Cholesterol-Lowering Effects of Probiotics and Prebiotics: A Review of in Vivo and in Vitro Findings

Abstract
:1. Introduction
2. Hypocholesterolemic Potential: In Vivo Evidence and Controversies
3. Dosage-Response Effects
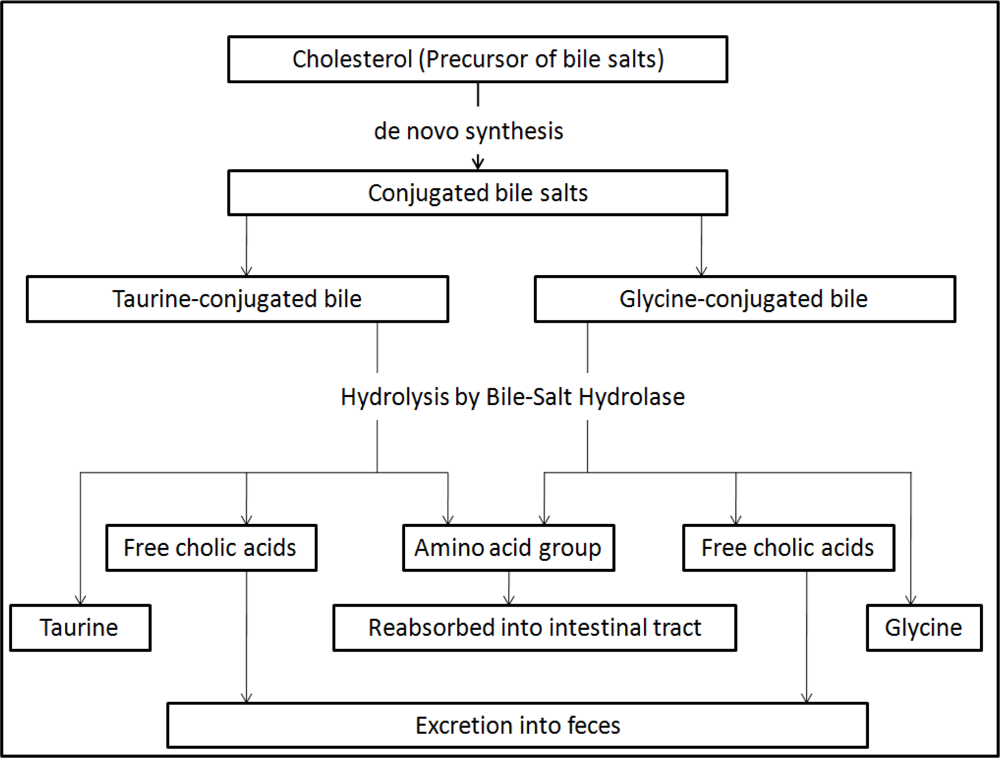
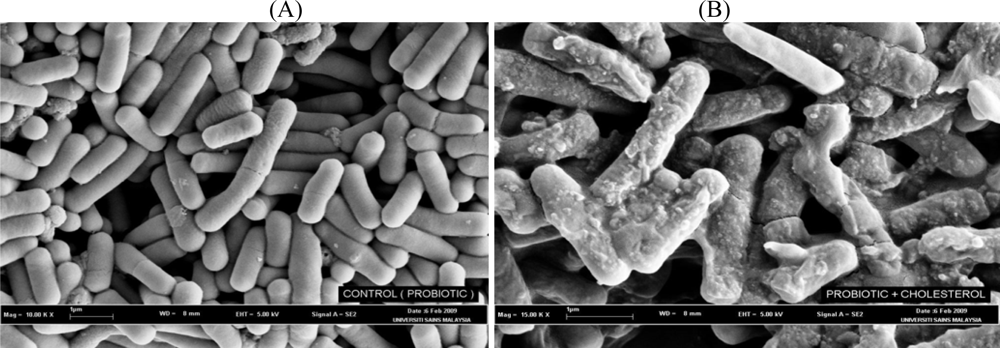
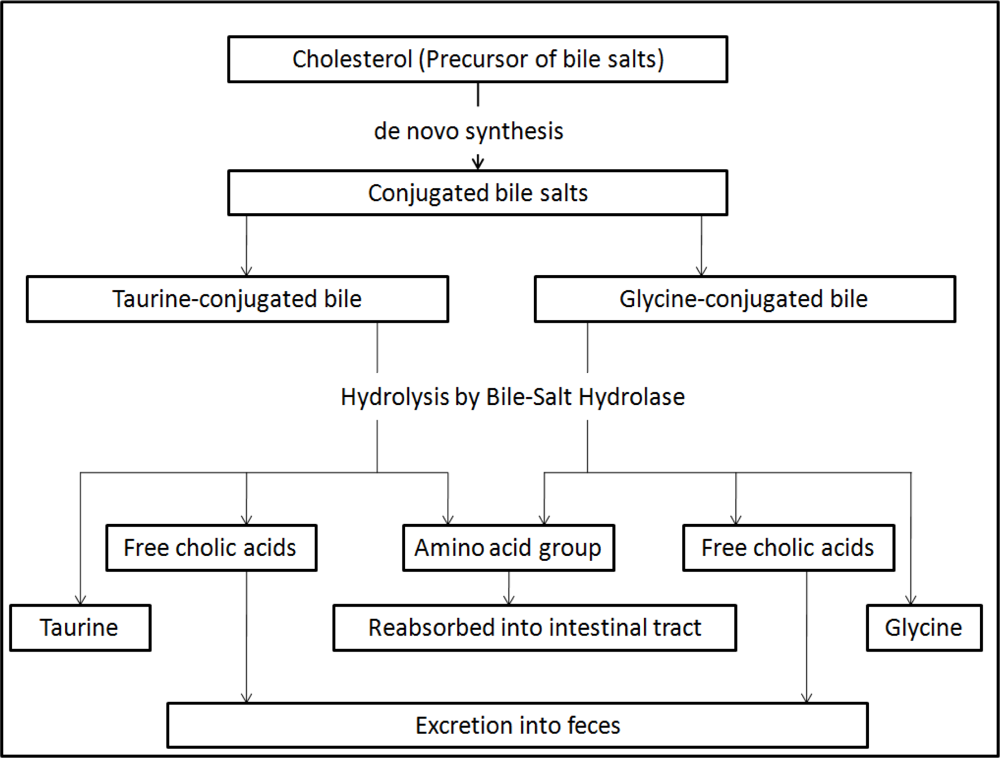
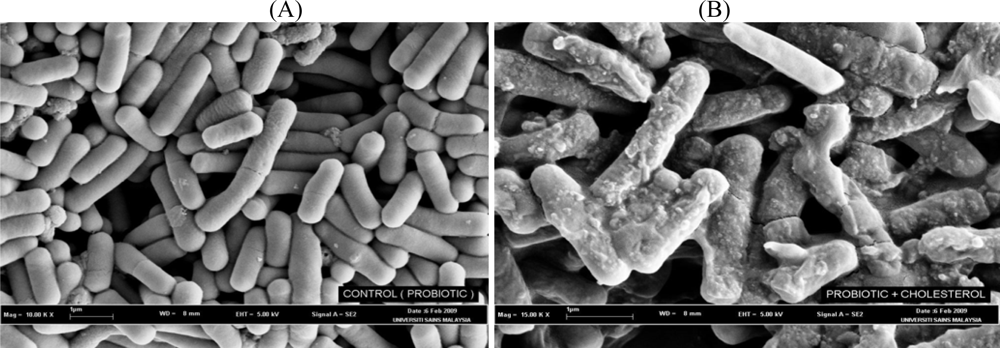
4. Mechanisms of Cholesterol-Lowering Effects
5. Safety of Probiotics and Prebiotics
6. Conclusions
Acknowledgments
References and Notes
- WHO. Cardiovascular Disease.
