Pilot Study: Alteration of Deleted in Liver Cancer1 and Phosphorylated Focal Adhesion Kinase Y397 Cytoplasmic Expression and the Prognostic Value in Advanced Epithelial Ovarian Carcinoma

Abstract
:Background
Methods
Results
Conclusions
1. Introduction
2. Materials and Methods
2.1. Patients
2.2. Sample Selection and Grouping
2.3. Immunohistochemical Staining
2.4. Statistical Analysis
3. Results
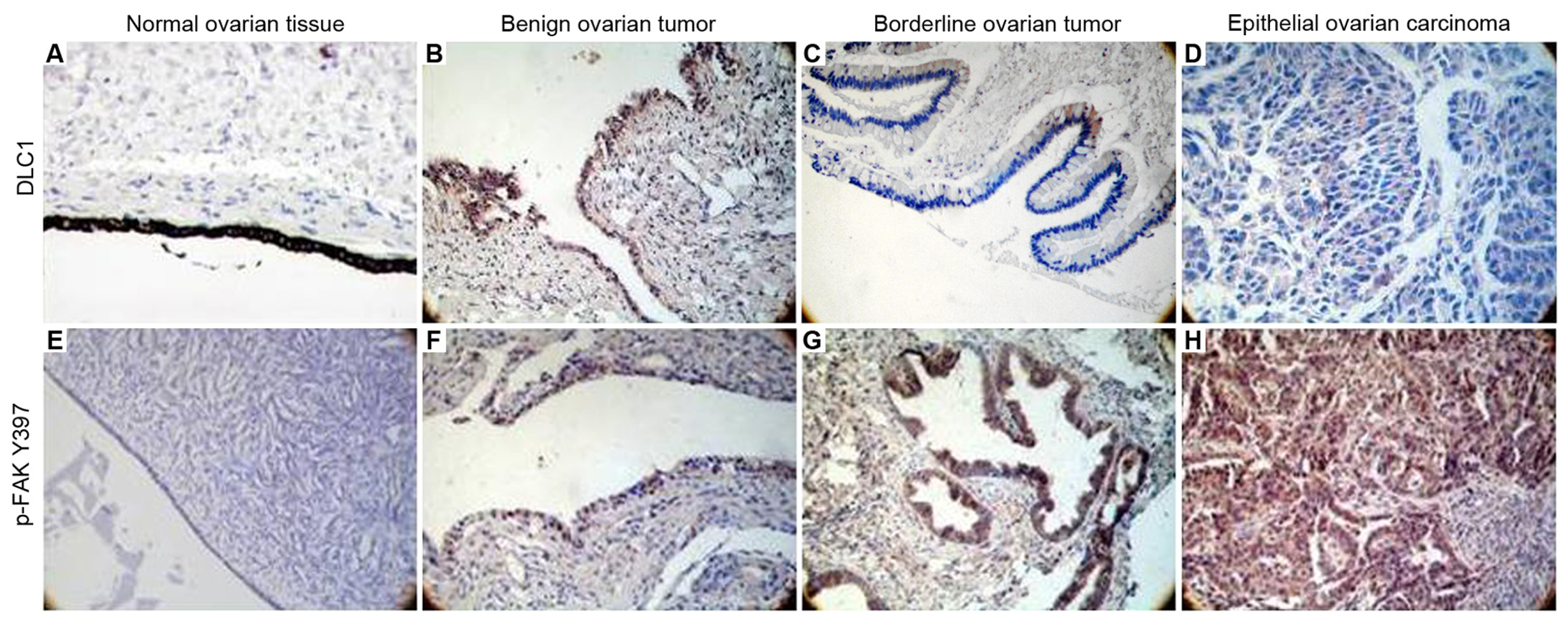
3.1. Relationship Between the Expression of DLC1 and p-FAK Y397 in EOC
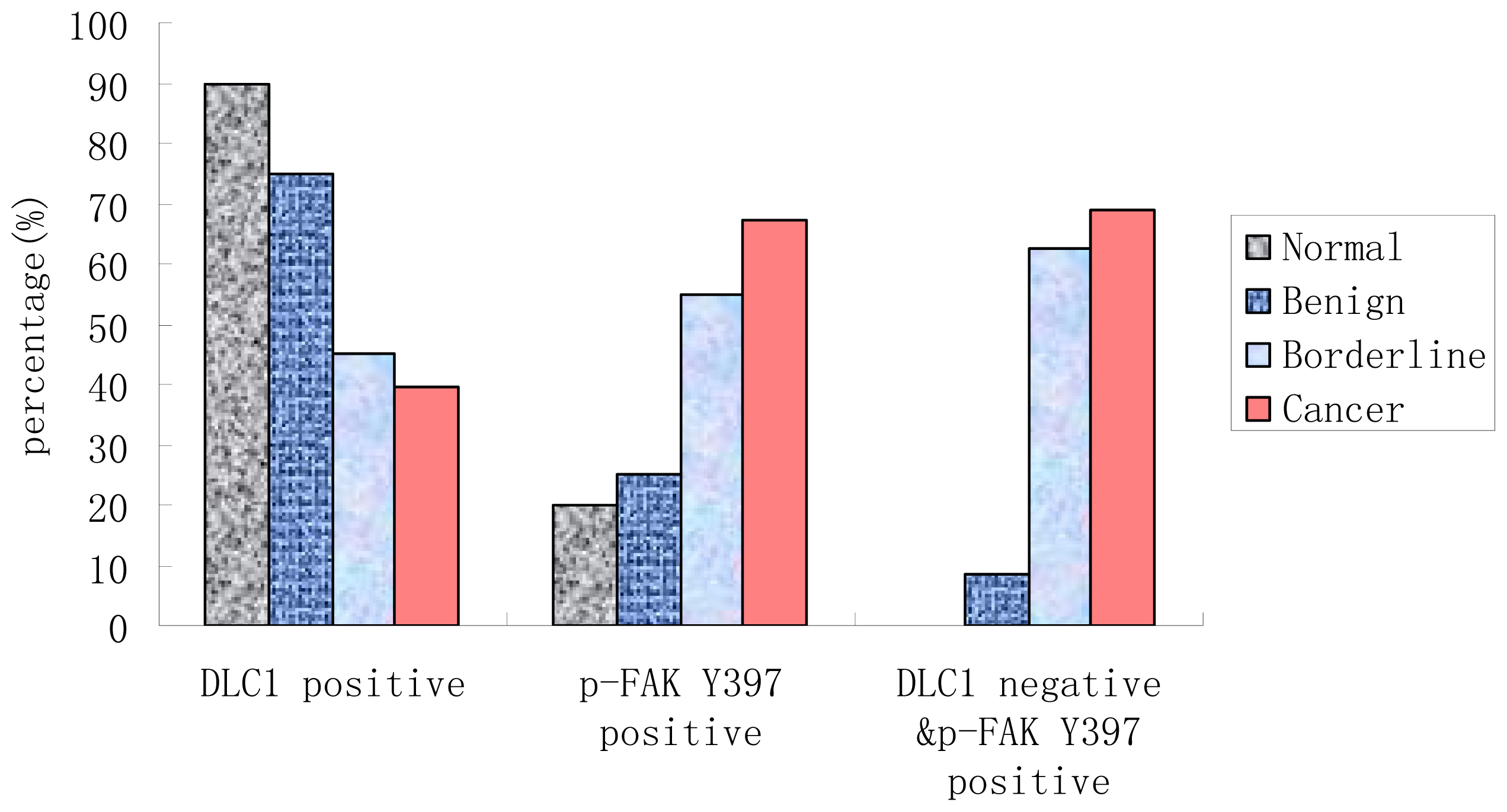
3.2. Expression of DLC1 and p-FAK Y397 in EOC
3.3. Relationship Between DLC1 or p-FAK Y397 Protein Expression and the Clinicopathological Parameters in 76 Cases of EOC
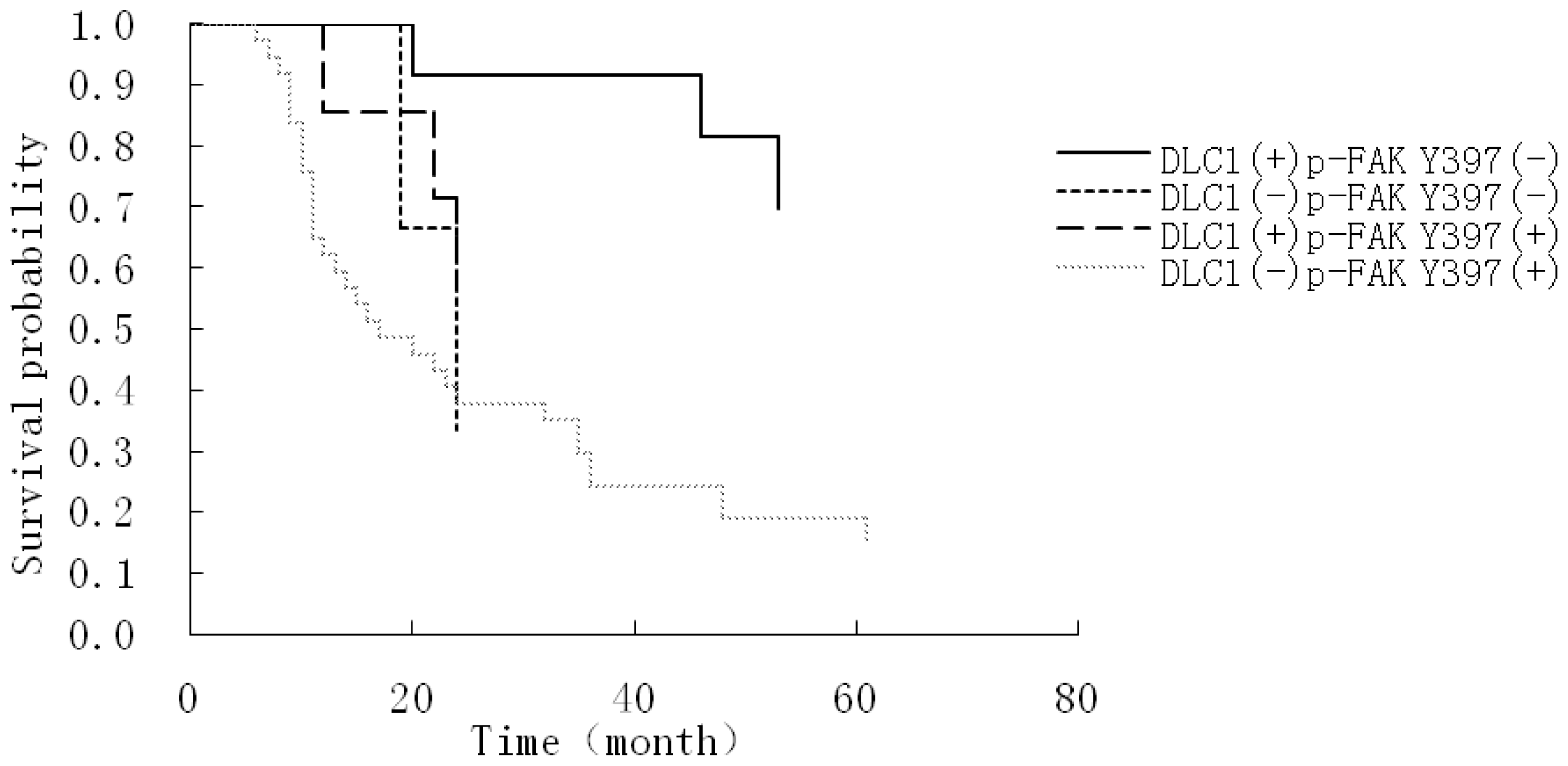
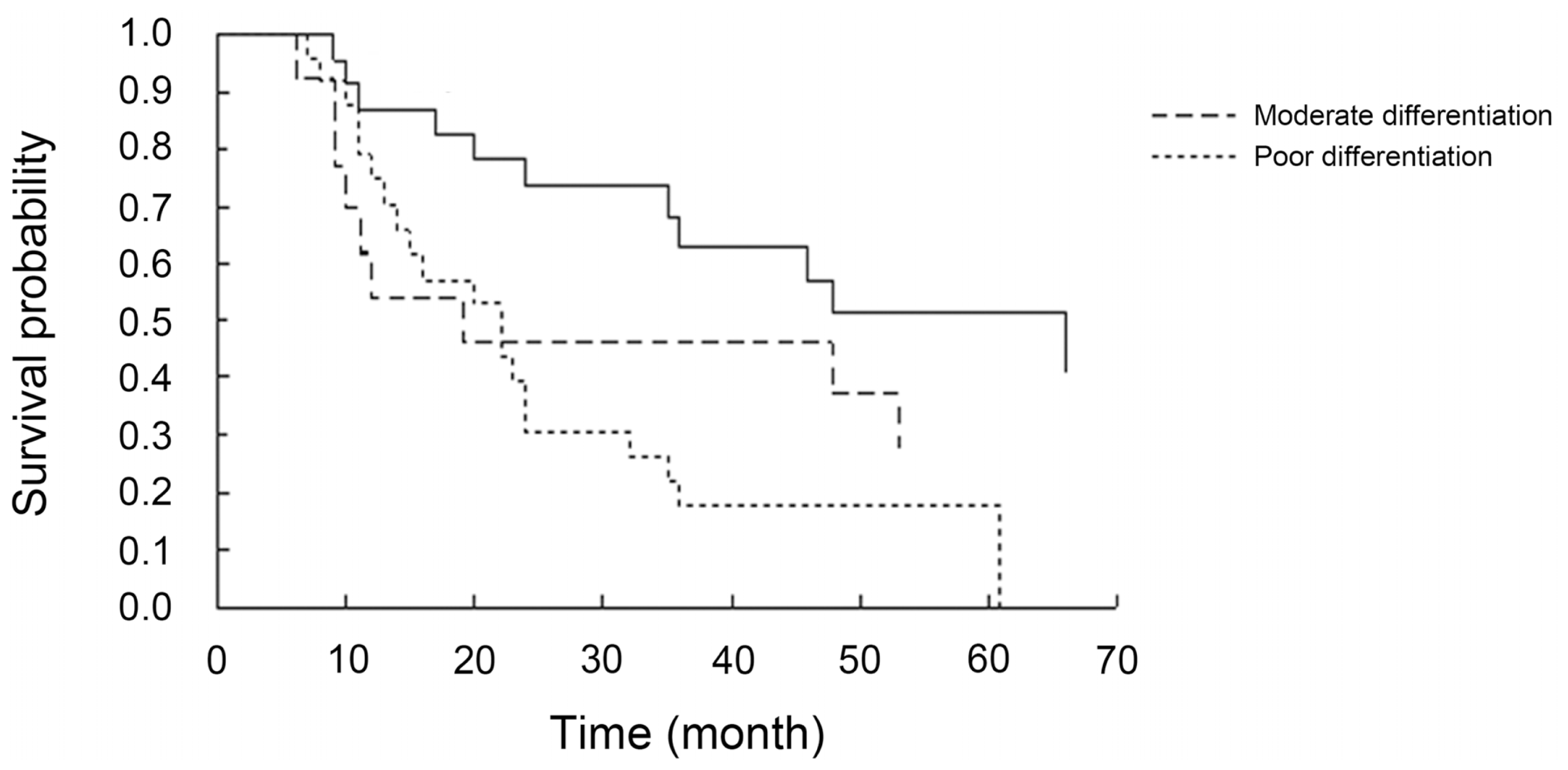
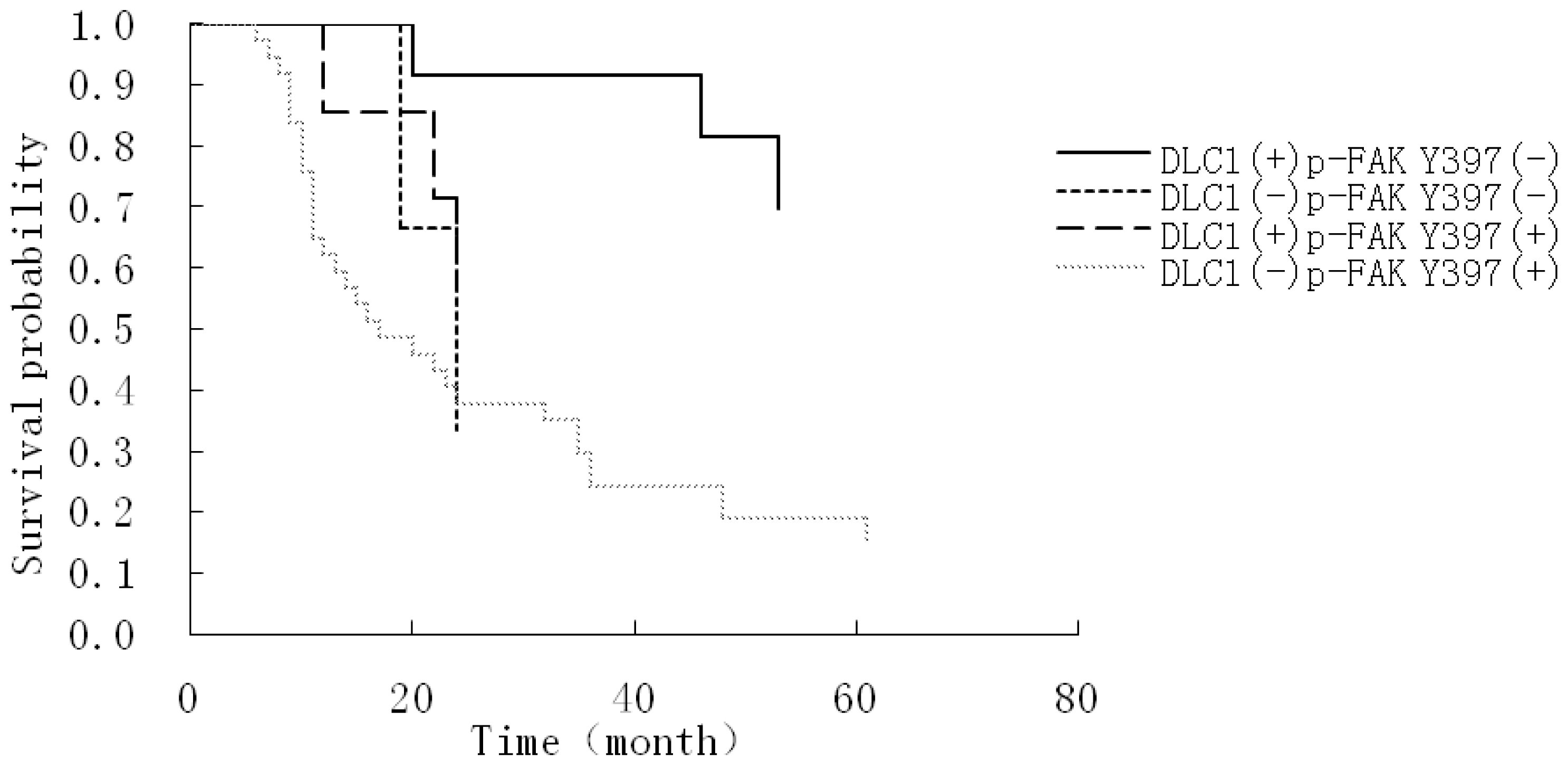
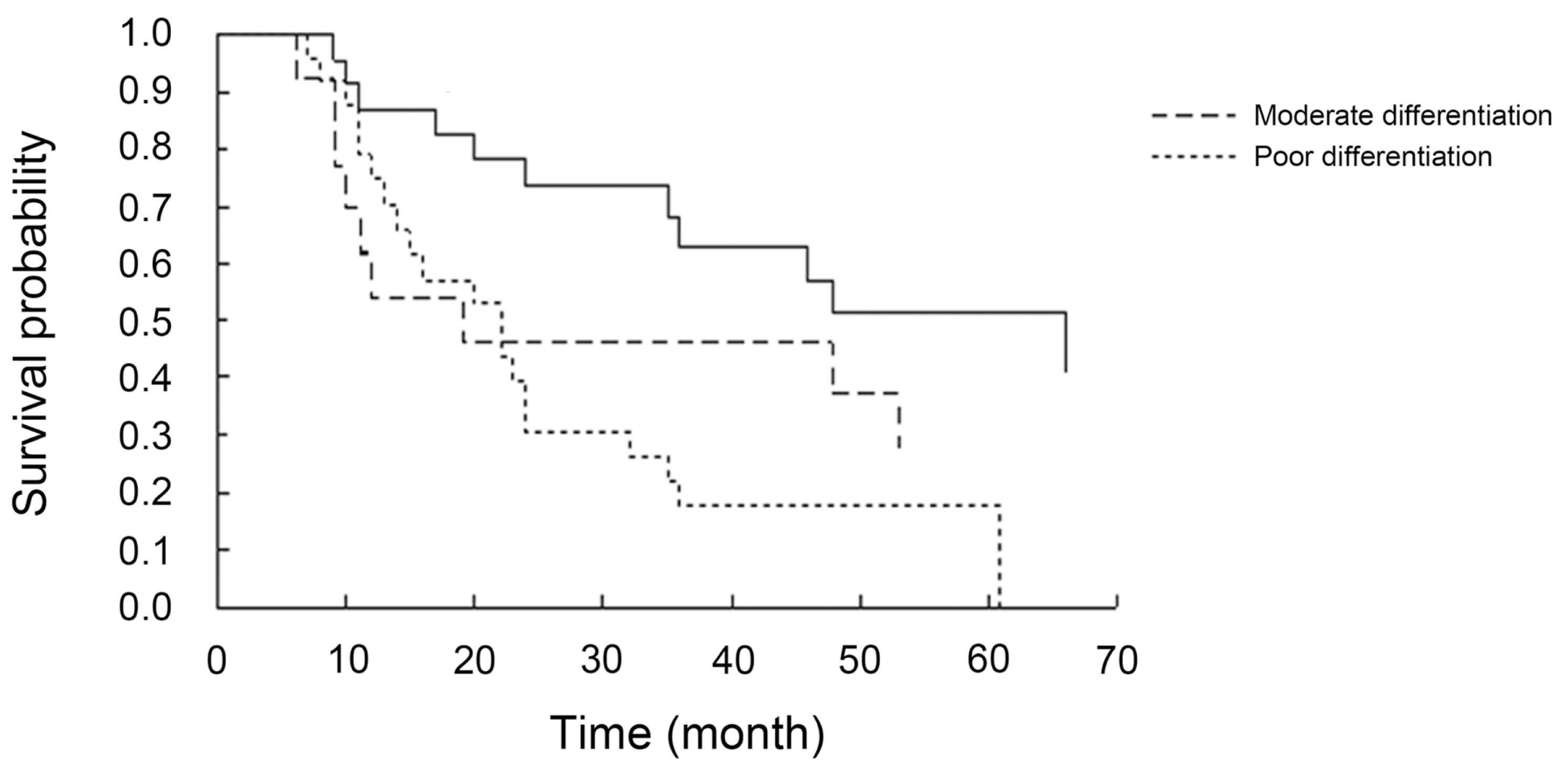
3.4. Relationship Between DLC1 and p-FAK Y397 Protein Cytoplasmic Expression, Histological Differentiation and the Prognostic Factors of EOC
4. Discussion
Acknowledgements
References
- Jemal, A.; Siegel, R.; Ward, E.; Hao, Y.; Xu, J.; Thun, M.J. Cancer statistics. CA Cancer J. Clin 2008, 58, 71–96. [Google Scholar]
- Tung, C.S.; Wong, K.K.; Mok, S.C. Biomarker discovery in ovarian cancer. Women’s Health 2008, 4, 27–40. [Google Scholar]
- Permuth-Wey, J.; Sellers, T.A. Epidemiology of ovarian cancer. Methods Mol. Biol 2009, 472, 413–437. [Google Scholar]
- Martin, L.P.; Schilder, R.J. Management of recurrent ovarian carcinoma: Current status and future directions. Semin. Oncol 2009, 36, 112–125. [Google Scholar]
