Effects of Intermediates between Vitamins K2 and K3 on Mammalian DNA Polymerase Inhibition and Anti-Inflammatory Activity

Abstract
:1. Introduction
2. Results
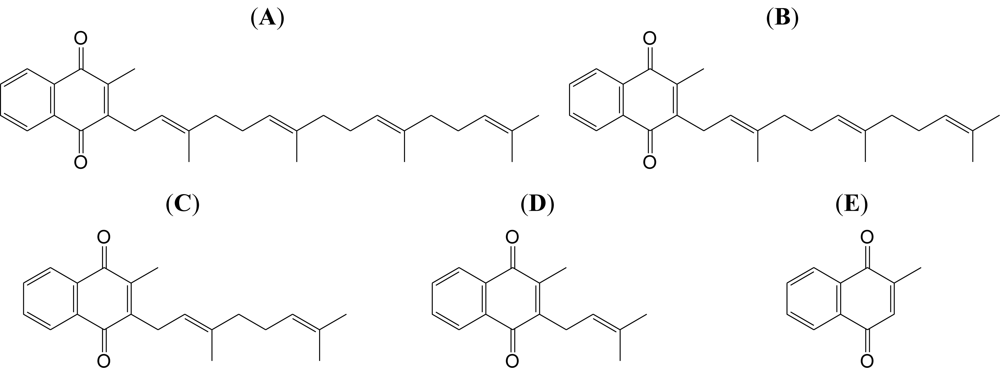
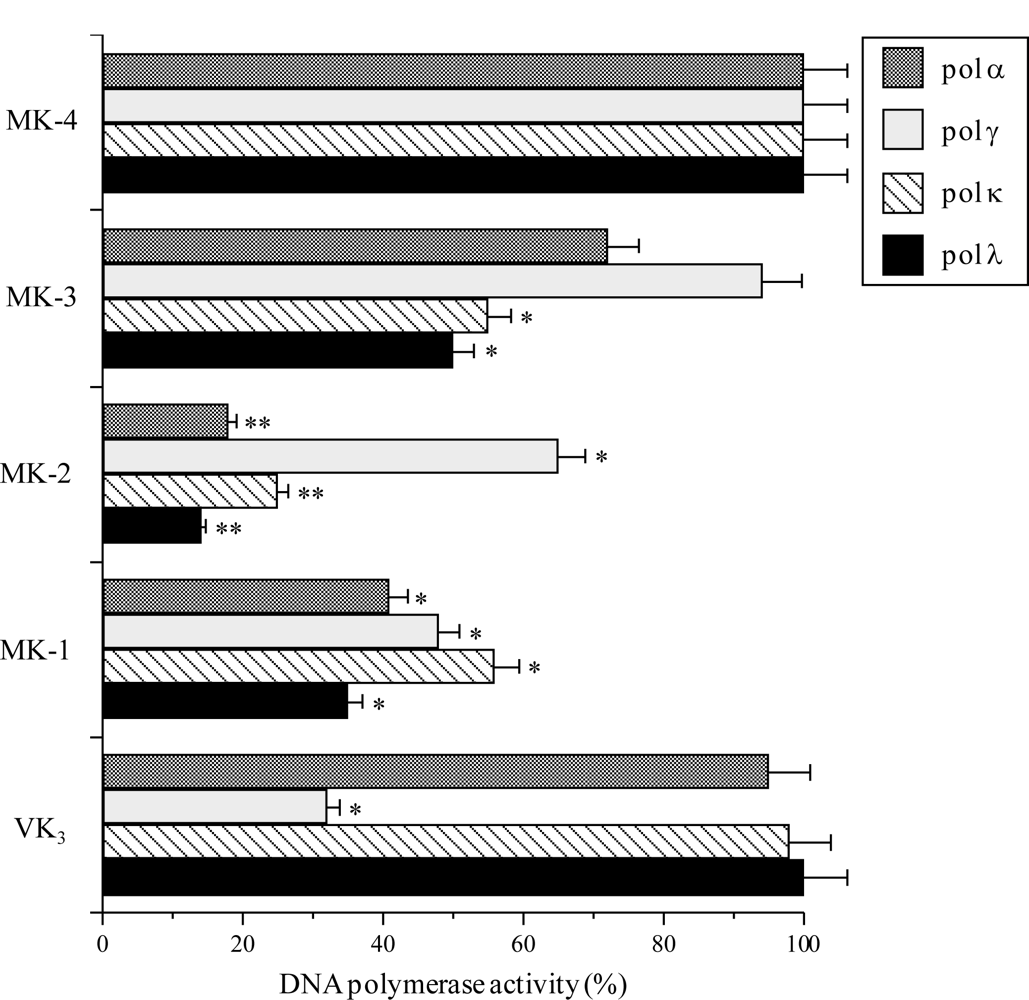
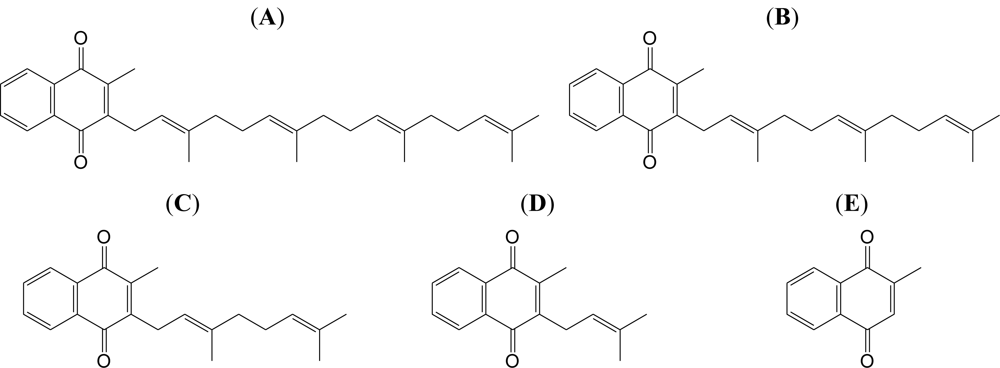
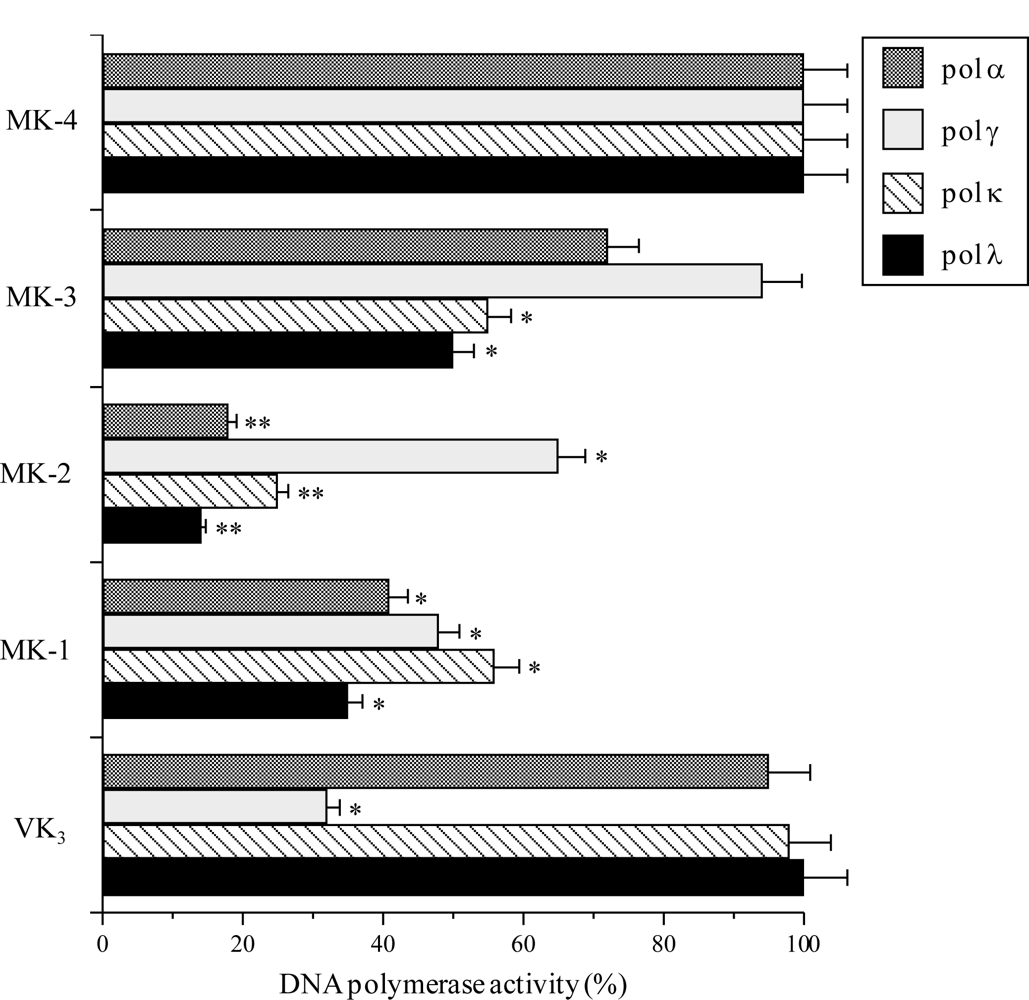
2.1. Effect of the VK2 and VK3 Intermediates on Mammalian Pol Activity
2.2. Effects of MK-2 on Pols and Other DNA Metabolic Enzymes
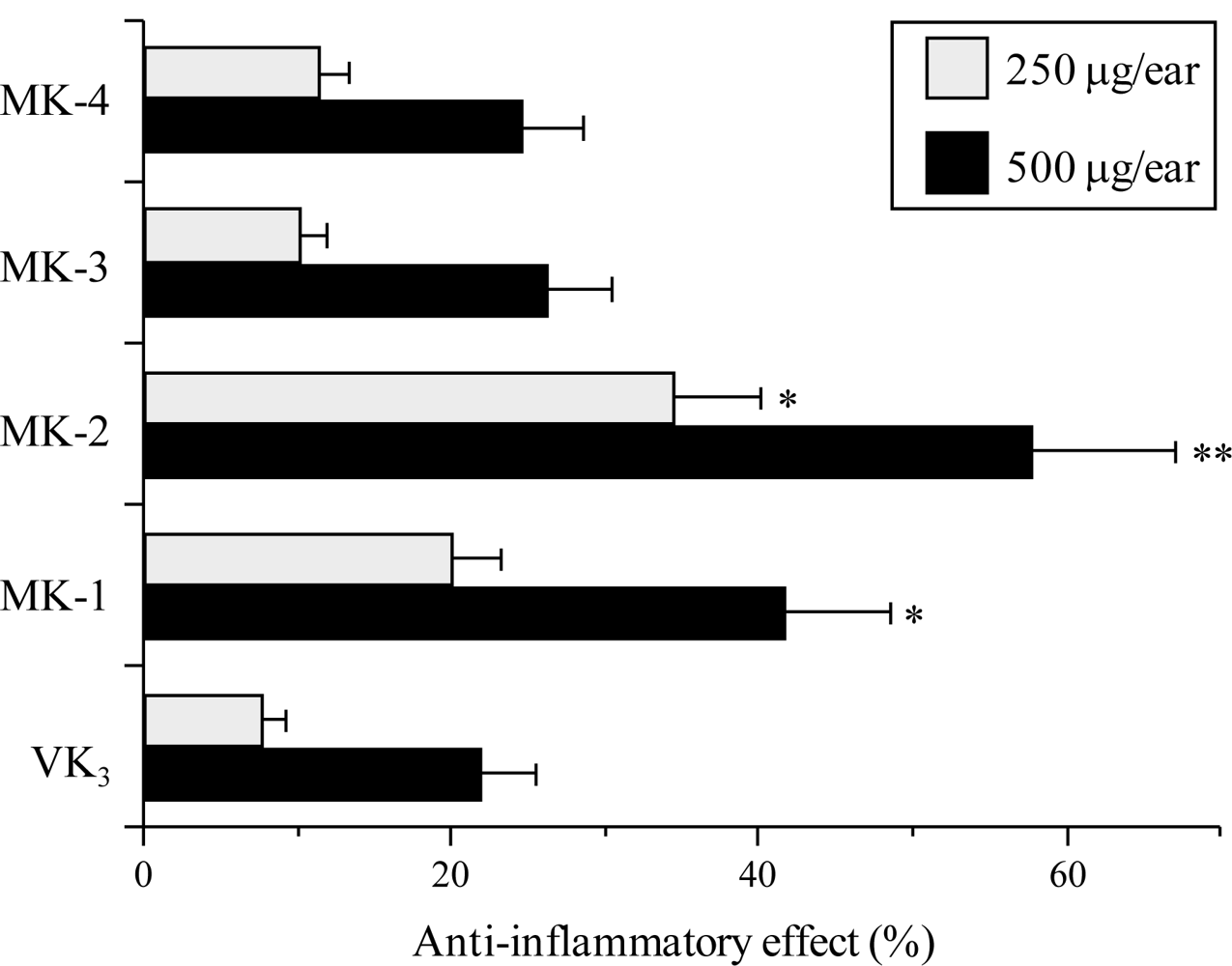
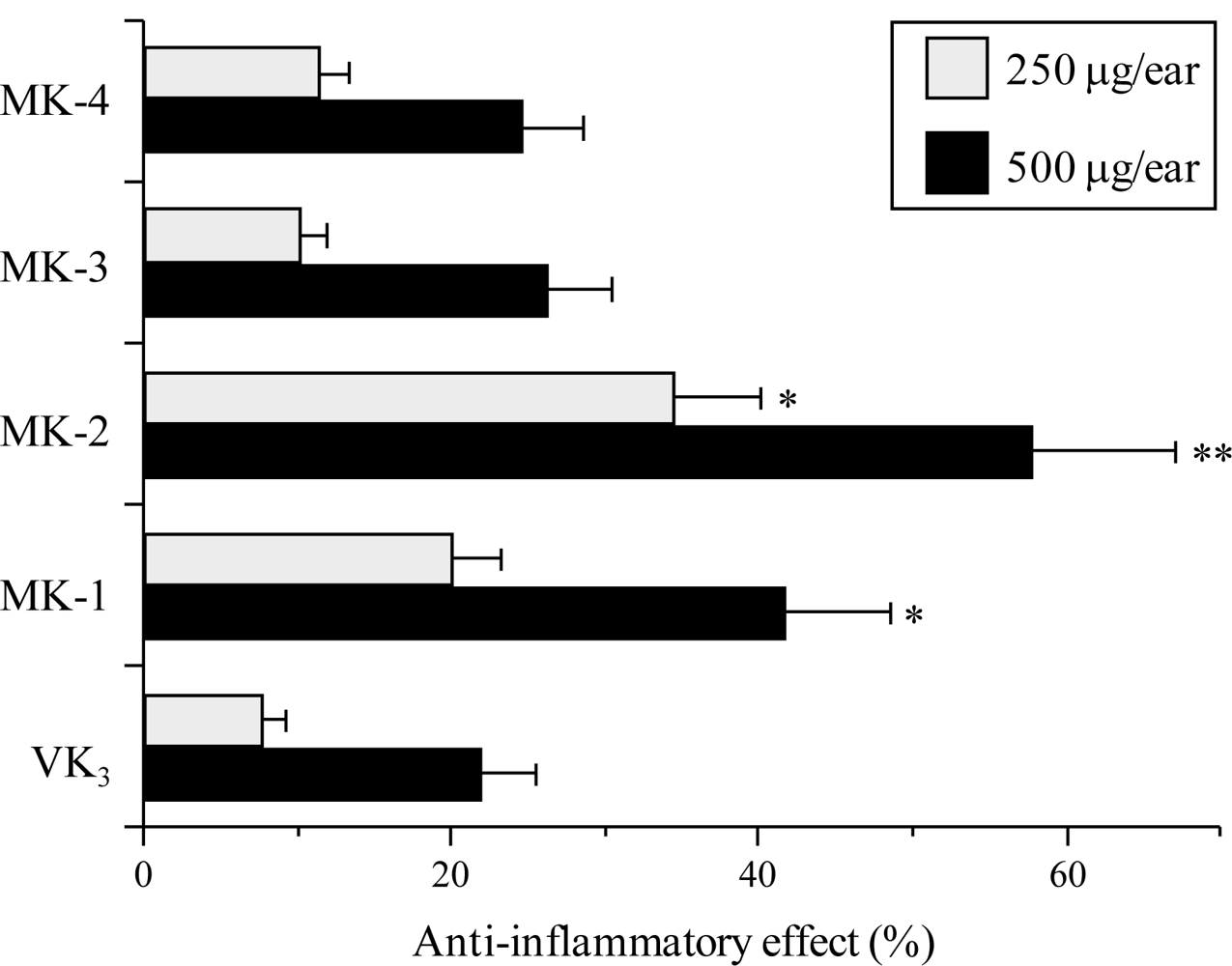
2.3. Effect of the VK2 and VK3 Intermediates on TPA-Induced Anti-Inflammatory Activity
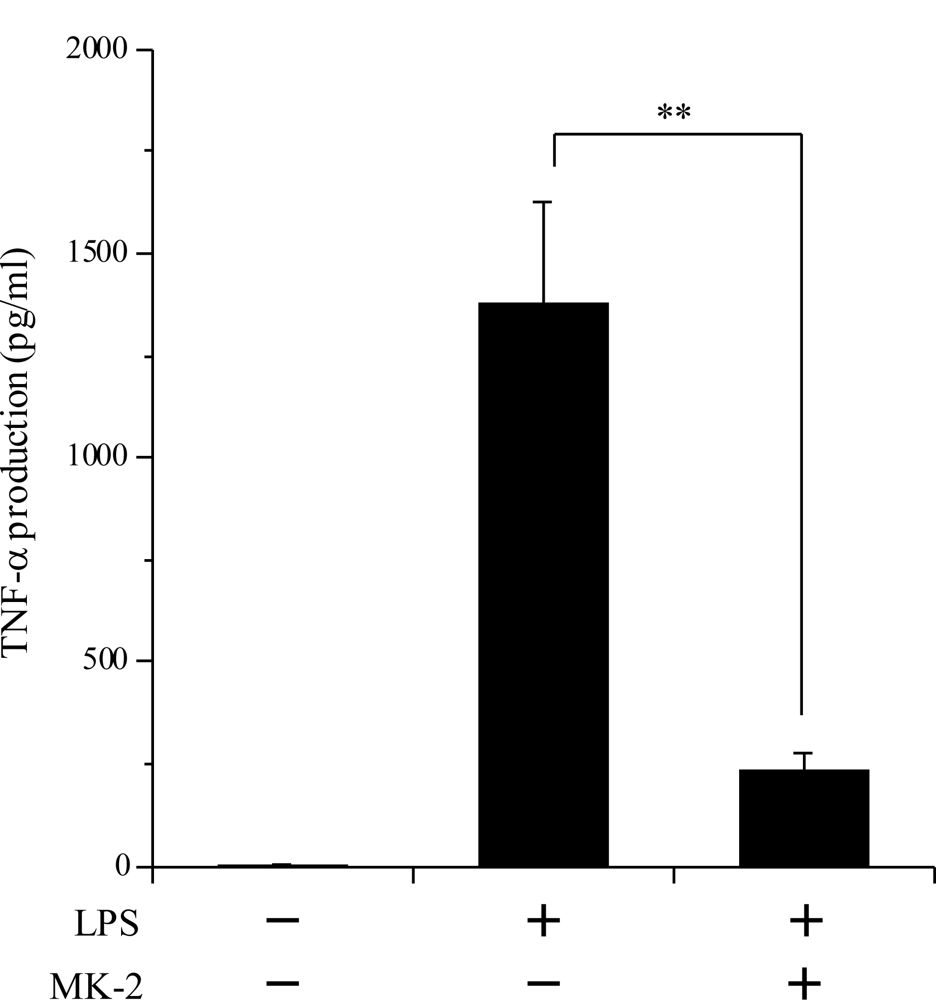
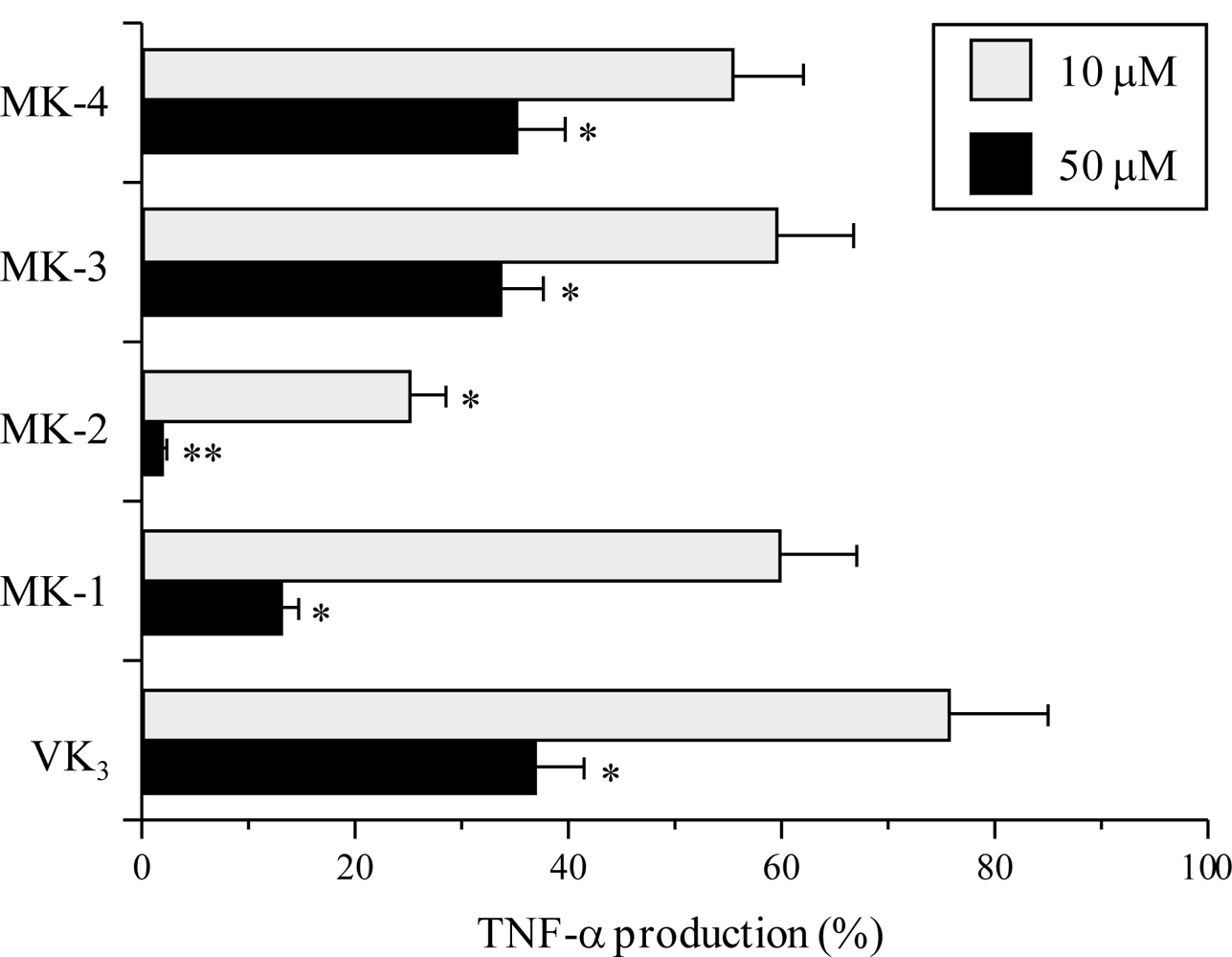
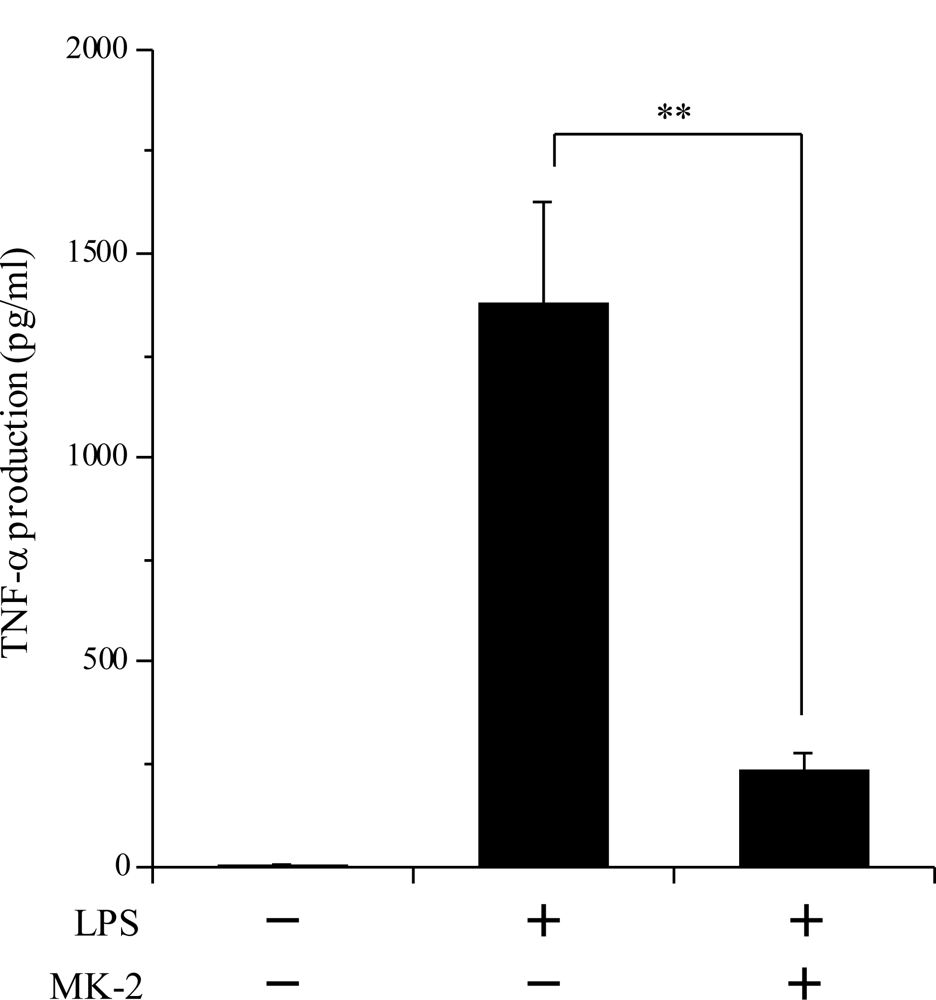
2.4. Inhibitory Effect of the VK2 and VK3 Intermediates on the LPS-Induced Inflammatory Response in Cultured Macrophage Cells
2.5. Inhibitory Effect of MK-2 on LPS-Induced Inflammation in Vivo
3. Discussion
4. Experimental Section
4.1. Materials
4.2. Preparation of the VK2 and VK3 Intermediates
4.3. Pol and Other DNA Metabolic Enzyme Assays
4.4. Animal Experiments
4.5. TPA-Induced Anti-Inflammatory Assay in Mouse
4.6. Measurement of TNF-α Level in the Cell Culture Medium of Mouse Macrophages
4.7. Cell Treatment and Preparation of Nuclear Proteins
4.8. Western Blotting
4.9. In Vivo LPS-Induced Inflammatory Experiment
4.10. Statistical Analysis
5. Conclusions
Acknowledgments
References
- Hubscher, U; Maga, G; Spadari, S. Eukaryotic DNA polymerases. Annu. Rev. Biochem 2002, 71, 133–163. [Google Scholar]
- Bebenek, K; Kunkel, TA. DNA repair and replication. In Advances in Protein Chemistry; Yang, W, Ed.; Elsevier: San Diego, CA, USA, 2004; Volume 69, pp. 137–165. [Google Scholar]
- Takata, K; Shimizu, T; Iwai, S; Wood, RD. Human DNA polymerase N (POLN) is a low fidelity enzyme capable of error-free bypass of 5S-thymine glycol. J. Biol. Chem 2006, 281, 23445–23455. [Google Scholar]
