Effects of Titanium Dioxide Nanoparticle Aggregate Size on Gene Expression


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
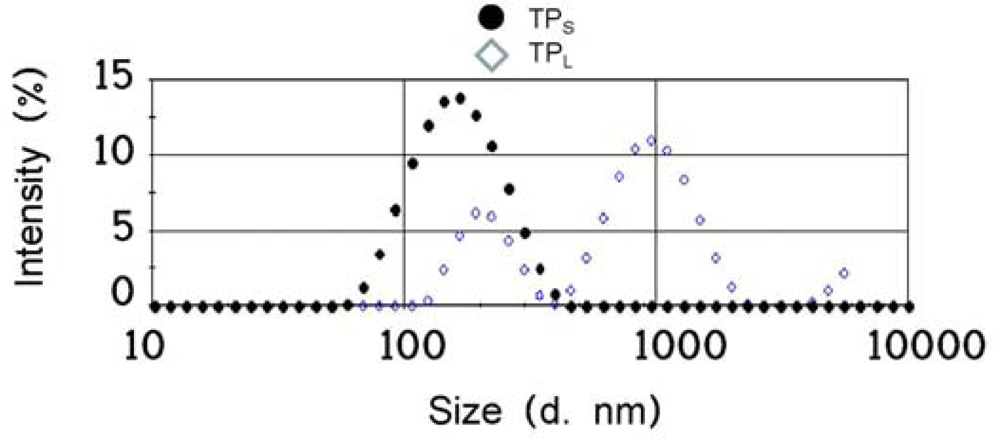
2.1. Preparation of Two Different Sizes of Aggregate Titaniananoparticles
2.2. Microscopic Images of Titania Particle-Exposed Cells
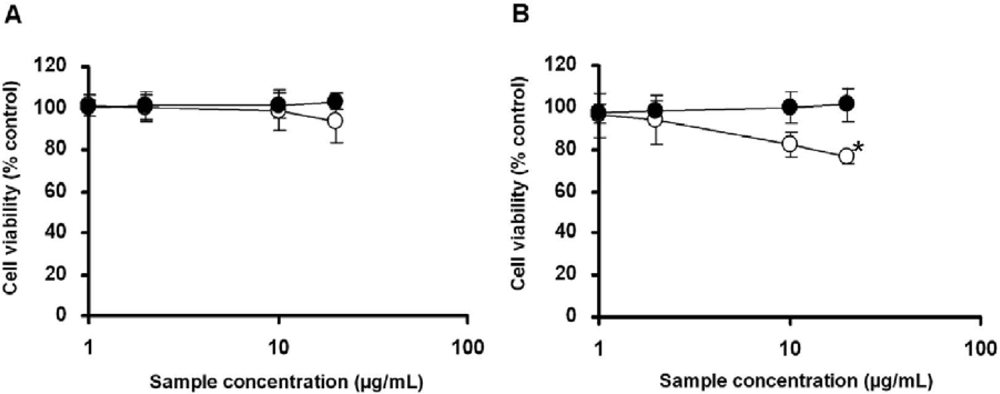
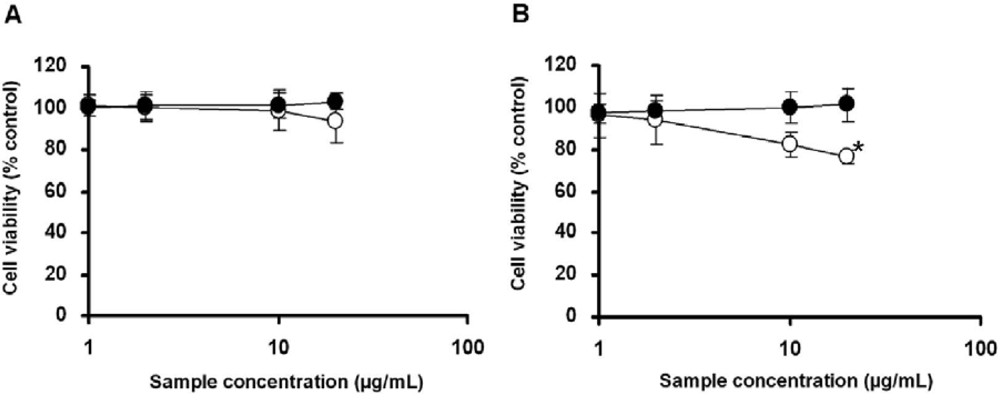
2.3. Cell Viability Test of Aggregate Titaniananoparticles
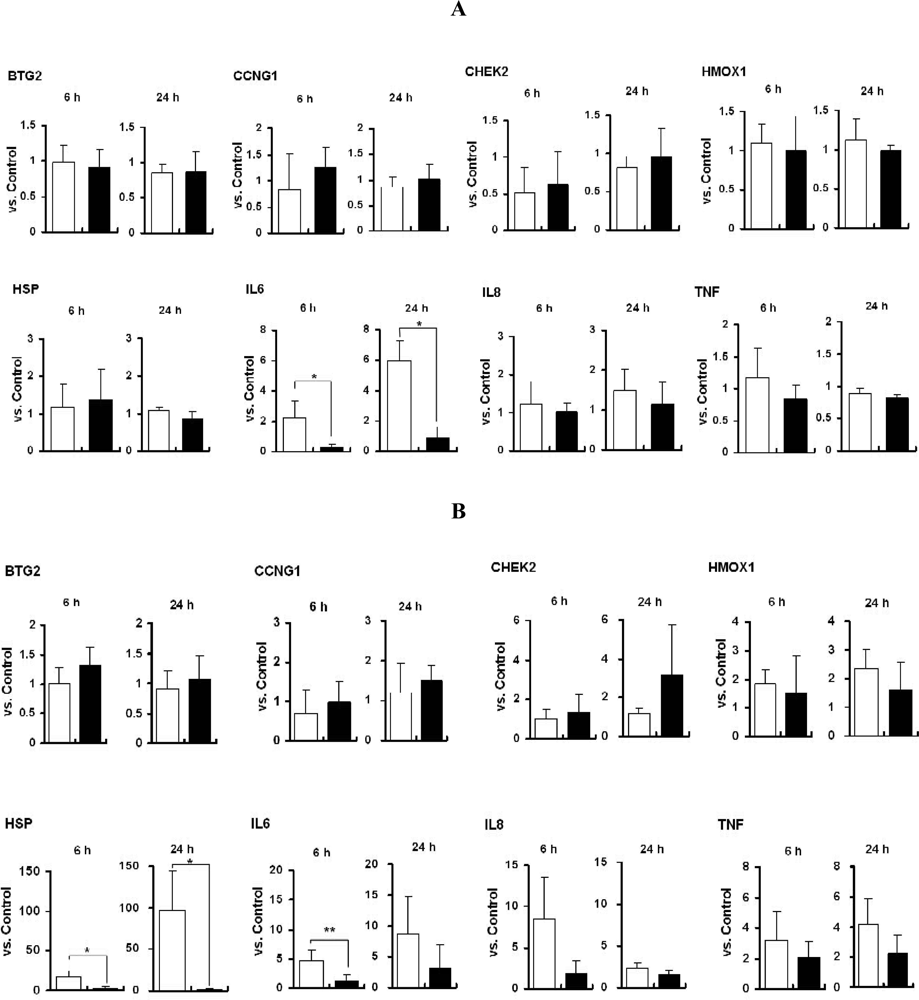
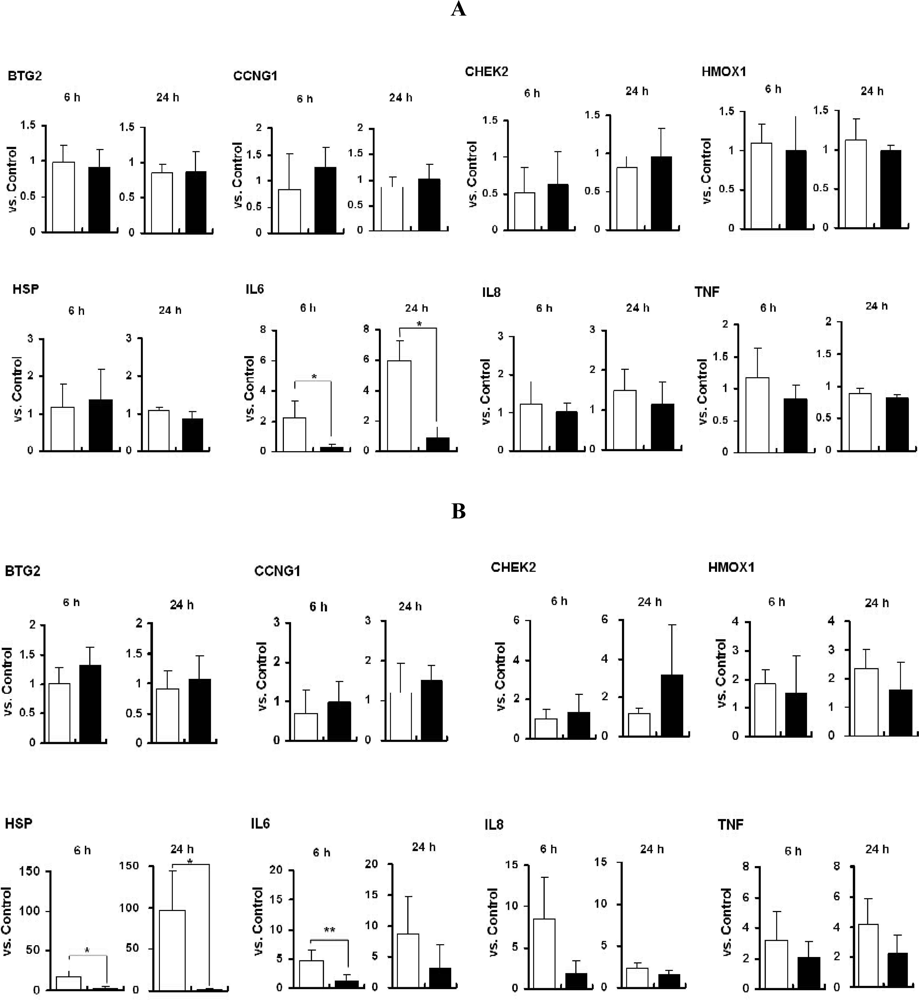
2.4. mRNA Expression of Marker Genes in Titania Particle-Exposed Cells
2.5. Discussion
3. Experimental Section
3.1. Titania Particles
3.2. Cell Cultures
3.3. Microscopic Observation
3.4. Cell Viability Test
3.5. Gene Expression Analysis
4. Conclusions
References
- Nel, A; Xia, T; Mädler, L; Li, N. Toxic potential of materials at the nanolevel. Science 2006, 311, 622–627. [Google Scholar]
- Service, RF. Nanotoxicology. Nanotechnology grows up. Science 2004, 304, 1732–1734. [Google Scholar]
- Oberdörster, G; Oberdörster, E; Oberdörster, J. Nanotoxicology: An emerging discipline evolving from studies of ultrafine particles. Environ. Health Perspect 2005, 113, 823–839. [Google Scholar]
- Poland, CA; Duffin, R; Kinloch, I; Maynard, A; Wallace, WA; Seaton, A; Stone, V; Brown, S; Macnee, W; Donaldson, K. Carbon nanotubes introduced into the abdominal cavity of mice show asbestos-like pathogenicity in a pilot study. Nat. Nanotechnol 2008, 3, 423–428. [Google Scholar]