- Pectasides, D.; Papaxoinis, G.; Fountzilas, G.; Aravantinos, G.; Bamias, A.; Pavlidis, N.; Kalofonos, H.P.; Timotheadou, E.; Samantas, E.; Briasoulis, E.; et al. Hellenic Cooperative Oncology Group. Epithelial ovarian cancer in Greece: A retrospective study of 1791 patients by the Hellenic Cooperative Oncology Group (HeCOG). Anticancer Res 2009, 29, 745–751. [Google Scholar]
- Larsen, E.; Blaakaer, J. Epithelial ovarian cancer: Does the time interval between primary surgery and postoperative chemotherapy have any prognostic importance? Acta Obstet. Gynecol. Scand 2009, 88, 373–377. [Google Scholar]
- Denkert, C.; Budczies, J.; Darb-Esfahani, S.; Györffy, B.; Sehouli, J.; Könsgen, D.; Zeillinger, R.; Weichert, W.; Noske, A.; Buckendahl, A.C.; et al. A prognostic gene expression index in ovarian cancer-validation across different independent data sets. J. Pathol 2009, 218, 273–280. [Google Scholar]
- Yuan, B.Z.; Miller, M.J.; Keck, C.L.; Zimonjic, D.B.; Thorgeirsson, S.S.; Popescu, N.C. Cloning, characterization, and chromosomal localization of a gene frequently deleted in human liver cancer (DLC-1) homologous to rat RhoGAP. Cancer Res 1998, 58, 2196–2199. [Google Scholar]
- Ng, I.O.; Liang, Z.D.; Cao, L.; Lee, T.K. DLC-1 is deleted in primary hepatocellular carcinoma and exerts inhibitory effects on the proliferation of hepatoma cell lines with deleted DLC-1. Cancer Res 2000, 60, 6581–6584. [Google Scholar]
- Yuan, B.Z.; Durkin, M.E.; Popescu, N.C. Promoter hypermethylation of DLC-1, a candidate tumor suppressor gene, in several common human cancers. Cancer Genet. Cytogenet 2003, 140, 113–117. [Google Scholar]