- Friedberg, EC; Feaver, WJ; Gerlach, VL. The many faces of DNA polymerases: strategies for mutagenesis and for mutational avoidance. Proc. Natl. Acad. Sci. USA 2000, 97, 5681–5683. [Google Scholar]
- Sakaguchi, K; Sugawara, F; Mizushina, Y. Inhibitors of eukaryotic DNA polymerases. Seikagaku 2002, 74, 244–251. [Google Scholar]
- Mizushina, Y. Specific inhibitors of mammalian DNA polymerase species. Biosci. Biotechnol. Biochem 2009, 73, 1239–1251. [Google Scholar]
- Sasaki, R; Suzuki, Y; Yonezawa, Y; Ota, Y; Okamoto, Y; Demizu, Y; Huang, P; Yoshida, H; Sugimura, K; Mizushina, Y. DNA polymerase γ inhibition by vitamin K3 induces mitochondria-mediated cytotoxicity in human cancer cells. Cancer Sci 2008, 99, 1040–1048. [Google Scholar]
- Matsubara, K; Kayashima, T; Mori, M; Yoshida, H; Mizushina, Y. Inhibitory effects of vitamin K3 on DNA polymerase and angiogenesis. Int. J. Mol. Med 2008, 22, 381–387. [Google Scholar]
- Tanaka, S; Nishiumi, S; Nishida, M; Mizushina, Y; Kobayashi, K; Masuda, A; Fujita, T; Morita, Y; Mizuno, S; Kutsumi, H; Azuma, T; Yoshida, M. Vitamin K3 attenuates lipopolysaccharide-induced acute lung injury through inhibition of nuclear factor-κB activation. Clin. Exp. Immunol 2010, 160, 283–292. [Google Scholar]
- Chinzei, R; Masuda, A; Nishiumi, S; Nishida, M; Onoyama, M; Sanki, T; Fujita, T; Moritoh, S; Itoh, T; Kutsumi, H; Mizuno, S; Azuma, T; Yoshida, M. Vitamin K3 attenuates cerulein-induced acute pancreatitis through inhibition of the autophagic pathway. Pancreas 2010, 40, 84–94. [Google Scholar]
- Seegers, WH; Bang, NU. Blood Clotting Enzymology; Academic Press: New York, NY, USA, 1967. [Google Scholar]
- Billeter, M; Bolliger, W; Martius, C. Untersuchungen uber die umwandlung von verfutterten K-vitamin durch austausch der seitenkette und die rolle der darmbakterien hierbei. Biochem. Z 1964, 340, 290–303. [Google Scholar]