- Yusuf, S; Hawken, S; Ounpuu, S; Dans, T; Avezum, A; Lanas, F; McQueen, M; Budaj, A; Pais, P; Varigo, J; Lisheng, A. Effect of Potentially Modifiable Risk Factors Associated with Myocardial Infarction in 52 Countries (The INTERHEART Study): Case-Control Study. Lancet 2004, 364, 937–952. [Google Scholar]
- WHO, Diet, Nutrition and Prevention of Chronic Diseases. Report of a Joint WHO/FAO Expert Consultation, Geneva, Switzerland, 2003.
- FAO, WHO. Health and Nutritional Properties of Probiotics in Food including Powder Milk with Live Lactic Acid Bacteria; Report of a Joint FAO/WHO Expert Consultation on Evaluation of Health and Nutritional Properties of Probiotics in Food Including Powder Milk with Live Lactic Acid Bacteria, Cordoba, Argentina, 1–4 October 2001. [Google Scholar]
- Roberfroid, M. Prebiotics: The Concept Revisited. J. Nutr 2007, 137, 830–837. [Google Scholar]
- Galdeano, CM; de LeBlanc, A; de, M; Vinderola, G; Bonet, MEB; Perdigón, G. Proposed Model: Mechanisms of Immunomodulation Induced by Probiotic Bacteria. Clin. Vaccine Immunol 2007, 14, 485–492. [Google Scholar]
- Yeo, SK; Liong, MT. Angiotensin I-Converting Enzyme Inhibitory Activity and Bioconversion of Isoflavones by Probiotics in Soymilk Supplemented with Prebiotics. Int. J. Food Sci. Nutr 2010, 61, 161–181. [Google Scholar]
- Hirayama, K; Rafter, J. The Role Of Probiotic Bacteria in Cancer Prevention. Microbes. Infect 2000, 2, 681–686. [Google Scholar]
- Songisepp, E; Kulisaar, T; Hütt, P; Elias, P; Brilene, T; Zilmer, M; Mikelsaar, M. A New Probiotic Cheese with Antioxidative and Antimicrobial. J. Dairy Sci 2004, 87, 2017–2023. [Google Scholar]
- Weston, S; Halbert, A; Richmond, P; Prescott, SL. Effects of Probiotics on Topic Dermatitis: A Randomised Controlled Trial. Arch. Dis. Child 2005, 90, 892–897. [Google Scholar]
- Scholz-Ahrens, KE; Ade, P; Marten, B; Weber, P; Timm, W; Asil, Y; Glüer, C-C; Schrezenmeir, J. Prebiotics, Probiotics and Synbiotics Affect Mineral Absorption, Bone Mineral Content and Bone Structure. J. Nutr 2007, 137, 838–846. [Google Scholar]
- Baharav, E; Mor, F; Halpern, M; Weinberger, A. Lactobacillus GG Bacteria Ameliorate Arthritis in Lewis Rats. J. Nutr 2004, 134, 1964–1969. [Google Scholar]
- Ouwehand, AC. Antiallergic Effects of Probiotics. J. Nutr 2007, 137, 794–797. [Google Scholar]
- Falagas, ME; Betsi, GI; Athanasiou, S. Probiotics for Prevention of Recurrent Vulvovaginal Candidiasis: A Review. J. Antimicrob. Chemother 2006, 58, 266–272. [Google Scholar]
- Pereira, DIA; Gibson, GR. Effects of Consumption of Probiotics and Prebiotics on Serum Lipid Levels in Human. Crit. Rev. Biochem. Mol. Biol 2002, 37, 259–281. [Google Scholar]
- Gibson, GR; Fuller, R. Aspects of in Vitro and in Vivo Research Approaches Directed Toward Identifying Probiotics and Prebiotics for Human Use. J. Nutr 2000, 130, 391S–395S. [Google Scholar]
- Cummings, JH; Macfarlane, GT; Englyst, HN. Prebiotic Digestion and Fermentation. Am. J. Clin. Nutr 2001, 73, 415–420. [Google Scholar]
- Bielecka, M; Biedrzycka, E; Majkowska, A. Selection of Probiotics and Prebiotics for Synbiotic and Confirmation of Their in Vivo Effectiveness. Food Res. Int 2002, 35, 139–144. [Google Scholar]
- Schley, PD; Field, CJ. The Immune-Enhancing Effects of Dietary Fiber and Prebiotics. Br. J. Nutr 2002, 87, 221–230. [Google Scholar]
- Klinder, A; Forster, A; Caderni, G; Femia, AP; Pool-Zobel, BL. Fecal Water Genotoxicity is Predictive of Tumor-Preventive Activities by Inulin-Like Oligofructoses, Probiotics (Lactobacillus rhamnosus and Bifidobacterium lactis) and Their Synbiotic Combination. Nutr. Cancer 2004, 49, 144–155. [Google Scholar]
- Scholz-Ahrens, KE; Acil, Y; Schrezenmeir, J. Effect of Oligofructose or Dietary Calcium on Repeated Calcium and Phosphorus Balances, Bone Mineralization and Trabecular Structure in Ovariectomized Rats. Br. J. Nutr 2002, 88, 365–377. [Google Scholar]
- Cummings, JH; Macfarlane, G. Gastrointestinal Effects of Prebiotics. Br. J. Nutr 2002, 87, 145–151. [Google Scholar]
