Escherichia coli mar and acrAB Mutants Display No Tolerance to Simple Alcohols



Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results and Discussion
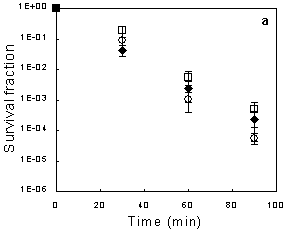
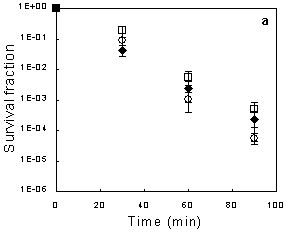
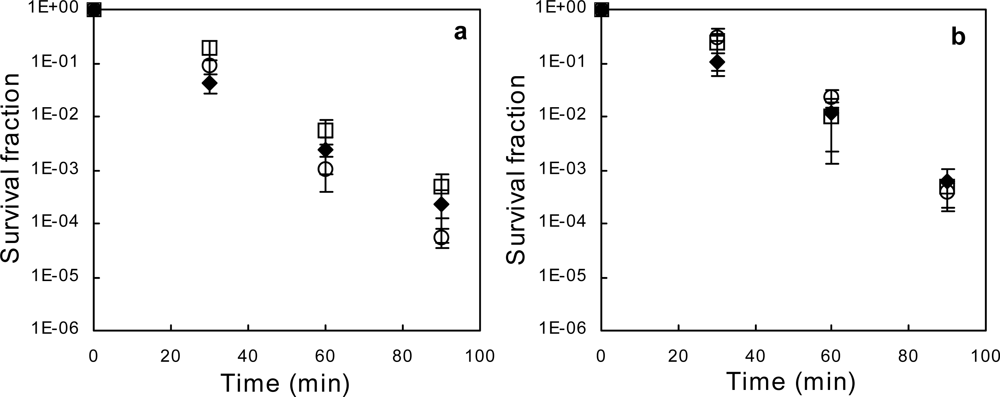
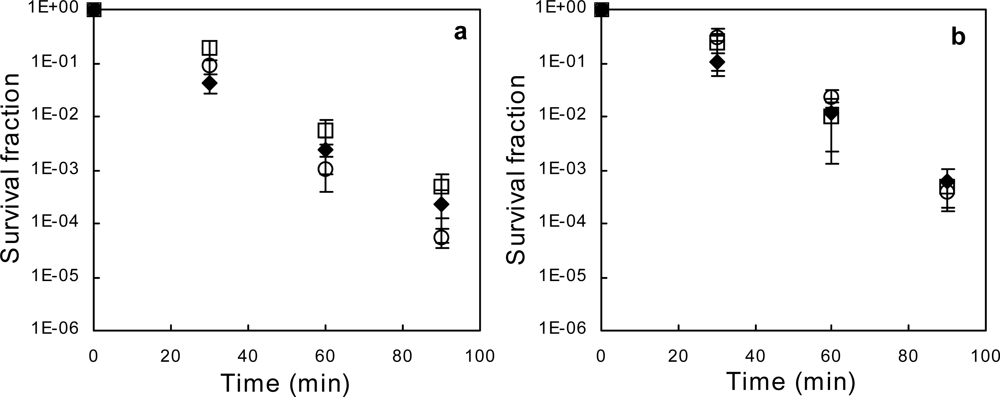
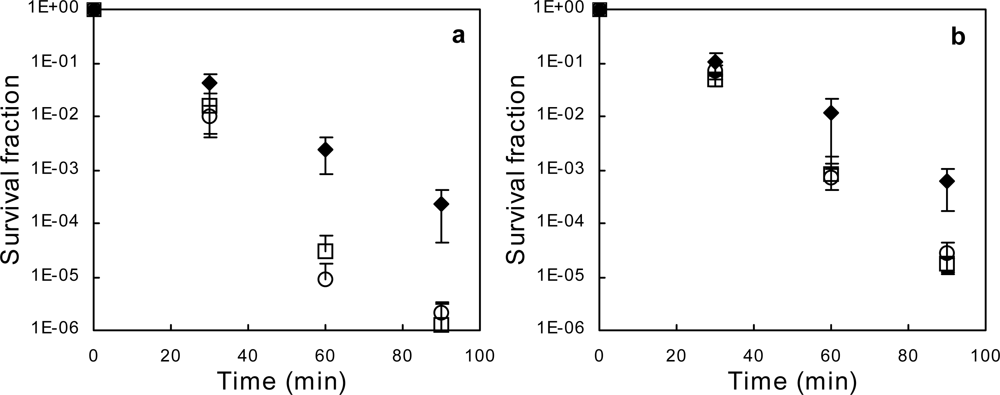
2.1. Exponential-phase Killing by Ethanol or 1-Propanol Is Unaffected by the mar Phenotype
2.2. Salicylate Increases the Killing Rate of Ethanol and 1-Propanol, Irrespective of mar Induction
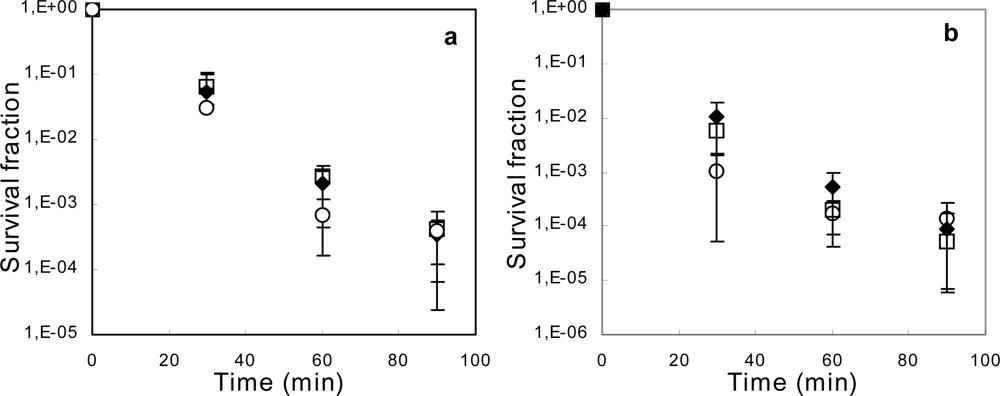
2.3. Deletion of acrAB Does Not Increase Sensitivity to Ethanol or 1-Propanol
3. Experimental Section
3.1. Bacterial Strains and Media
3.2. Solvent Killing Assay
4. Conclusions
Acknowledgments
References and Notes
- Vermuë, M; Sikkema, J; Verheul, A; Bakker, R; Tramper, J. Toxicity of homologous series of organic solvents for the Gram-positive bacteria Arthrobacter and Nocardia sp. and the Gram-negative bacteria Acinetobacter and Pseudomonas sp. Biotechnol. Bioeng 1993, 42, 747–758. [Google Scholar]
- Ingram, LO. Mechanism of lysis of Escherichia coli by ethanol and other chaotropic agents. J. Bacteriol 1981, 146, 331–336. [Google Scholar]
- Albert, A. Selective Toxicity, 7th ed; Chapman and Hall: London, UK, 1985. [Google Scholar]
- Sikkema, J; Poolman, B; Konings, WN; de Bont, JA. Effects of the membrane action of tetralin on the functional and structural properties of artificial and bacterial membranes. J. Bacteriol 1992, 174, 2986–2992. [Google Scholar]
- Sikkema, J; de Bont, JA; Poolman, B. Interactions of cyclic hydrocarbons with biological membranes. J. Biol. Chem 1994, 269, 8022–8028. [Google Scholar]
- Cartwright, CP; Veazey, FJ; Rose, AH. Effect of ethanol on activity of the plasma-membrane ATPase in, and accumulation of glycine by Saccharomyces cerevisiae. J. Gen. Microbiol 1987, 133, 857–865. [Google Scholar]
- Eaton, LC; Tedder, TF; Ingram, LO. Effects of fatty acid composition on the sensitivity of membrane functions to ethanol in Escherichia coli. Subst. Alcohol Actions Misuse 1982, 3, 77–87. [Google Scholar]
- Cohen, SP; Yan, W; Levy, SB. A multidrug resistance regulatory chromosomal locus is widespread among enteric bacteria. J. Infect. Dis 1993, 168, 484–488. [Google Scholar]
- McDermott, PF; White, DG; Podglajen, I; Alekshun, MN; Levy, SB. Multidrug resistance following expression of the Escherichia coli marA gene in Mycobacterium smegmatis. J. Bacteriol 1998, 180, 2995–2998. [Google Scholar]