- Davidson, RT; Foley, AL; Engelke, JA; Suttie, JW. Conversion of dietary phylloquinone to tissue menaquinone-4 in rats is not dependent on gut bacteria. J. Nutr 1998, 128, 220–223. [Google Scholar]
- The Merck Index, Budavari, S; O’Neil, MJ; Smith, A; Heckelman, PE. Merck & Co. Inc.: Rahway, NJ, USA, 1989.
- Taggart, WV; Matschiner, JT. Metabolism of menadione-6,7-3H in the rat. Biochemistry 1969, 8, 1141–1146. [Google Scholar]
- Mizushina, Y; Kamisuki, S; Kasai, N; Ishidoh, T; Shimazaki, N; Takemura, M; Asahara, H; Linn, S; Yoshida, S; Koiwai, O; Sugawara, F; Yoshida, H; Sakaguchi, K. Petasiphenol: a DNA polymerase λ inhibitor. Biochemistry 2002, 41, 14463–14471. [Google Scholar]
- Mizushina, Y; Ishidoh, T; Takeuchi, T; Shimazaki, N; Koiwai, O; Kuramochi, K; Kobayashi, S; Sugawara, F; Sakaguchi, K; Yoshida, H. Monoacetylcurcumin: a new inhibitor of eukaryotic DNA polymerase λ and a new ligand for inhibitor-affinity chromatography. Biochem. Biophys. Res. Commun 2005, 337, 1288–1295. [Google Scholar]
- Takeuchi, T; Ishidoh, T; Iijima, H; Kuriyama, I; Shimazaki, N; Koiwai, O; Kuramochi, K; Kobayashi, S; Sugawara, F; Sakaguchi, K; Yoshida, H; Mizushina, Y. Structural relationship of curcumin derivatives binding to the BRCT domain of human DNA polymerase λ. Genes Cells 2006, 11, 223–235. [Google Scholar]
- Mizushina, Y; Hirota, M; Murakami, C; Ishidoh, T; Kamisuki, S; Shimazaki, N; Takemura, M; Perpelescu, M; Suzuki, M; Yoshida, H; Sugawara, F; Koiwai, O; Sakaguchi, K. Some anti-chronic inflammatory compounds are DNA polymerase λ-specific inhibitors. Biochem. Pharmacol 2003, 66, 1935–1944. [Google Scholar]
- Mizushina, Y; Takeuchi, T; Kuramochi, K; Kobayashi, S; Sugawara, F; Sakaguchi, K; Yoshida, H. Study on the molecular structure and bio-activity (DNA polymerase inhibitory activity, anti-inflammatory activity and anti-oxidant activity) relationship of curcumin derivatives. Curr. Bioactive Compd 2007, 3, 171–177. [Google Scholar]
