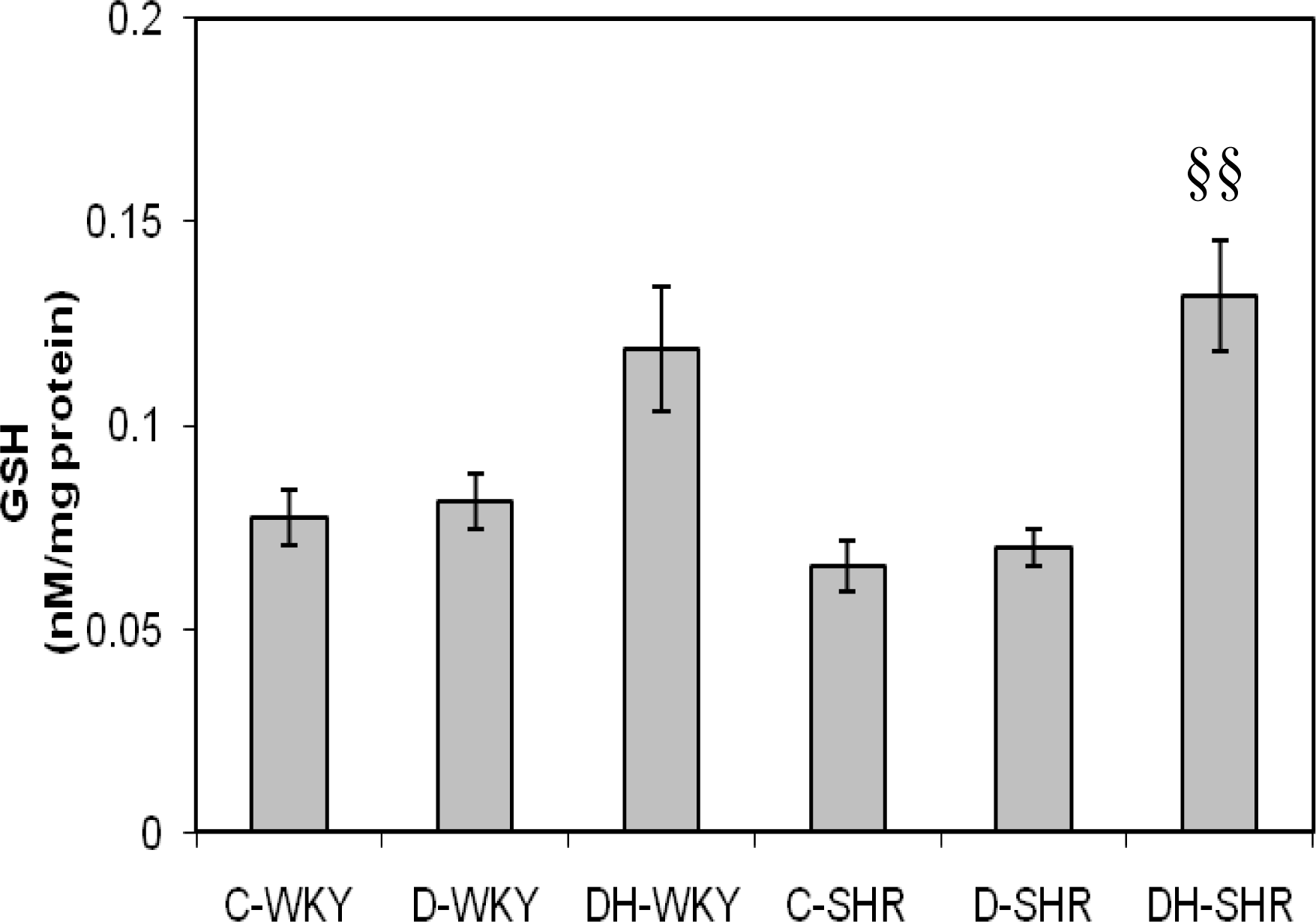
2.4. Markers of Oxidative Stress
3. Materials and Methods
3.1. Animals
3.2. Chemicals and Tualang Honey
3.3. Induction of Diabetes and Treatment
3.4. Measurement of Blood Pressure, Blood Glucose and Body Weight
3.5. Processing of Tissues and Biochemical Assays
3.6. Superoxide Dismutase (SOD) Assay
3.7. Catalase (CAT) Assay
3.8. Glutathione Peroxidase (GPx) Assay
3.9. Glutathione-S-Transferase (GST) Assay
3.10. Glutathione Reductase (GR) Assay
3.11. Total Antioxidant Status (TAS) Assay
3.12. Lipid Peroxidation Assay
3.13. Assay of Reduced and Oxidized Glutathione
3.14. Protein Assay
3.15. Statistical Analysis
4. Conclusions
Acknowledgments
References
- Kannel, WB; McGee, DL. Diabetes and cardiovascular disease: The Framingham study. JAMA 1978, 241, 2035–2038. [Google Scholar]
- Christlieb, AR. Diabetes and hypertensive vascular disease. Am. J. Cardiol 1973, 32, 592–606. [Google Scholar]
- Fein, FS; Capasso, JM; Aronson, RS; Cho, S; Nordin, C; Miller-Green, B; Sonnenblick, EH; Factor, SM. Combined renovascular hypertension and diabetes in rats: A new preparation of congestive cardiomyopathy. Circulation 1984, 70, 318–330. [Google Scholar]
- Ross, R. The pathogenesis of atherosclerosis. N. Engl. J. Med 1986, 314, 488–500. [Google Scholar]
- Reaven, GM. Pathophysiology of insulin resistance in human disease. Physiol. Rev 1995, 75, 473–486. [Google Scholar]
- Pieper, GM; Gross, GJ. Oxygen-derived free radicals abolish endothelium-dependent relaxation in diabetic rat aorta. Am. J. Physiol 1988, 255, H825–H833. [Google Scholar]
- Tesfamariam, B; Cohen, RA. Free radicals mediate endothelial cell dysfunction caused by elevated glucose. Am. J. Physiol 1992, 263, H321–H326. [Google Scholar]
- Ohara, Y; Petersen, TE; Harrison, DG. Hypercholesterolemia increases endothelial O2− anion production. J. Clin. Invest 1993, 91, 2546–2551. [Google Scholar]
- Marui, N; Offerman, MK; Swerlick, R; Kunsch, C; Rosen, CA; Ahmad, M; Alexander, RW; Medford, RM. Vascular cell adhesion molecule-1 (VCAM-1) gene transcription and expression are regulated through an antioxidant-sensitive mechanism in human vascular endothelial cells. J. Clin. Invest 1993, 92, 1866–1874. [Google Scholar]
- Laight, DW; Desai, KM; Anggard, EE; Carrier, MJ. Endothelial dysfunction accompanies a pro-oxidant, pro-diabetic challenge in the insulin resistant, obese Zucker rat in vivo. Eur. J. Pharmacol 2000, 402, 95–99. [Google Scholar]
- Weber, C; Erl, W; Pietsch, A; Strobel, M; Ziegler-Heitbrock, HWL; Weber, PC. Antioxidants inhibit monocyte adhesion by suppressing nuclear factor-κB mobilization and induction of vascular cell adhesion molecule-1 in endothelial cells stimulated to generate radicals. Arterioscler. Thromb 1994, 14, 1665–1673. [Google Scholar]