- Kunonga, NI; Sobieski, RJ; Crupper, SS. Prevalence of the multiple antibiotic resistance operon (marRAB) in the genus Salmonella. FEMS Microbiol. Lett 2000, 187, 155–160. [Google Scholar]
- Cohen, SP; Hachler, H; Levy, SB. Genetic and functional analysis of the multiple antibiotic resistance (mar) locus in Escherichia coli. J. Bacteriol 1993, 175, 1484–1492. [Google Scholar]
- Barbosa, TM; Levy, SB. Differential expression of over 60 chromosomal genes in Escherichia coli by constitutive expression of MarA. J. Bacteriol 2000, 182, 3467–3474. [Google Scholar]
- Ma, D; Cook, DN; Alberti, M; Pon, NG; Nikaido, H; Hearst, JE. Molecular cloning and characterization of acrA and acrE genes of Escherichia coli. J. Bacteriol 1993, 175, 6299–6313. [Google Scholar]
- Ma, D; Alberti, M; Lynch, C; Nikaido, H; Hearst, JE. The local repressor AcrR plays a modulating role in the regulation of acrAB genes of Escherichia coli by global stress signals. Mol. Microbiol 1996, 19, 101–112. [Google Scholar]
- Thanassi, DG; Cheng, LW; Nikaido, H. Active efflux of bile salts by Escherichia coli. J. Bacteriol 1997, 179, 2512–2518. [Google Scholar]
- Moken, MC; McMurry, LM; Levy, SB. Selection of multiple-antibiotic-resistant (mar) mutants of Escherichia coli by using the disinfectant pine oil: roles of the mar and acrAB loci. Antimicrob. Agents Chemother 1997, 41, 2770–2772. [Google Scholar]
- McMurry, LM; Oethinger, M; Levy, SB. Overexpression of marA, soxS, or acrAB produces resistance to triclosan in laboratory and clinical strains of Escherichia coli. FEMS Microbiol. Lett 1998, 166, 305–309. [Google Scholar]
- Hachler, H; Cohen, SP; Levy, SB. marA, a regulated locus which controls expression of chromosomal multiple antibiotic resistance in Escherichia coli. J. Bacteriol 1991, 173, 5532–5538. [Google Scholar]
- Okusu, H; Ma, D; Nikaido, H. AcrAB efflux pump plays a major role in the antibiotic resistance phenotype of Escherichia coli multiple-antibiotic-resistance (Mar) mutants. J. Bacteriol 1996, 178, 306–308. [Google Scholar]
- Martin, RG; Rosner, JL. Binding of purified multiple antibiotic-resistance repressor protein (MarR) to mar operator sequences. Proc. Natl. Acad. Sci. USA 1995, 92, 5456–5460. [Google Scholar]
- Alekshun, MN; Levy, SB. Alteration of the repressor activity of MarR, the negative regulator of the Escherichia coli marRAB locus, by multiple chemicals in vitro. J. Bacteriol 1999, 181, 4669–4672. [Google Scholar]
- Cohen, SP; Levy, SB; Foulds, J; Rosner, JL. Salicylate induction of antibiotic resistance in Escherichia coli: Activation of the mar operon and a mar-independent pathway. J. Bacteriol 1993, 175, 7856–7862. [Google Scholar]