- Nishida, M; Nishiumi, S; Mizushina, Y; Fujishima, Y; Yamamoto, K; Masuda, A; Mizuno, S; Fujita, T; Morita, Y; Kutsumi, H; Yoshida, H; Azuma, T; Yoshida, M. Monoacetylcurcumin strongly regulates inflammatory responses through inhibition of NF-κB activation. Int. J. Mol. Med 2010, 25, 761–767. [Google Scholar]
- Hecker, E. Carcinogenesis; Raven Press: New York, NY, USA, 1978; pp. 11–48. [Google Scholar]
- Fujiki, H; Sugimura, T. Advances in Cancer Research; Academic Press Inc: London, UK, 1987; pp. 223–264. [Google Scholar]
- Nakamura, Y; Murakami, A; Ohto, Y; Torikai, K; Tanaka, T; Ohigashi, H. Suppression of tumor promoter-induced oxidative stress and inflammatory responses in mouse skin by a superoxide generation inhibitor 1′-acetoxychavicol acetate. Cancer Res 1995, 58, 4832–4839. [Google Scholar]
- Huang, TT; Wuerzberger-Davis, SM. Sequential modification of NEMO/IKKÁ by SUMO-1 and ubiquitin mediates NF-κB activation by genotoxic stress. Cell 2003, 115, 565–576. [Google Scholar]
- Hayden, MS; Ghosh, S. Signaling to NF-κB. Genes Dev 2004, 18, 2195–2224. [Google Scholar]
- Bonizzi, G; Karin, M. The two NF-κB activation pathways and their role in innate and adaptive immunity. Trends Immunol 2004, 25, 280–288. [Google Scholar]
- Wajant, H; Pfizenmaier, K; Scheurich, P. Tumor necrosis factor signaling. Cell Death Differ 2003, 10, 45–65. [Google Scholar]
- Elson, CO; Sartor, RB; Tennyson, GS; Riddell, RH. Experimental models of inflammatory bowel disease. Gastroenterology 1995, 109, 1344–1367. [Google Scholar]
- Aggarwal, BB. Signalling pathways of the TNF superfamily: A double-edged sword. Nat. Rev. Immunol 2003, 3, 745–756. [Google Scholar]
- Furie, B; Furie, BC. Molecular basis of vitamin K-dependent gamma-carboxylation. Blood 1990, 75, 1753–1762. [Google Scholar]