- Yuan, B.Z.; Zhou, X.; Durkin, M.E.; Zimonjic, D.B.; Gumundsdottir, K.; Eyfjord, J.E.; Thorgeirsson, S.S.; Popescu, N.C. DLC-1 gene inhibits human breast cancer cell growth and in vivo tumorigenicity. Oncogene 2003, 22, 445–450. [Google Scholar]
- Kim, T.Y.; Jong, H.S.; Song, S.H.; Dimtchev, A.; Jeong, S.J.; Lee, J.W.; Kim, T.Y.; Kim, N.K.; Jung, M.; Bang, Y.J. Transcriptional silencing of the DLC-1 tumor suppressor gene by epigenetic mechanism in gastric cancer cells. Oncogene 2003, 22, 3943–3951. [Google Scholar]
- Yuan, B.Z.; Jefferson, A.M.; Baldwin, K.T.; Thorgeirsson, S.S.; Popescu, N.C.; Reynolds, S.H. DLC-1 operates as a tumor suppressor gene in human non-small cell lung carcinomas. Oncogene 2004, 23, 1405–1411. [Google Scholar]
- Hall, A. Rho GTPases and the actin cytoskeleton. Science 1998, 279, 509–514. [Google Scholar]
- Durkin, M.E.; Yuan, B.Z.; Zhou, X.; Zimonjic, D.B.; Lowy, D.R.; Thorgeirsson, S.S.; Popescu, N.C. DLC-1, a Rho GTPase-activating protein and tumour suppressor. J. Cell. Mol. Med 2007, 11, 1185–1207. [Google Scholar]
- Kim, T.Y.; Lee, J.W.; Kim, H.P.; Jong, H.S.; Kim, T.Y.; Jung, M.; Bang, Y.J. DLC-1, a GTPase-activating protein for Rho, is associated with cell proliferation, morphology, and migration in human hepatocellular carcinoma. Biochem. Biophys. Res. Commun 2007, 355, 72–77. [Google Scholar]
- Wong, C.M.; Yam, J.W.; Ching, Y.P.; Yau, T.O.; Leung, T.H.; Jin, D.Y.; Ng, I.O. Rho GTPase-activating protein deleted in liver cancer suppresses cell proliferation and invasion in hepatocellular carcinoma. Cancer Res 2005, 65, 8861–8868. [Google Scholar]
- Goodison, S.; Yuan, J.; Sloan, D.; Kim, R.; Li, C.; Popescu, N.C.; Urquidi, V. The RhoGAP protein DLC-1 functions as a metastasis suppressor in breast cancer cells. Cancer Res 2005, 65, 6042–6053. [Google Scholar]