- Friedman, J; Peleg, E; Kagan, T; Shnizer, S; Rosenthal, T. Oxidative stress in hypertensive, diabetic, and diabetic hypertensive rats. Am. J. Hypertens 2003, 16, 1049–1052. [Google Scholar]
- Tan, HT; Rahman, RA; Gan, SH; Halim, AS; Hassan, SA; Sulaiman, SA; Kirnpal-Kaur, B. The antibacterial properties of Malaysian tualang honey against wound and enteric microorganisms in comparison to manuka honey. BMC Complement. Altern. Med 2009, 9, 1–8. [Google Scholar]
- Ghashm, AA; Othman, NH; Khattak, MN; Ismail, NM; Saini, R. Antiproliferative effect of tualang honey on oral squamous cell carcinoma and osteosarcoma cell lines. BMC Complement. Altern. Med 2010, 10, 49. [Google Scholar]
- Mohamed, M; Sirajudeen, KNS; Swamy, M; Yaacob, NS; Sulaiman, SA. Studies on the antioxidant properties of tualang honey of Malaysia. Afr. J. Trad. CAM 2010, 7, 59–63. [Google Scholar]
- Erejuwa, OO; Gurtu, S; Sulaiman, SA; Ab Wahab, MS; Sirajudeen, KNS; Salleh, MS. Hypoglycemic and antioxidant effects of honey supplementation in streptozotocin-induced diabetic rats. Int. J. Vitam. Nutr. Res 2010, 80, 74–82. [Google Scholar]
- Erejuwa, OO; Sulaiman, SA; Ab Wahab, MS; Sirajudeen, KNS; Salzihan, MS; Gurtu, S. Antioxidant protective effect of glibenclamide and metformin in combination with honey in pancreas of streptozotocin-induced diabetic rats. Int. J. Mol. Sci 2010, 11, 2056–2066. [Google Scholar]
- Erejuwa, OO; Sulaiman, SA; Ab Wahab, MS; Sirajudeen, KNS; Salzihan, MS; Gurtu, S. Comparison of antioxidant effects of honey, glibenclamide, metformin, and their combinations in kidney of streptozotocin-induced diabetic rats. Int. J. Mol. Sci 2011, 12, 829–843. [Google Scholar]
- Trippodo, NC; Frolich, ED. The genetically hypertensive rat as a model for human essential hypertension. Circ. Res 1981, 48, 309–319. [Google Scholar]
- Rakieten, N; Rakieten, ML; Nadkarni, MV. Studies on the diabetogenic action of streptozotocin (NSC-37917). Cancer Chemother. Rep 1963, 29, 91–98. [Google Scholar]
- Davidoff, AJ; Pinault, FM; Rodgers, RL. Ventricular relaxation of diabetic spontaneously hypertensive rats. Hypertension 1990, 15, 643–651. [Google Scholar]
- Erejuwa, OO; Sulaiman, SA; Ab Wahab, MS; Sirajudeen, KNS; Salzihan, MS; Gurtu, S. Universiti Sains Malaysia, Kubang Kerian, Kelantan. Unpublished work. 2010.
- Roos, MD; Wen, X; Kaihong, S; Clark, JA; Xiaoyong, Y; Chin, E; Paterson, AJ; Kudlow, JE. Streptozotocin, an analog of N-acetylglucosamine, blocks the removal of O-GlcNAc from intracellular proteins. Proc. Assoc. Am. Phys 1998, 110, 1–11. [Google Scholar]
- Suzuki, H; Swei, A; Zweifach, BW; Schmid-Schonbein, GW. In vivo evidence for microvascular oxidative stress in spontaneously hypertensive rats: Hydroethidine microfluorography. Hypertension 1995, 25, 1083–1089. [Google Scholar]