- Suttie, JW. Synthesis of vitamin K-dependent proteins. FASEB J 1993, 7, 445–452. [Google Scholar]
- Shearer, MJ. Role of vitamin K and Gla proteins in the pathophysiology of osteoporosis and vascular calcification. Curr. Opin. Clin. Nutr. Metab. Care 2000, 3, 433–438. [Google Scholar]
- Ohsaki, Y; Shirakawa, H; Hiwatashi, K; Furukawa, Y; Mizutani, T; Komai, M. Vitamin K suppresses lipopolysaccharide-induced inflammation in the rat. Biosci. Biotechnol. Biochem 2006, 70, 926–932. [Google Scholar]
- Elder, SJ; Haytowitz, DB; Howe, J; Peterson, JW; Booth, SL. Vitamin K contents of meat, dairy, and fast food in the U.S. Diet. J. Agric. Food Chem 2006, 54, 463–467. [Google Scholar]
- Tsukamoto, Y; Ichise, H; Kakuda, H; Yamaguchi, M. Intake of fermented soybean (natto) increases circulating vitamin K2 (menaquinone-7) and gamma-carboxylated osteocalcin concentration in normal individuals. J. Bone Miner. Metab 2000, 18, 216–222. [Google Scholar]
- Suttie, JW. The importance of menaquinone in human nutrition. Annu. Rev. Nutr 1995, 15, 399–417. [Google Scholar]
- Booth, SL. Dietary vitamin K guidance: An effective strategy for stable control of oral anticoagulation? Nutr. Rev 2010, 68, 178–181. [Google Scholar]
- Garcia-Diaz, M; Bebenek, K; Sabariegos, R; Dominguez, O; Rodriguez, J; Kirchhoff, T; Garcia-Palomero, E; Picher, AJ; Juarez, R; Ruiz, JF; Kunkel, TA; Blanco, L. DNA polymerase λ, a novel DNA repair enzyme in human cells. J. Biol. Chem 2002, 277, 13184–13191. [Google Scholar]
- Singhal, RK; Wilson, SH. Short gap-filling synthesis by DNA polymerase β is processive. J. Biol. Chem 1993, 268, 15906–15911. [Google Scholar]
- Matsumoto, Y; Kim, K. Excision of deoxyribose phosphate residues by DNA polymerase β during DNA repair. Science 1995, 269, 699–702. [Google Scholar]