- Takagi, A; Hirose, A; Nishimura, T; Fukumori, N; Ogata, A; Ohashi, N; Kitajima, S; Kanno, J. Induction of mesothelioma in p53+/− mouse by intraperitoneal application of multi-wall carbon nanotube. J. Toxicol. Sci 2008, 33, 105–116. [Google Scholar]
- Baveye, P; Laba, M. Aggregation and toxicology of titanium dioxide nanoparticles. Environ. Health Perspect 2008, 116, A152. [Google Scholar]
- Waters, KM; Masiello, LM; Zangar, RC; Tarasevich, BJ; Karin, NJ; Quesenberry, RD; Bandyopadhyay, S; Teeguarden, JG; Pounds, JG; Thrall, BD. Macrophage responses to silica nanoparticles are highly conserved across particle sizes. Toxicol. Sci 2009, 107, 553–569. [Google Scholar]
- Donaldson, K; Stone, V; Duffin, R; Clouter, A; Schins, R; Borm, P. The quartz hazard: Effects of surface and matrix on inflammogenic activity. J. Environ. Pathol. Toxicol. Oncol 2001, 20, S109–S118. [Google Scholar]
- Warheit, DB; Webb, TR; Colvin, VL; Reed, KL; Sayes, CM. Pulmonary bioassay studies with nanoscale and fine-quartz particles in rats: Toxicity is not dependent upon particle size but on surface characteristics. Toxicol. Sci 2007, 95, 270–280. [Google Scholar]
- Warheit, DB; Webb, TR; Sayes, CM; Colvin, VL; Reed, KL. Pulmonary instillation studies with nanoscale TiO2 rods and dots in rats: Toxicity is not dependent upon particle size and surface area. Toxicol. Sci 2006, 91, 227–236. [Google Scholar]
- Karlsson, HL; Gustafsson, J; Cronholm, P; Möller, L. Size-dependent toxicity of metal oxide particles--a comparison between nano- and micrometer size. Toxicol. Lett 2009, 188, 112–118. [Google Scholar]