- Luo, Y.; Liang, F.; Zhang, Z.Y. PRL1 promotes cell migration and invasion by increasing MMP2 and MMP9 expression through Src and ERK1/2 pathways. Biochemistry 2009, 48, 1838–1846. [Google Scholar]
- Pylayeva, Y.; Gillen, K.M.; Gerald, W.; Beggs, H.E.; Reichardt, L.F.; Giancotti, F.G. Ras- and PI3K-dependent breast tumorigenesis in mice and humans requires focal adhesion kinase signaling. J. Clin. Invest 2009, 119, 252–266. [Google Scholar]
- Cornillon, J.; Campos, L.; Guyotat, D. Focal adhesion kinase (FAK), a multifunctional protein. Med. Sci (Paris) 2003, 19, 743–752. [Google Scholar]
- Watari, H.; Xiong, Y.; Hassan, M.K.; Sakuragi, N. Cyr61, a member of ccn (connective tissue growth factor/cysteine-rich 61/nephroblastoma overexpressed) family, predicts survival of patients with endometrial cancer of endometrioid subtype. Gynecologic Oncology 2009, 112, 229–234. [Google Scholar]
- Ullmannova-Benson, V.; Guan, M.; Zhou, X.; Tripathi, V.; Yang, X.Y.; Zimonjic, D.B.; Popescu, N.C. DLC1 tumor suppressor gene inhibits migration and invasion of multiple myeloma cells through RhoA GTPase pathway. Leukemia 2009, 23, 383–390. [Google Scholar]
- Wong, C.C.; Wong, C.M.; Ko, F.C.; Chan, L.K.; Ching, Y.P.; Yam, J.W.; Ng, I.O. Deleted in liver cancer 1 (DLC1) negatively regulates Rho/ROCK/MLC pathway in hepatocellular carcinoma. PLoS One 2008, 3. [Google Scholar] [CrossRef]
- Pils, D.; Horak, P.; Gleiss, A.; Sax, C.; Fabjani, G.; Moebus, V.J.; Zielinski, C.; Reinthaller, A.; Zeillinger, R.; Krainer, M. Five genes from chromosomal band 8p22 are significantly down-regulated in ovarian carcinoma: N33 and EFA6R have a potential impact on overall survival. Cancer 2005, 104, 2417–2429. [Google Scholar]