- Sobol, RW; Horton, JK; Kuhn, R; Gu, H; Singhal, RK; Prasad, R; Rajewsky, K; Wilson, SH. Requirement of mammalian DNA polymerase-β in base-excision repair. Nature 1996, 379, 183–186. [Google Scholar]
- Ramadan, K; Shevelev, IV; Maga, G; Hubscher, U. DNA polymerase λ from calf thymus preferentially replicates damaged DNA. J. Biol. Chem 2002, 277, 18454–18458. [Google Scholar]
- Sugo, N; Aratani, Y; Nagashima, Y; Kubota, Y; Koyama, H. Neonatal lethality with abnormal neurogenesis in mice deficient in DNA polymerase β. EMBO J 2000, 19, 1397–1404. [Google Scholar]
- Garcia-Diaz, M; Bebenek, K; Kunkel, TA; Blanco, L. Identification of an intrinsic 5′-deoxyribose-5-phosphate lyase activity in human DNA polymerase λ: A possible role in base excision repair. J. Biol. Chem 2001, 276, 34659–34663. [Google Scholar]
- Hirose, F; Hotta, Y; Yamaguchi, M; Matsukage, A. Difference in the expression level of DNA polymerase β among mouse tissues: high expression in the pachytene spermatocyte. Exp. Cell Res 1989, 181, 169–180. [Google Scholar]
- Garcia-Diaz, M; Dominguez, O; Lopez-Fernandez, LA; De Lera, LT; Saniger, ML; Ruiz, JF; Parraga, M; Garcia-Ortiz, MJ; Kirchhoff, T; Del Mazo, J; Bernad, A; Blanco, L. DNA polymerase λ, a novel DNA repair enzyme in human cells. J. Mol. Biol 2000, 301, 851–867. [Google Scholar]
- Bertocci, B; de Smet, A; Flatter, E; Dahan, A; Bories, JC; Landreau, C; Weill, JC; Reynaud, CA. Cutting edge: DNA polymerases μ and λ are dispensable for Ig gene hypermutation. J. Immunol 2002, 168, 3702–3706. [Google Scholar]
- Mayer, H; Isler, O. Synthesis of vitamins K. Methods Enzymol 1971, 18, 491–547. [Google Scholar]
- Mizushina, Y; Motoshima, H; Yamaguchi, Y; Takeuchi, T; Hirano, K; Sugawara, F; Yoshida, H. 3-O-methylfunicone, a selective inhibitor of mammalian Y-family DNA polymerases from an Australian sea salt fungal strain. Mar. Drugs 2009, 7, 624–639. [Google Scholar]