- Topping, DL; Clifton, PM. Short-Chain Fatty Acids and Human Colonic Function: Roles of Resistant Starch and Nonstarch Polysaccharides. Physiol. Rev 2001, 81, 1031–1064. [Google Scholar]
- Hinnebusch, BF; Meng, S-F; Wu, JT; Archer, SY; Hodin, RA. The Effects of Short-Chain Fatty Acids on Human Colon Cancer Cell Phenotype Are Associated with Histone Hyperacetylation. J. Nutr 2002, 132, 1012–1017. [Google Scholar]
- Dewailly, É; Blanchet, C; Gingras, S; Lemieux, S; Sauvé, L; Bergeron, J; Holub, BJ. Relations Between N-3 Fatty Acid Status and Cardiovascular Disease Risk Factors Among Quebecers. Am. J. Clin. Nutr 2001, 74, 603–611. [Google Scholar]
- Wolever, TMS; Schrade, KB; Vogt, JA; Tsihlias, EB; McBurney, MI. Do Colonic Short-Chain Fatty Acids Contribute to the Long-Term Adaptation of Blood Lipids in Subjects with Type 2 Diabetes Consuming a High-Fiber Diet? Am. J. Clin. Nutr 2002, 75, 1023–1030. [Google Scholar]
- Lambert, JM; Bongers, RS; de Vos, WM; Kleerebezem, M. Functional Analysis of Four Bile Salt Hydrolase and Penicillin Acylase Family Members in Lactobacillus plantarum WCFS1. Appl. Environ. Microbiol 2008, 74, 4719–4726. [Google Scholar]
- Pereira, DIA; Gibson, GR. Cholesterol Assimilation by Lactic Acid Bacteria and Bifidobacteria Isolated from the Human Gut. Appl. Environ. Microbiol 2002, 68, 4689–4693. [Google Scholar]
- Liong, MT; Shah, NP. Effects of A Lactobacillus casei Synbiotic on Serum Lipoprotein, Intestinal Microflora, and Organic Acids in Rats. J. Dairy Sci 2006, 89, 1390–1399. [Google Scholar]
- Liong, MT; Shah, NP. Acid and Bile Tolerance and Cholesterol Removal Ability of Lactobacilli Strains. J. Dairy Sci 2005, 88, 55–66. [Google Scholar]
- Lye, HS; Rusul, G; Liong, MT. Mechanisms of Cholesterol Removal by Lactoballi Under Conditions That Mimic the Human Gastrointestinal Tract. Int. Dairy J 2010, 20, 169–175. [Google Scholar]
- Lye, HS; Rusul, G; Liong, MT. Removal of Cholesterol by Lactobacilli via Incorporation of and Conversion to Coprostanol. J. Dairy Sci 2010, 93, 1383–1392. [Google Scholar]
- De Preter, V; Vanhoutte, T; Huys, G; Swings, J; De Vuyst, L; Rutgeerts, P; Verbeke, K. Effects of Lactobacillus casei Shirota, Bifidobacterium breve, and Oligofructose-Enriched Inulin on Colonic Nitrogen-Protein Metabolism in Healthy Humans. Am. J. Physiol. Gastrointest. Liver Physiol 2007, 292, 358–368. [Google Scholar]
- Peret-Filho, LA; Penna, FJ; Bambirra, EA; Nicoli, JR. Dose Effect of Oral Saccharomyces boulardii Treatments on Morbidity and Mortality in Immunosuppressed Mice. J. Med. Microbiol 1998, 47, 111–116. [Google Scholar]
- Land, MH; Rouster-Stevens, K; Woods, CR; Cannon, ML; Cnota, J; Shetty, AK. Lactobacillus Sepsis Associated with Probiotic Therapy. Pediatrics 2005, 115, 178–181. [Google Scholar]
- Huys, G; D’Haene, K; Swings, J. Genetic Basis of Tetracycline and Minocycline Resistance in Potentially Probiotic Lactobacillus plantarum Strain CCUG 43738. Antimicrob. Agents Chemother 2006, 50, 1550–1551. [Google Scholar]
- Shinnick, FL; Longacre, MJ; Ink, SL; Maelett, JA. Oat Fiber: Composition Versus Physiological Function in Rats. J. Nutr 1988, 118, 144–151. [Google Scholar]
- Gallaher, CM; Munion, J; Hesslink, R; Wise, J; Gallaher, DD. Cholesterol Reduction by Glucomannan and Chitosan is Mediated by Changes in Cholesterol Absorption and Bile Acid and Fat Excretion Iin Rats. J. Nutr 2000, 130, 2753–2759. [Google Scholar]
- Lichtman, AH; Clinton, SK; Iiyama, K; Connelly, PW; Libby, P; Cybulsky, MI. Hyperlipidemia and Atherosclerotic Lesion Development in LDL Receptor–Deficient Mice Fed Defined Semipurified Diets with and without Cholate. Arterioscler. Thromb. Vasc. Biol 1999, 19, 1938–1944. [Google Scholar]
- Lin, Y-G; Meijer, GW; Vermeer, MA; Trautwein, EA. Soy Protein Enhances the Cholesterol-Lowering Effect of Plant Sterol Esters in Cholesterol-Fed Hamsters. J. Nutr 2004, 134, 143–148. [Google Scholar]