- Haluzik, M; Nedvidkova, J. The role of nitric oxide in the development of streptozotocin-induced diabetes mellitus: experimental and clinical implication. Physiol. Res 2000, 49, 37–42. [Google Scholar]
- Takasu, N; Komiya, I; Asawa, T; Nagasawa, Y; Yamada, T. Streptozotocin- and alloxan-induced H2O2 generation and DNA fragmentation in pancreatic islets. H2O2 as mediator for DNA fragmentation. Diabetes 1991, 40, 1141–1145. [Google Scholar]
- Pamela, CC; Richard, AH. Biochemistry; J.B. Lippincott Co: Philadelphia, PA, USA, 1994; pp. 269–280. [Google Scholar]
- Stearns, SB; Tepperman, HM; Tepperman, J. Studies on the utilization and mobilization of lipid in skeletal muscles from streptozotocin-diabetic and control rats. J. Lipid Res 1979, 20, 654–662. [Google Scholar]
- Factor, SM; Bhan, R; Minase, T; Wolinsky, H; Sonnenblick, EH. Hypertensive-diabetic cardiomyopathy in the rat. An experimental model of human disease. Am. J. Pathol 1981, 102, 219–228. [Google Scholar]
- Touyz, RM; Schiffrin, EL. Oxidative stress in arterial hypertension: oxidative stress and hypertension. In Atherosclerosis and Oxidant Stress—A New Perspective; Holtzman, J, Ed.; Springer: New York, NY, USA, 2008; pp. 51–78. [Google Scholar]
- Bunag, RD; Tomita, T; Sasaki, S. Streptozotocin diabetic rats are hypertensive despite reduced hypothalamic responsiveness. Hypertension 1982, 4, 556–565. [Google Scholar]
- Sharma, JN; Kesavarao, U. Cardiac kallikrein in hypertensive and normotensive rats with and without diabetes. Immunopharmacology 1996, 33, 341–343. [Google Scholar]
- To mLinson, KC; Gardiner, SM; Bennett, T. Blood pressure in streptozotocin-treated Brattleboro and Long Evans rats. Am. J. Physiol 1990, 258, R852–R859. [Google Scholar]
- Rodgers, RL. Depressor effect of diabetes in the spontaneously hypertensive rat: Associated changes in heart performance. Can. J. Physiol. Pharmacol 1986, 64, 1177–1184. [Google Scholar]
- Kawashima, H; Igarashi, T; Nakajima, Y; Akiyama, Y; Usuki, K; Ohtake, S. Chronic hypertension induced by streptozotocin in rats. Naunyn-Schmiedebergs Arch. Pharmacol 1978, 305, 123–126. [Google Scholar]
- Susic, D; Mandal, AK; Jovovic, DJ; Radujkovic, G; Kentera, D. Streptozotocin-induced diabetes mellitus lowers blood pressure in spontaneously hypertensive rat. Clin. Exp. Hypertens. A 1990, 12, 1021–1035. [Google Scholar]
- Kelly, DJ; Wilkinson-Berka, JL; Allen, TJ; Cooper, ME; Skinner, SL. A new model of diabetic nephropathy with progressive renal impairment in the transgenic (mRen-2)27 rat (TGR). Kidney Int 1998, 54, 343–352. [Google Scholar]
- Davidoff, AJ; Rodgers, RL. Insulin, thyroid hormone, and heart function of diabetic spontaneously hypertensive rat. Hypertension 1990, 15, 633–642. [Google Scholar]