- Mizushina, Y; Tanaka, N; Yagi, H; Kurosawa, T; Onoue, M; Seto, H; Horie, T; Aoyagi, N; Yamaoka, M; Matsukage, A; Yoshida, S; Sakaguchi, K. Fatty acids selectively inhibit eukaryotic DNA polymerase activities in vitro. Biochim. Biophys. Acta 1996, 1308, 256–262. [Google Scholar]
- Mizushina, Y; Yoshida, S; Matsukage, A; Sakaguchi, K. The inhibitory action of fatty acids on DNA polymerase β. Biochim. Biophys. Acta 1997, 1336, 509–521. [Google Scholar]
- Gschwendt, M; Kittstein, W; Furstenberger, G; Marks, F. The mouse ear edema: a quantitatively evaluable assay for tumor promoting compounds and for inhibitors of tumor promotion. Cancer Lett 1984, 25, 177–185. [Google Scholar]
- Medeiros, R; Otuki, MF; Avellar, MC; Calixto, JB. Mechanisms underlying the inhibitory actions of the pentacyclic triterpene α-amyrin in the mouse skin inflammation induced by phorbol ester 12-O-tetradecanoylphorbol-13-acetate. Eur. J. Pharmacol 2007, 559, 227–235. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzyme | IC50 Value of MK-2 (μM) |
|---|---|
| • Mammalian DNA Polymerases | |
| A-family DNA polymerase | |
| Human DNA polymerase γ | 68.8 ± 3.4 |
| B-family DNA polymerase | |
| Calf DNA polymerase α | 27.6 ± 1.6 |
| Human DNA polymerase δ | 29.1 ± 1.8 |
| Human DNA polymerase ɛ | 28.2 ± 1.6 |
| X-family DNA polymerase | |
| Rat DNA polymerase β | 31.0 ± 1.8 |
| Human DNA polymerase λ | 24.6 ± 1.4 |
| Calf Terminal deoxynucleotidyl transferase | 29.4 ± 1.7 |
| Y-family DNA polymerase | |
| Human DNA polymerase η | 37.8 ± 2.2 |
| Mouse DNA polymerase ι | 39.0 ± 2.3 |
| Human DNA polymerase κ | 35.3 ± 2.1 |
| • Fish DNA Polymerase | |
| Cherry salmon DNA polymerase δ | 32.5 ± 1.9 |
| • Insect DNA Polymerases | |
| Fruit fly DNA polymerase α | 34.0 ± 2.0 |
| Fruit fly DNA polymerase δ | 36.9 ± 2.2 |
| Fruit fly DNA polymerase ɛ | 36.7 ± 2.1 |
| • Plant DNA Polymerase | |
| Cauliflower DNA polymerase α | >200 |