- Madsen, CS; Janovitz, E; Zhang, R; Nguyen-Tran, V; Ryan, CS; Yin, X-H; Monshizadegan, H; Chang, M; D’Arienzo, C; Scheer, S; Setters, R; Search, D; Chen, X; Zhuang, S-B; Kunselman, L; Peters, A; Harrity, T; Apedo, A; Huang, C; Cuff, CA; Kowala, MC; Blanar, MA; Sun, C-Q; Robl, JA; Stein, PD. The Guinea Pig as a Preclinical Model for Demonstrating the Efficacy and Safety of Statins. J. Pharmacol. Exp. Ther 2007, 324, 576–586. [Google Scholar]
- Patterson, JK; Lei, XG; Miller, DD. The Pig as an Experimental Model for Elucidating the Mechanisms Governing Dietary Influence on Mineral Absorption. Exp. Biol. Med 2008, 233, 651–664. [Google Scholar]
- Fernandez, ML; Roy, S; Vergara-Jimenez, M. Resistant Starch and Cholestyramine Have Distinct Effects on Hepatic Cholesterol Metabolism in Guinea Pigs Fed a Hypercholesterolemic Diet. Nutr. Res 2000, 20, 837–849. [Google Scholar]
- Nguyen, TDT; Kang, JH; Lee, MS. Characterization of Lactobacillus plantarum PH04, a Potential Probiotic Bacterium with Cholesterol-Lowering Effects. Int. J. Food Microbiol 2007, 113, 358–361. [Google Scholar]
- Abd El-Gawad, IA; El-Sayed, EM; Hafez, SA; El-Zeini, HM; Saleh, FA. The Hypocholesterolaemic Effect of Milk Yoghurt and Soy-Yoghurt Containing Bifidobacteria in Rats Fed on a Cholesterol-Enriched Diet. Int. Dairy J 2005, 15, 37–44. [Google Scholar]
- Fukushima, M; Yamada, A; Endo, T; Nakano, M. Effects of A Mixture of Organisms, Lactobacillus acidophilus or Streptococcus faecalis on D6-Desaturase Activity in The Livers of Rats Fed A Fat- and Cholesterol-Enriched Diet. Nutrrients 1999, 15, 373–378. [Google Scholar]
- Anderson, JW; Gilliland, SE. Effect of Fermented Milk (Yogurt) Containing Lactobacillus acidophilus L1 on Serum Cholesterol in Hypercholesterolemic Humans. J. Am. Coll. Nutr 1999, 18, 43–50. [Google Scholar]
- Xiao, JZ; Kondo, S; Takahashi, N; Miyaji, K; Oshida, K; Hiramatsu, A; Iwatsuki, K; Kokubo, S; Hosono, A. Effects of Milk Products Fermented by Bifidobacterium longum on Blood Lipids in Rats and Healthy Adult Male Volunteers. J. Dairy Sci 2003, 86, 2452–2461. [Google Scholar]
- Causey, JL; Feirtag, JM; Gallaher, DD; Tungland, BC; Slavin, JL. Effects of Dietary Inulin on Serum Lipids, Blood Glucose and the Gastrointestinal Environment in Hypercholesterolemic Men. Nutr. Res 2000, 20, 191–201. [Google Scholar]
- Letexier, D; Diraison, F; Beylot, M. Addition of Inulin to a Moderately High-Carbohydrate Diet Reduces Hepatic Lipogenesis and Plasma Triacylglycerol Concentrations in Humans. Am. J. Clin. Nutr 2003, 77, 559–564. [Google Scholar]
- Brighenti, F; Casiraghi, MC; Canzi, E; Ferrari, A. Effect of Consumption of a Ready-to-Eat Breakfast Cereal Containing Inulin on the Intestinal Milieu and Blood Lipids in Healthy Male Volunteers. Eur. J. Clin. Nutr 1999, 53, 726–733. [Google Scholar]
- Mortensen, A; Poulsen, M; Frandsen, H. Effect of Long-Chained Fructan Raftilose ® HP on Blood Lipids and Spontaneous Atherosclerosis in Low Density Receptor Knockout Mice. Nutr. Res 2002, 22, 473–480. [Google Scholar]
- Gibson, GR; Probert, HM; Van Loo, JAE; Roberfroid, MB. Dietary Modulation of the Human Colonic Microbiota: Updating the Concept of Prebiotics. Nutr. Res. Rev 2004, 17, 257–259. [Google Scholar]
- Wang, SJ; Yu, JL; Liu, HY; Chen, WP. Characterisation and Preliminary Lipid-Lowering Evaluation of Starch from Chinese Yam. Food Chem 2008, 108, 176–181. [Google Scholar]
- Favier, M-L; Moundras, C; Demigné, C; Rémésy, C. Fermentable Carbohydrates Exert a More Potent Cholesterol-Lowering Effect than Cholestyramine. Biochim. Biophys. Acta 1995, 1258, 115–121. [Google Scholar]
- Liong, MT; Dunshea, FR; Shah, NP. Effects of A Synbiotic Containing Lactobacillus acidophilus ATCC 4962 on Plasma Lipid Profiles and Morphology of Erythrocytes in Hypercholesterolemic Pigs on High- and Low-Fat Diets. Br. J. Nutr 2007, 98, 736–744. [Google Scholar]
- Kießling, G; Schneider, J; Jahreis, G. Long-Term Consumption of Fermented Dairy Products over 6 Months Increases HDL Cholesterol. Eur. J. Clin. Nutr 2002, 56, 843–849. [Google Scholar]