- Medeiros, FJ; Aguila, MB; Mandarim-de-Lacerda, CA. Renal cortex remodeling in streptozotocin-induced diabetic spontaneously hypertensive rats treated with olive oil, palm oil and fish oil from Menhaden. Prostaglandins Leukot. Essent. Fatty Acids 2006, 75, 357–365. [Google Scholar]
- Moreno, JJ; Carbonell, T; Sánchez, T; Miret, S; Mitjavila, MT. Olive oil decreases both oxidative stress and the production of arachidonic acid metabolites by the prostaglandin G/H synthase pathway in rat macrophages1. J. Nutr 2001, 131, 2145–2149. [Google Scholar]
- Nyby, MD; Abedi, K; Eslami, P; Hernandez, G; Smutko, V; Berger, ME; Tuck, ML. Dietary fish oil prevents hypertension, oxidative stress and suppression of endothelial nitric oxide synthase expression in fructose-fed rats. Am. J. Hypertens 2004, 17, 216. [Google Scholar]
- Ge, X; Yu, Q; Qi, W; Shi, X; Zhai, Q. Chronic insulin treatment causes insulin resistance in 3T3-L1 adipocytes through oxidative stress. Free Radic. Res 2008, 42, 582–591. [Google Scholar]
- Tiedge, M; Lortz, S; Munday, R; Lenzen, S. Protection against the co-operative toxicity of nitric oxide and oxygen free radicals by over-expression of antioxidant enzymes in bioengineered insulin-producing RI nm5F cells. Diabetologia 1999, 42, 849–855. [Google Scholar]
- Wolff, SP; Dean, RT. Glucose autoxidation and protein modification: The potential role of autoxidative glycosylation in diabetes. Biochem. J 1987, 245, 243–250. [Google Scholar]
- Pigeolet, E; Corbisier, P; Houbion, A; Lambert, D; Michiels, C; Raes, M; Zachary, MD; Remacle, J. Glutathione peroxidase, superoxide dismutase and catalase inactivation by peroxides and oxygen derived free radicals. Mech. Ageing Dev 1990, 51, 283–297. [Google Scholar]
- Sartori-Valinotti, JC; Iliescu, R; Fortepiani, L; Yanes, L; Reckelhoff, J. Sex differences in oxidative stress and the impact on blood pressure control and cardiovascular disease. Clin. Exp. Pharm. Physiol 2007, 34, 938–945. [Google Scholar]
- Maritim, AC; Sanders, RA; Watkins, JB. Diabetes, oxidative stress and antioxidants: A review. J. Biochem. Mol. Toxicol 2003, 17, 24–38. [Google Scholar]
- Newaz, MA; Nawal, NN. Effect of alpha-tocopherol on lipid peroxidation and total antioxidant status in spontaneously hypertensive rats. Am. J. Hypertens 1998, 11, 1480–1485. [Google Scholar]
- Mantle, D; Patel, VB; Why, HJ; Ahmed, S; Rahman, I; MacNee, W; Wassif, WS; Richardson, PJ; Preedy, VR. Effects of lisinopril and a mLodipine on antioxidant status in experimental hypertension. Clin. Chim. Acta 2000, 299, 1–10. [Google Scholar]
- Young, IS. Measurement of total antioxidant capacity. J. Clin. Pathol 2001, 54, 339–340. [Google Scholar]
- Jackson, P; Loughrey, CM; Lightbody, JH; McNamee, PT; Young, IS. Effect of hemodialysis on total antioxidant capacity and serum antioxidants in patients with chronic renal failure. Clin. Chem 1995, 41, 1135–1138. [Google Scholar]