- Amirghofran, Z.; Jalali, S.A.; Hosseini, S.V.; Vasei, M.; Sabayan, B.; Ghaderi, A. Evaluation of CD44 and CD44v6 in colorectal carcinoma patients: Soluble forms in relation to tumor tissue expression and metastasis. J. Gastrointest. Cancer 2008, 39, 73–78. [Google Scholar]
- Inose, T.; Kato, H.; Kimura, H.; Faried, A.; Tanaka, N.; Sakai, M.; Sano, A.; Sohda, M.; Nakajima, M.; Fukai, Y.; et al. Correlation between connexin 26 expression and poor prognosis of esophageal squamous cell carcinoma. Ann. Surg. Oncol 2009, 16, 1704–1710. [Google Scholar]
- Hsu, P.I.; Hsieh, H.L.; Lee, J.; Lin, L.F.; Chen, H.C.; Lu, P.J.; Hsiao, M. Loss of RUNX3 expression correlates with differentiation, nodal metastasis, and poor prognosis of gastric cancer. Ann. Surg. Oncol 2009, 16, 1686–1694. [Google Scholar]
- Fan, T.; Zhao, Q.; Chen, J.J.; Chen, W.T.; Pearl, M.L. Clinical significance of circulating tumor cells detected by an invasion assay in peripheral blood of patients with ovarian cancer. Gynecol. Oncol 2009, 112, 185–191. [Google Scholar]
- Shield, K.; Ackland, M.L.; Ahmed, N.; Rice, G.E. Multicellular spheroids in ovarian cancer metastases: Biology and pathology. Gynecol. Oncol 2009, 113, 143–148. [Google Scholar]
- Zhang, T.; Zheng, J.; Liu, C.; Lu, Y. Expression of DLC-1 in clear cell renal cell carcinoma: Prognostic significance for progression and metastasis. Urol. Int 2009, 82, 380–387. [Google Scholar]
- Liao, Y.C.; Si, L.; deVere White, R.W.; Lo, S.H. The phosphotyrosine-independent interaction of DLC-1 and the SH2 domain of cten regulates focal adhesion localization and growth suppression activity of DLC-1. J. Cell Biol 2007, 176, 43–49. [Google Scholar]