- Schaafsma, G; Meuling, WJA; van Dokkum, W; Bouley, C. Effects of A Milk Product, Fermented by Lactobacillus acidophilus and with Fructo-Oligosaccharides Added, on Blood Lipids in Male Volunteers. Eur. J. Clin. Nutr 1998, 52, 436–440. [Google Scholar]
- Hatakka, K; Mutanen, M; Holma, R; Saxelin, M; Korpela, R. Lactobacillus rhamnosus LC705 together with Propionibacterium freudenreichii ssp shermanii JS Administered in Capsules Is Ineffective in Lowering Serum Lipids. J. Am. Coll. Nutr 2008, 27, 441–447. [Google Scholar]
- Simons, LA; Amansec, SG; Conway, P. Effect of Lactobacillus fermentum on Serum Lipids in Subjects with Elevated Serum Cholesterol. Nutr. Metabolism Cardiovas. Dis 2006, 16, 531–535. [Google Scholar]
- Lewis, SJ; Burmeister, S. A Double-Blind Placebo-Controlled Study of the Effects of Lactobacillus acidophilus on Plasma Lipids. Eur. J. Clin. Nutr 2005, 59, 776–780. [Google Scholar]
- Tarpila, S; Aro, A; Salminen, I; Tarpila, A; Kleemola, P; Akkila, J; Adlercreutz, H. The Effect of Flaxseed Supplementation in Processed Foods on Serum Fatty Acids and Enterolactone. Eur. J. Clin. Nutr 2002, 56, 157–165. [Google Scholar]
- Luo, J; Van Yperselle, M; Rizkalla, SW; Rossi, F; Bornet, FRJ; Slama, G. Chronic Consumption of Short-Chain Fructooligosaccharides Does Not Affect Basal Hepatic Glucose Production or Insulin Resistance in Type 2 Diabetics. J. Nutr 2000, 130, 1572–1577. [Google Scholar]
- Davidson, MH; Maki, KC; Synecki, C; Toni, SA; Drennan, KB. Effects of Dietary Inulin on Serum Lipids in Men and Women with Hypercholesterolemia. Nutr. Res 1998, 18, 503–517. [Google Scholar]
- Greany, KA; Bonorden, MJL; Halmiton-Reeves, JM; McMullen, MH; Wangen, KE; Phipps, WR; Feirtag, J; Thomas, W; Kurzer, MS. Probiotic Capsules Do Not Lower Plasma Lipid in Young Women and Men. Eur. J. Clin. Nutr 2008, 62, 232–237. [Google Scholar]
- Liong, MT. Probiotics: A Critical Review of Their Potential Role as Antihypertensive, Immune Modulators, Hypocholesterolemics and Perimenopausal Treatments. Nutr. Rev 2007, 65, 1–13. [Google Scholar]
- Food and Agriculture Organization of the United Nations and World Health Organization, Probiotics in Food: Health and Nutritional Properties and Guidelines for Evaluation, Report of a Joint FAO/WHO Expert Consultation on Evaluation of Health and Nutritional Properties of Probiotics in Food including Powder Milk with Live Lactic Acid Bacteria, Cordoba, Argentina, 1–4 October 2001 [and] Report of a Joint FAO/WHO Working Group on Drafting Guidelines for the Evaluation of Probiotics in Food, London, Ontario, Canada, 30 April-1 May 2002.
- Naruszewicz, M; Johansson, M-L; Zapolska-Downar, D; Bukowska, H. Effect of Lactobacillus plantarum 299v on Cardiovascular Disease Risk Factors in Smokers. Am. J. Clin. Nutr 2002, 76, 1249–1255. [Google Scholar]
- Lubbadeh, W; Haddadin, MSY; Al-Tamimi, MA; Robinson, RK. Effect on the Cholesterol Content of Fresh Lamb of supplementing the Feed of Awassi Ewes and Lambs with Lactobacillus acidophilus. Meat Sci 1999, 52, 381–385. [Google Scholar]
- Ha, CG; Cho, JK; Lee, CH; Chai, YG; Ha, YA; Shin, SH. Cholesterol Lowering Effect of Lactobacillus plantarum Isolated from Human Feces. J. Mol. Microbiol. Biotechnol 2006, 16, 1201–1209. [Google Scholar]
- Greany, KA; Nettleton, JA; Wangen, KE; Thomas, W; Kurzer, MS. Probiotic Consumption Does Not Enhance the Cholesterol-Lowering Effect of Soy in Postmenopausal Women. J. Nutr 2004, 134, 3277–3283. [Google Scholar]
- Jeun, J; Kim, S-Y; Cho, S-Y; Jun, H-J; Park, H-J; Seo, J-G; Chung, M-J; Lee, S-J. Hypocholesterolemic Effects of Lactobacillus plantarum KCTC3928 by Increased Bile Acid Excretion in C57BL/6 Mice. Nutrition 2010, 26, 321–330. [Google Scholar]
- Bertolami, MC; Faludi, AA; Batlouni, M. Evaluation of the Effects of a New Fermented Milk Product (Gaio) on Primary Hypercholesterolemia. Eur. J. Clin. Nutr 1999, 53, 97–101. [Google Scholar]
- FAO Technical Meeting on Prebiotics: Food Quality and Standards Service (AGNS); Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 15–16 September 2007.