- Nabha, L; Garbern, JC; Buller, CL; Charpie, JR. Vascular oxidative stress precedes high blood pressure in spontaneously hypertensive rats. Clin. Exp. Hypertens 2005, 27, 71–82. [Google Scholar]
- Gott, L. A simple method for determination of serum catalase activity and revision of reference range. Clin. Chim. Acta 1991, 196, 143–151. [Google Scholar]
- Habig, WH; Pabst, MJ; Jakoby, WB. Glutathione-S-transferases. The first enzymatic step in mercapturic acid formation. J. Biol. Chem 1974, 249, 7130–7139. [Google Scholar]
- Goldberg, DM; Spooner, RJ. Assay of glutathione reductase. In Methods of Enzymatic Analysis; Bergmeyen, HV, Ed.; Verlag Chemie: Weinheim, Germany, 1983; pp. 258–265. [Google Scholar]
- Koracevic, D; Koracevic, G; Djordjevic, V; Andrejevic, S; Cosic, V. Method for the measurement of antioxidant activity in human fluids. J. Clin. Pathol 2000, 54, 356–361. [Google Scholar]
- Ohkawa, H; Ohishi, N; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem 1979, 95, 351–358. [Google Scholar]
- Bradford, MA. Rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem 1976, 72, 248–254. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group Treatment |
|---|
| Non-diabetic WKY Distilled water (0.5 mL) |
| Diabetic WKY Distilled water (0.5 mL) |
| Diabetic WKY Tualang honey (1.0 g/kg/body weight) |
| Non-diabetic SHR Distilled water (0.5 mL) |
| Diabetic SHR Distilled water (0.5 mL) |
| Diabetic SHR Tualang honey (1.0 g/kg/body weight) |
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Erejuwa, O.O.; Sulaiman, S.A.; Wahab, M.S.A.; Sirajudeen, K.N.S.; Salleh, M.S.M.; Gurtu, S. Differential Responses to Blood Pressure and Oxidative Stress in Streptozotocin-Induced Diabetic Wistar-Kyoto Rats and Spontaneously Hypertensive Rats: Effects of Antioxidant (Honey) Treatment. Int. J. Mol. Sci. 2011, 12, 1888-1907. https://doi.org/10.3390/ijms12031888
Erejuwa OO, Sulaiman SA, Wahab MSA, Sirajudeen KNS, Salleh MSM, Gurtu S. Differential Responses to Blood Pressure and Oxidative Stress in Streptozotocin-Induced Diabetic Wistar-Kyoto Rats and Spontaneously Hypertensive Rats: Effects of Antioxidant (Honey) Treatment. International Journal of Molecular Sciences. 2011; 12(3):1888-1907. https://doi.org/10.3390/ijms12031888
Chicago/Turabian StyleErejuwa, Omotayo O., Siti A. Sulaiman, Mohd Suhaimi Ab Wahab, Kuttulebbai N. S. Sirajudeen, Md Salzihan Md Salleh, and Sunil Gurtu. 2011. "Differential Responses to Blood Pressure and Oxidative Stress in Streptozotocin-Induced Diabetic Wistar-Kyoto Rats and Spontaneously Hypertensive Rats: Effects of Antioxidant (Honey) Treatment" International Journal of Molecular Sciences 12, no. 3: 1888-1907. https://doi.org/10.3390/ijms12031888