- Yam, J.W.; Ko, F.C.; Chan, C.Y.; Jin, D.Y.; Ng, I.O. Interaction of deleted in liver cancer 1 with tensin2 in caveolae and implications in tumor suppression. Cancer Res 2006, 66, 8367–8372. [Google Scholar]
- Kawai, K.; Kitamura, S.Y.; Maehira, K.; Seike, J.; Yagisawa, H. START-GAP1/DLC1 is localized in focal adhesions through interaction with the PTB domain of tensin2. Adv. Enzyme Regul 2010, 50, 202–215. [Google Scholar]
- Cho, K.R.; Shih, I.M. Ovarian cancer. Ann. Rev. Pathol 2009, 4, 287–313. [Google Scholar]
- Lund, B.; Williamson, P. Prognostic factors for overall survival in patients with advanced ovarian carcinoma. Ann. Oncol 1991, 2, 281–287. [Google Scholar]
- Vergote, I. Prognostic factors in stage I ovarian carcinoma. Verh. K. Acad. Geneeskd. Belg 2001, 63, 257–271, discussion 272–276. [Google Scholar]
- Villedieu, M.; Deslandes, E.; Duval, M.; Héron, J.F.; Gauduchon, P.; Poulain, L. Acquisition of chemoresistance following discontinuous exposures to cisplatin is associated in ovarian carcinoma cells with progressive alteration of FAK, ERK and p38 activation in response to treatment. Gynecol. Oncol 2006, 101, 507–519. [Google Scholar]
- Duxbury, M.S.; Ito, H.; Benoit, E.; Zinner, M.J.; Ashley, S.W.; Whang, E.E. RNA interference targeting focal adhesion kinase enhances pancreatic adenocarcinoma gemcitabine chemosensitivity. Biochem. Biophys. Res. Commun 2003, 311, 786–792. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| p-FAK Y397(−) | p-FAK Y397(+) | χ2 | P | |
|---|---|---|---|---|
| DLC1(−) | 8 | 38 | 12.6889 | <0.001 |