- Vogt, JA; Ishii-Schrade, KB; Pencharz, PB; Jones, PJH; Wolever, TMS. L-Rhamnose and Lactulose Decrease Serum Triacylglycerols and Their Rates of Synthesis, But Do Not Affect Serum Cholesterol Concentrations in Men. J. Nutr 2006, 136, 2160–2166. [Google Scholar]
- Robinson, RR; Feirtag, J; Slavin, JL. Effects of Dietary Arabinogalactan on Gastrointestinal and Blood Parameters in Healthy Human Subjects. J. Am. Coll. Nutr 2001, 20, 279–285. [Google Scholar]
- Trautwein, EA; Rieckhoff, D; Erbersdobler, HF. Dietary Inulin Lowers Plasma Cholesterol and Triacylglycerol and Alters Biliary Bile Acid Profile in Hamsters. J. Nutr 1998, 128, 1937–1943. [Google Scholar]
- Li, XJ; Piao, XS; Kim, SW; Liu, P; Wang, L; Shen, YB; Jung, SC; Lee, HS. Effects of Chito-Oligosaccharide Supplementation on Performance, Nutrient Digestibility, and Serum Composition in Broiler Chickens. Poultry Sci 2007, 86, 1107–1114. [Google Scholar]
- Hsu, CK; Liao, JW; Chung, YC; Hsieh, CP; Chan, YC. Xylooligosaccharides and Fructooligosaccharides Affect the Intestinal Microbiota and Precancerous Colonic Lesion Development in Rats. J. Nutr 2004, 134, 1523–1528. [Google Scholar]
- Chen, H; Liu, L-J; Zhu, J-J; Xu, B; Li, R. Effect of Soybean Oligosaccharides on Blood Lipid, Glucose Levels and Antioxidant Enzymes Activity in High Fat Rats. Food Chem 2010, 119, 1633–1636. [Google Scholar]
- Alles, MS; de Roos, NM; Bakx, JC; van de Lisdonk, E; Zock, PL; Hautvast, JGAJ. Consumption of Fructooligosaccharides Does Not Favorably Affect Blood Glucose and Serum Lipid Concentrations in Patients with Type 2 Diabetes. Am. J. Clin. Nutr 1999, 69, 64–69. [Google Scholar]
- Vulevic, J; Drakoularakou, A; Yaqoob, P; Tzortzis, G; Gibson, GR. Modulation of the Fecal Microflora Profile and Immune Function by a Novel Trans-Galactooligosaccharide Mixture (B-GOS) in Healthy Elderly Volunteers. Am. J. Clin. Nutr 2008, 88, 1438–1446. [Google Scholar]
- Liong, MT; Shah, NP. Optimization of Cholesterol Removal by Probiotics in the Presence of Prebiotics by Using a Response Surface Method. Appl. Environ. Microbiol 2005, 71, 1745–1753. [Google Scholar]
- Zhang, F; Hang, X; Fan, X; Li, G; Yang, H. Selection and Optimization Procedure of Synbiotic for Cholesterol Removal. Anaerobe 2007, 13, 185–192. [Google Scholar]
- Begley, M; Hill, C; Gahan, CGM. Bile Salt Hydrolase Activity in Probiotics. Appl. Environ. Microbiol 2006, 72, 1729–1738. [Google Scholar]
- Jones, ML; Chen, H; Ouyang, W; Metz, T; Prakash, S. Microencapsulated Genetically Engineered Lactobacillus plantarum 80 (pCBH1) for Bile Acid Deconjugation and Its Implication in Lowering Cholesterol. J. Biomed. Biotechnol 2004, 1, 61–69. [Google Scholar]
- Usman, HA. Bile Tolerance, Taurocholate Deconjugation, and Binding of Cholesterol by Lactobacillus gasseri Strains. J. Dairy Sci 1999, 82, 243–248. [Google Scholar]
- Kimoto, H; Ohmomo, S; Okamoto, T. Cholesterol Removal from Media by Lactococci. J. Dairy Sci 2002, 85, 3182–3188. [Google Scholar]
- Chiang, YR; Ismail, W; Heintz, D; Schaeffer, C; van Dorsselaer, A; Fuchs, G. Study of Anoxic and Oxic Cholesterol Metabolism by Sterolibacterium denitrificans. J. Bacteriol 2008, 190, 905–914. [Google Scholar]
- Arjmandi, BH; Craig, J; Nathani, S; Reeves, RD. Soluble Dietary Fiber and Cholesterol Influence in Vivo Hepatic and Intestinal Cholesterol Biosynthesis in Rats. J. Nutr 1992, 122, 1559–1565. [Google Scholar]
- Kim, M-H; Shin, HK. The Water-Soluble Extract Of Chicory Influences Serum and Liver Lipid Concentrations, Cecal Short-Chain Fatty Acid Concentrations and Fecal Lipid Excretion in Rats. J. Nutr 1998, 128, 1731–1736. [Google Scholar]
- Dikeman, CL; Murphy, MR; Fahey, GC. Dietary Fibers Affect Viscosity of Solutions and Simulated Human Gastric and Small Intestinal Digesta. J. Nutr 2006, 136, 913–919. [Google Scholar]
- Rossi, M; Corradini, C; Amaretti, A; Nicolini, M; Pompei, A; Zanoni, S; Matteuzzi, D. Fermentation of Fructooligosaccharides and Inulin By Bifidobacteria: A Comparative Study of Pure and Fecal Cultures. Appl. Environ. Microbiol 2005, 71, 6150–6158. [Google Scholar]
- Olano, A; Chua, J; Schroeder, S; Minari, A; Salvia, ML; Hall, G. Weissella confusa (Basonym: Lactobacillus confusus) Bacteremia: A Case Report. J. Clin. Microbiol 2001, 39, 1604–1607. [Google Scholar]
- d’Escrivan, T; Meybeck, A; Legout, L; Georges, H; Senneville, E. Severe Sepsis Due to Lactobacillus helvetica Bacteremia. Med. Maladies Infect 2003, 33, 356–357. [Google Scholar]
- Chazan, B; Raz, R; Shental, Y; Sprecher, H; Colodner, R. Bacteremia and Pyelonephritis Caused by Lactobacillus jensenii in a Patient with Urolithiasis. Isr. Med. Assoc. J 2008, 10, 164–165. [Google Scholar]
- Zé-Zé, L; Tenreiro, R; Duarte, A; Salgado, MJ; Melo-Cristino, J; Lito, L; Carmo, MM; Felisberto, S; Carmo, G. Case of Aortic Endocarditis Caused by Lactobacillus casei. J. Med. Microbiol 2004, 53, 451–453. [Google Scholar]
- Rautio, M; Jousimies-Somer, H; Kauma, H; Pietarinen, I; Saxelin, M; Tynkkynen, S; Koskela, M. Liver Abscess Due to a Lactobacillus rhamnosus Indistinguishable from L. rhamnosus Strain GG. Clin. Infect. Dis 1999, 28, 1159–1160. [Google Scholar]