- Kiang, JG; Tsokos, GC. Heat shock protein 70 kDa: Molecular biology, biochemistry, and physiology. Pharmacol. Ther 1998, 80, 183–201. [Google Scholar]
- Schlesinger, MJ. Heat shock proteins. J. Biol. Chem 1990, 265, 12111–12114. [Google Scholar]
- Wada, K; Taniguchi, A; Xu, L; Okano, T. Rapid and highly sensitive detection of cadmium chloride induced cytotoxicity using the HSP70B' promoter in live cells. Biotechnol. Bioeng 2005, 92, 410–415. [Google Scholar]
- Ito, A; Shinkai, M; Honda, H; Yoshikawa, K; Saga, S; Wakabayashi, T; Yoshida, J; Kobayashi, T. Heat shock protein 70 expression induces antitumor immunity during intracellular hyperthermia using magnetite nanoparticles. Cancer Immunol. Immunother 2003, 52, 80–88. [Google Scholar]
- Ito, A; Honda, H; Kobayashi, T. Cancer immunotherapy based on intracellular hyperthermia using magnetite nanoparticles: A novel concept of “heat-controlled necrosis” with heat shock protein expression. Cancer Immunol. Immunother 2006, 55, 320–328. [Google Scholar]
- Akira, S; Kishimoto, T. IL-6 and NF-IL6 in acute-phase response and viral infection. Immunol. Rev 1992, 127, 25–50. [Google Scholar]
- Taira, M; Nezu, T; Sasaki, M; Kimura, S; Kagiya, T; Harada, H; Narushima, T; Araki, Y. Gene expression analyses of human macrophage phagocytizing sub-micro titanium particles by allergy DNA chip (Genopal). Biomed. Mater. Eng 2009, 19, 63–70. [Google Scholar]
- Swanson, JA; Watts, C. Macropinocytosis. Trends Cell Biol 1995, 5, 424–428. [Google Scholar]
- Sansonetti, P. Phagocytosis of bacterial pathogens: Implications in the host response. Semin Immunol 2001, 13, 381–390. [Google Scholar]