| DLC1(+) | 17 | 13 |
| Group | DLC1 Positive Rate (%) | OR (95% Confidence Interval) | p-FAK Y397 Positive Rate (%) | OR (95% CI) | DLC1 Negative & p-FAK Y397 Positive n (%) | OR (95% CI) |
|---|---|---|---|---|---|---|
| Normal | 18 (90.00) | 4 (20.00) | 0 (0.00) | |||
| Benign | 15 (75.00) | 0.333 (0.056–1.971) | 5 (25.00) | 1.333 (0.300–5.925) | 1 (8.33) | - |
| Borderline | 9 (45.00) | 0.091 (0.017–0.501) | 11 (55.00) | 4.889 (1.199–19.942) | 5 (62.50) | - |
| Cancer | 30 (39.47) | 0.072 (0.016–0.335) | 51 (67.11) | 8.16 (2.469–26.972) | 38 (69.09) | - |
| DLC1 (n) | p-FAK Y397 (n) | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| (−) | (+) | χ2 | P | (−) | (+) | χ2 | P | ||
| Age | ≤50 | 19 | 13 | 0.03 | 0.86 | 10 | 22 | 0.07 | 0.79 |
| >50 | 27 | 17 | 15 | 29 | |||||
| FIGO stage | I~II | 5 | 11 | 7.27 | 0.01 * | 9 | 7 | 5.01 | 0.03 * |
| III~IV | 41 | 19 | 16 | 44 | |||||
| Lymph metastasis | N | 17 | 18 | 3.88 | 0.05 | 17 | 18 | 7.22 | 0.01 * |
| Y | 29 | 12 | 8 | 33 | |||||
| Ascites | N | 8 | 19 | 16.73 | <0.001 ** | 14 | 13 | 6.82 | 0.01 * |
| Y | 38 | 11 | 11 | 38 | |||||
| Histological differentiation | poor | 14 | 12 | 1.22 | 0.54 | 8 | 18 | 0.99 | 0.61 |
| moderate | 11 | 8 | 8 | 11 | |||||
| well | 21 | 10 | 9 | 22 | |||||
| Familial history | Y | 44 | 27 | 0.94 | 0.34 | 23 | 48 | 0.12 | 0.73 |
| N | 2 | 3 | 2 | 3 | |||||
| Poor | Moderate | Well | χ2 | P | |
|---|---|---|---|---|---|
| DLC1(+) & p-FAK Y397(−) | 7(41.18) | 4(23.53) | 6(35.29) | 0.7024 | 0.7038 |
| DLC1(−) & p-FAK Y397(+) | 13(34.21) | 7(18.42) | 18(47.37) |
| Parameter Estimation | Standard Error | χ2 | P Value | Hazard Ratio (HR) | 95% Confidence Interval | ||