- Ruseler-van Ebmden, JHG; van Lieshout, L; Gosselink, M; Marteau, P. Inability of Lactobacillus casei Strain GG, Lactobacillus acidophilus, and Bifidobacterium bifidum to Degrade Intestinal Mucosa Glycoproteins; Clearing the Way for Mucosa-Protective Therapy. Scand. J. Gastroenterol 1995, 30, 675–680. [Google Scholar]
- Abe, F; Muto, M; Yaeshima, T; Iwatsuki, K; Aihara, H; Ohashi, Y; Fujisawa, T. Safety Evaluation of Probiotic Bifidobacteria by Analysis of Mucin Degradation Activity and Translocation Ability. Anaerobe 2010, 16, 131–136. [Google Scholar]
- Tan, KP; Yang, M; Ito, S. Activation of Nuclear Factor (Erythroid-2 Like) Factor 2 by Toxic Bile Acids Provokes Adaptive Defense Responses to Enhance Cell Survival at the Emergence of Oxidative Stress. Mol. Pharmacol 2007, 72, 1380–1390. [Google Scholar]
- Miettinen, M; Vuopio-Varkila, J; Varkila, K. Production of Human Necrosis Factor Alpha, Interleukin 6, and Interleukin 10 Is Induced by Lactic Acid Bacteria. Infect. Immun 1996, 64, 5403–5405. [Google Scholar]
- Shimizu, N; Kamezaki, F; Shigematsu, S. Tracking of Microinjected DNA in Live Cells Reveals the Intracellular Behavior and Elimination of Extrachromosomal Genetic Material. Nucleic Acids Res 2005, 33, 6296–6307. [Google Scholar]
- Deichelbohrer, I; Alonso, JC; Lüder, G; Trautner, TA. Plasmid Transduction by Bacillus subtilis Bacteriophage SPP1: Effects of DNA Homology between Plasmid and Bacteriophage. J. Bacteriol 1985, 162, 1238–1243. [Google Scholar]
- Merryweather, A; Barth, PT; Wilkins, BM. Role and Specificity of Plasmid RP4-Encoded DNA Primase in Bacterial Conjugation. J. Bacteriol 1986, 167, 12–17. [Google Scholar]
- Prágai, Z; Holczinger, A; Sík, T. Transformation of Bacillus licheniformis Protoplasts by Plasmid DNA. Microbiology 1994, 140, 305–310. [Google Scholar]
- Liong, MT; Yeo, SK; Kuan, CY; Fung, WY; Ewe, JA. Antibiotic Resistance and Probiotics: Roles, Mechanisms and Evidence. In Antibiotic Resistance: Causes and Risk Factors; Bonilla, AR, Muniz, KP, Eds.; Nova Science Publishers, Inc.:: New York, NY, USA, 2009; pp. 5–38. [Google Scholar]
- D’Aimmo, MR; Modesto, M; Biavati, B. Antibiotic Resistance of Lactic Acid Bacteria and Bifidobacterium spp. Isolated from Dairy and Pharmaceutical Products. Int. J. Food Microbiol 2007, 115, 35–42. [Google Scholar]
- Hummel, AS; Hertel, C; Holzapfel, WH; Franz, CMAP. Antibiotic Resistances of Starter and Probiotic Strains of Lactic Acid Bacteria. Appl. Environ. Microbiol 2007, 73, 730–739. [Google Scholar]
- Coussement, PAA. Inulin and Oligofructose: Safe Intakes and Legal Status. J. Nutr 1999, 129, 1412–1417. [Google Scholar]
- Williams, CM. Effects of Inulin on Lipid Parameters in Humans. J. Nutr 1999, 129, 1471–1473. [Google Scholar]
- Bruggencate, SJMT; Bovee-Oudenhoven, IMJ; Lettink-Wissink, MLG; van der Meer, R. Dietary Fructooligosaccharides Increase Intestinal Permeability in Rats. J. Nutr 2005, 135, 837–842. [Google Scholar]
- Bruggencate, SJMT; Bovee-Oudenhoven, IMJ; Lettink-Wissink, MLG; Katan, MB; van der Meer, R. Dietary Fructooligosaccharides Affect Intestinal Barrier Function in Healthy Men. J. Nutr 2006, 136, 70–74. [Google Scholar]

{kind=link}
{kind=link}
| Compound(s) | Experimental design | Subjects | Dose; duration of the study | Effects | Ref. |
|---|---|---|---|---|---|
| Inulin | Randomized, placebo-controlled, double-blind & crossover. | 8 volunteers. | 3–4 g/100 of inulin & wheat fiber daily for 12 weeks. | No significant improvement in lipid profiles. | [62] |
| Fructooligo-saccharides (FOS) | Randomized, placebo-controlled, double-blind & crossover. | 10 diabetic patients (6 men and 4 women); with plasma TC of 4.85–5.58 mmol/L. | 20 g FOS/day for 4 weeks. | No significant improvement in lipid profiles. | [63] |
| Inulin | Randomized, placebo-controlled, double-blind & crossover designed study; with 2 six-week treatment periods, separated by a six-week washout period. | 25 subjects; with baseline LDL-C ranging from 3.36–5.17 mmol/L. | 45 g chocolate bar (containing of 18 g of inulin) daily during treatment period. | No significant improvement in lipid profiles. | [64] |
| L. acidophilus & B. longum & fructooligo-saccharides (FOS) | Randomized, single-blind, placebo-controlled & parallel. | 55 normocholesterolemic volunteers. | 3 capsules of synbiotics product (consisted of 109 CFU/g of L. acidophilus & B. longum, & 10–15 mg of FOS) once daily for 2 months. | No significant improvement in lipid profiles. | [65] |
| Products/Probiotic strains | Experimenta l design | Animals/Subjects | Dose; duration of the study | Effects | Ref. |
|---|---|---|---|---|---|
| Animal models | |||||
| L. plantarum CK 102(healthy human isolate) | Randomized, placebo-controlled, parallel. | 32 Sprague-Dawley (SD) male rats; 5 weeks old; induced hypercholesterolemic; mean BW of 129 ± 1 g. | 5.0 × 107 CFU/mL daily, 6 weeks. | TC: 27.9% decrease (P < 0.05) LDL-C: 28.7% decrease (P < 0.05) TG: 61.6% decrease (P < 0.05) | [70] |
| L. acidophilus (wild chickens & human isolates) | Randomized, placebo-controlled, parallel. | 30 Awassi weaning lambs; hypercholesterolemic ; mean BW of 55.1 ± 3.4 & 57.9 ± 4.7 kg for the treated & control groups, respectively. | 1 × 109 CFU/capsule, 2 capsules daily, 120 days. | TC: 22.6% decrease (P < 0.05) [treatment group with mean plasma TC of 72.8 ± 5.7 mg/100 mL; control group with mean plasma TC of 94.0 ± 7.8 mg/100 mL] | [69] |