- Nudejima, S; Miyazawa, K; Okuda, J; Taniguchi, A. Observation of phagocytosis of fullerene nanowhiskers by PMA-treated THP-1 cells. J. Phys 2009, 159, 1–6. [Google Scholar]
- Ishibashi, Y; Imai, S; Inouye, Y; Okano, T; Taniguchi, A. Effects of carbocisteine on sialyl-Lewis x expression in NCI-H292 cells, an airway carcinoma cell line, stimulated with tumor necrosis factor-α. Eur. J. Pharmacol 2006, 530, 223–228. [Google Scholar]
- Xu, L; Harada, H; Yokohama-Tamaki, T; Matsumoto, S; Tanaka, J; Taniguchi, A. Reuptake of extracellular amelogenin by dental epithelial cells results in increased levels of amelogenin mRNA through enhanced mRNA stabilization. J. Biol. Chem 2006, 281, 32439–32444. [Google Scholar]


© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Okuda-Shimazaki, J.; Takaku, S.; Kanehira, K.; Sonezaki, S.; Taniguchi, A. Effects of Titanium Dioxide Nanoparticle Aggregate Size on Gene Expression. Int. J. Mol. Sci. 2010, 11, 2383-2392. https://doi.org/10.3390/ijms11062383
Okuda-Shimazaki J, Takaku S, Kanehira K, Sonezaki S, Taniguchi A. Effects of Titanium Dioxide Nanoparticle Aggregate Size on Gene Expression. International Journal of Molecular Sciences. 2010; 11(6):2383-2392. https://doi.org/10.3390/ijms11062383
Chicago/Turabian StyleOkuda-Shimazaki, Junko, Saiko Takaku, Koki Kanehira, Shuji Sonezaki, and Akiyohshi Taniguchi. 2010. "Effects of Titanium Dioxide Nanoparticle Aggregate Size on Gene Expression" International Journal of Molecular Sciences 11, no. 6: 2383-2392. https://doi.org/10.3390/ijms11062383