|---|---|---|---|---|---|---|---|
| Histological differentiation moderate vs. well | 0.75737 | 0.46074 | 2.7021 | 0.1002 | 2.133 | 0.864 | 5.261 |
| Histological differentiation poor vs. well | 0.90318 | 0.40025 | 5.0919 | 0.0240 | 2.467 | 1.126 | 5.407 |
| Lymph metastasis | 0.24549 | 0.41800 | 0.3449 | 0.5570 | 1.278 | 0.563 | 2.900 |
| Ascites | 1.07839 | 0.65236 | 2.7326 | 0.0983 | 2.940 | 0.819 | 10.559 |
| DLC1 | 0.20422 | 0.53087 | 0.1480 | 0.7005 | 1.227 | 0.433 | 3.472 |
| p-FAK Y397 | 1.02819 | 0.59392 | 2.9971 | 0.0834 | 2.796 | 0.873 | 8.955 |
| DLC1 combined with p-FAK Y397 | 1.36657 | 0.63686 | 4.6045 | 0.0319 | 3.922 | 1.126 | 13.664 |
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Fan, D.-M.; Shi, H.-R. Pilot Study: Alteration of Deleted in Liver Cancer1 and Phosphorylated Focal Adhesion Kinase Y397 Cytoplasmic Expression and the Prognostic Value in Advanced Epithelial Ovarian Carcinoma. Int. J. Mol. Sci. 2011, 12, 8489-8501. https://doi.org/10.3390/ijms12128489
Fan D-M, Shi H-R. Pilot Study: Alteration of Deleted in Liver Cancer1 and Phosphorylated Focal Adhesion Kinase Y397 Cytoplasmic Expression and the Prognostic Value in Advanced Epithelial Ovarian Carcinoma. International Journal of Molecular Sciences. 2011; 12(12):8489-8501. https://doi.org/10.3390/ijms12128489
Chicago/Turabian StyleFan, Dong-Mei, and Hui-Rong Shi. 2011. "Pilot Study: Alteration of Deleted in Liver Cancer1 and Phosphorylated Focal Adhesion Kinase Y397 Cytoplasmic Expression and the Prognostic Value in Advanced Epithelial Ovarian Carcinoma" International Journal of Molecular Sciences 12, no. 12: 8489-8501. https://doi.org/10.3390/ijms12128489