| L. plantarum KCTC3928 (Cellbiotech Co. Ltd, Korea) | Randomized, placebo-controlled, parallel. | 21 six-week-old C57BL/6 male mice; induced hypercholesterolemic. | 1 × 109 CFU/mL of L. plantarum KCTC3928, 4 weeks. | TC: 33% decrease (P < 0.05) LDL-C: 42% decrease (P < 0.05) TG: 32% decrease (P < 0.05) HDL-C: 35% increase (P < 0.05) | [72] |
| Human models | |||||
| L. plantarum 299v (ProViva) | Randomized, placebo-controlled, double-blind, parallel. | 36 healthy volunteers with moderately elevated fibrinogen concentrations (>3.0 g/L); 35–45 years old; mean TC of 5.59 ± 0.88 mmol/L for treatment group & 5.51 ± 0.75 mmol/L for control group. | 400 mL of rose-hip drink containing 5.0 × 107 CFU/mL daily, 6 weeks. | TC: 2.5% decrease LDL-C: 7.9% decrease | [68] |
| Enterococcus faecium & 2 strains of Streptococcus thermophilus (Causido®; Gaio®) | Randomized, placebo controlled, double-blind, crossover. | 32 patients; 36–65 years old; mean TC of 248.47 ± 26.75 mg/dL, mean LDL-C of 172.22 ± 21.17 mg/dL. | 200 g of Gaio® containing 105–109/mL of E. faecium & 5–20 × 108/mL of S. thermophilus daily, 16 weeks. | TC: 5.3% decrease (P = 0.004) LDL-C: 6.15% decrease (P = 0.012) | [73] |
| Prebiotics/Oligosaccharides | Experimental design | Animals/Subjects | Dose; duration of the study | Effects | Ref. |
|---|---|---|---|---|---|
| Animal Models | |||||
| Inulin | Randomized, placebo-controlled, parallel. | 10 male golden Syrian hamsters, mean BW of 58 ± 4 g. | 16% of inulin daily, 5 weeks. | TC: 29% decrease (P < 0.05) TG: 63% decrease (P < 0.05) | [77] |
| Chito-oligosaccharides (COS) | Randomized, placebo-controlled, parallel. | 49 male Arbor Acres broiler chickens; 196 days old. | 100 mg/kg BW daily, 42 days. | TG: 26.9% decrease (P < 0.05) HDL-C: 12.3% increase (P < 0.05) | [78] |
| Xylo-oligosaccharides (XOS) | Randomized, placebo-controlled, parallel. | 40 male Sprague-Dawley rats; 6 weeks old. | 60 g XOS/kg diet, 35 days. | TG: 33.9% decrease (P < 0.05) | [79] |
| Soybean oligosaccharides | Randomized, placebo-controlled, parallel. | 50 Wistar rats; aged of 4-week; induced hypercholesterolemic. | 450 mg/kg BW/day, 45 days. | TC: 38.5% decrease (P < 0.05) LDL-C: 43.0% decrease (P < 0.05) TG: 40.8% decrease (P < 0.05) HDL-C: 81.9% increase (P < 0.05) (compared to the positive control group) | [80] |
| Human Models | |||||
| Inulin | Randomized, placebo-controlled, double-blind, crossover. | 8 healthy volunteers; 23–32 years old, BMI of 19–25 kgm−2. | 10 g/day, 3 weeks. | TG: 16.3% decrease (P < 0.05) | [50] |
| Fructo-oligosaccharides (FOS) | Randomized, placebo-controlled, single-blind, crossover. | 20 diabetic & hypercholesterolemic volunteers with fasting serum TC concentrations > 6 mmol/L. | 15 g/day, two 20 days treatment period, no washout period between treatments. | HDL-C: 2.8% increase | [81] |
| Galacto-oligosaccharides | Randomized, placebo-controlled, double-blind, crossover. | 44 elderly volunteers (16 men & 28 women); 64–79 years old. | 5.5 g/d, two 10 weeks treatment period, 4-week washout period. | No significant improvement in lipid profiles. | [82] |
| Patient’s age (year)/sex | Diagnosis | Underlying condition (s) | Organism (s) isolated | Therapy; duration of the therapy | Outcome | Ref. |
|---|---|---|---|---|---|---|
| 46/M | Bacteremia | Short-bowel syndrome, history of Enterococcus faecalis aortic valve endocarditis, Klebsiella pneumonia bacteremia & candidemia. | L. confuses; isolated from blood culture. | Piperacillin-tazobactam & gentamicin, 4 weeks. | Recovered. | [94] |
| 21/F | Bacteremia | Fever, drownsiness & stiff neck. | L. helvetica; isolated from blood culture. | Amoxicillin & gentamicin, 10 days; clindamycin, 15 days. | Recovered | [95] |
| 59/F | Bacteremia | Hypertension, diabetes mellitus type 2, history of breast cancer with conservative surgery & kidney stone. | L. jensenii; isolated from blood & urine culture. | Ampicillin, 2 weeks. | Recovered | [96] |
| 53/M | Endocarditis | History of rheumatic fever. | L. casei; isolated from the blood & bone marrow. | Valve replacement surgery and was treated with doxycycline. | Recovered | [97] |
| 74/F | Liver abscess | Hypertension, non-insulin-dependent diabetes mellitus & pneumonia. | L. rhamnosus; isolated from blood culture. | Ciprofloxacin & clindamycin, 6 weeks. | Recovered | [98] |
© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Ooi, L.-G.; Liong, M.-T. Cholesterol-Lowering Effects of Probiotics and Prebiotics: A Review of in Vivo and in Vitro Findings. Int. J. Mol. Sci. 2010, 11, 2499-2522. https://doi.org/10.3390/ijms11062499
Ooi L-G, Liong M-T. Cholesterol-Lowering Effects of Probiotics and Prebiotics: A Review of in Vivo and in Vitro Findings. International Journal of Molecular Sciences. 2010; 11(6):2499-2522. https://doi.org/10.3390/ijms11062499
Chicago/Turabian StyleOoi, Lay-Gaik, and Min-Tze Liong. 2010. "Cholesterol-Lowering Effects of Probiotics and Prebiotics: A Review of in Vivo and in Vitro Findings" International Journal of Molecular Sciences 11, no. 6: 2499-2522. https://doi.org/10.3390/ijms11062499
APA StyleOoi, L.-G., & Liong, M.-T. (2010). Cholesterol-Lowering Effects of Probiotics and Prebiotics: A Review of in Vivo and in Vitro Findings. International Journal of Molecular Sciences, 11(6), 2499-2522. https://doi.org/10.3390/ijms11062499