| • Prokaryotic DNA Polymerases | |
| E. coli DNA polymerase I | >200 |
| Taq DNA polymerase | >200 |
| T4 DNA polymerase | >200 |
| • Other DNA Metabolic Enzymes | |
| Calf primase of DNA polymerase α | >200 |
| T7 RNA polymerase | >200 |
| T4 polynucleotide kinase | >200 |
| Bovine deoxyribonuclease I | >200 |
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Mizushina, Y.; Maeda, J.; Irino, Y.; Nishida, M.; Nishiumi, S.; Kondo, Y.; Nishio, K.; Kuramochi, K.; Tsubaki, K.; Kuriyama, I.; et al. Effects of Intermediates between Vitamins K2 and K3 on Mammalian DNA Polymerase Inhibition and Anti-Inflammatory Activity. Int. J. Mol. Sci. 2011, 12, 1115-1132. https://doi.org/10.3390/ijms12021115
Mizushina Y, Maeda J, Irino Y, Nishida M, Nishiumi S, Kondo Y, Nishio K, Kuramochi K, Tsubaki K, Kuriyama I, et al. Effects of Intermediates between Vitamins K2 and K3 on Mammalian DNA Polymerase Inhibition and Anti-Inflammatory Activity. International Journal of Molecular Sciences. 2011; 12(2):1115-1132. https://doi.org/10.3390/ijms12021115
Chicago/Turabian StyleMizushina, Yoshiyuki, Jun Maeda, Yasuhiro Irino, Masayuki Nishida, Shin Nishiumi, Yasuyuki Kondo, Kazuyuki Nishio, Kouji Kuramochi, Kazunori Tsubaki, Isoko Kuriyama, and et al. 2011. "Effects of Intermediates between Vitamins K2 and K3 on Mammalian DNA Polymerase Inhibition and Anti-Inflammatory Activity" International Journal of Molecular Sciences 12, no. 2: 1115-1132. https://doi.org/10.3390/ijms12021115
APA StyleMizushina, Y., Maeda, J., Irino, Y., Nishida, M., Nishiumi, S., Kondo, Y., Nishio, K., Kuramochi, K., Tsubaki, K., Kuriyama, I., Azuma, T., Yoshida, H., & Yoshida, M. (2011). Effects of Intermediates between Vitamins K2 and K3 on Mammalian DNA Polymerase Inhibition and Anti-Inflammatory Activity. International Journal of Molecular Sciences, 12(2), 1115-1132. https://doi.org/10.3390/ijms12021115