- Brooun, A; Tomashek, JJ; Lewis, K. Purification and ligand binding of EmrR, a regulator of a multidrug transporter. J. Bacteriol 1999, 181, 5131–5133. [Google Scholar]
- Aono, R; Kobayashi, M; Nakajima, H; Kobayashi, H. A close correlation between improvement of organic solvent tolerance levels and alteration of resistance toward low levels of multiple antibiotics in Escherichia coli. Biosci. Biotechnol. Biochem 1995, 59, 213–218. [Google Scholar]
- Asako, H; Nakajima, H; Kobayashi, K; Kobayashi, M; Aono, R. Organic solvent tolerance and antibiotic resistance increased by overexpression of marA in Escherichia coli. Appl. Environ. Microbiol 1997, 63, 1428–1433. [Google Scholar]
- Oethinger, M; Kern, WV; Goldman, JD; Levy, SB. Association of organic solvent tolerance and fluoroquinolone resistance in clinical isolates of Escherichia coli. J. Antimicrob. Chemother 1998, 41, 111–114. [Google Scholar]
- George, AM; Levy, SB. Amplifiable resistance to tetracycline, chloramphenicol, and other antibiotics in Escherichia coli: involvement of a non-plasmid-determined efflux of tetracycline. J. Bacteriol 1983, 155, 531–540. [Google Scholar]
- Oethinger, M; Kern, WV; Jellen-Ritter, AS; McMurry, LM; Levy, SB. Ineffectiveness of topoisomerase mutations in mediating clinically significant fluoroquinolone resistance in Escherichia coli in the absence of the AcrAB efflux pump. Antimicrob. Agents Chemother 2000, 44, 10–13. [Google Scholar]
- Maneewannakul, K; Levy, SB. Identification for mar mutants among quinolone-resistant clinical isolates of Escherichia coli. Antimicrob. Agents Chemother 1996, 40, 1695–1698. [Google Scholar]
- Farewell, A; Kvint, K; Nystrom, T. uspB, a new sigmaS-regulated gene in Escherichia coli which is required for stationary-phase resistance to ethanol. J. Bacteriol 1998, 180, 6140–6147. [Google Scholar]
- Fried, VA; Novick, A. Organic solvents as probes for the structure and function of the bacterial membrane: effects of ethanol on the wild type and an ethanol-resistant mutant of Escherichia coli K-12. J. Bacteriol 1973, 114, 239–248. [Google Scholar]
- Ingram, LO; Vreeland, NS. Differential effects of ethanol and hexanol on the Escherichia coli cell envelope. J. Bacteriol 1980, 144, 481–488. [Google Scholar]
- Jordan, SL; Glover, J; Malcolm, L; Thomson-Carter, FM; Booth, IR; Park, SF. Augmentation of killing of Escherichia coli O157 by combinations of lactate, ethanol, and low-pH conditions. Appl. Environ. Microbiol 1999, 65, 1308–1311. [Google Scholar]
- Aumercier, M; Murray, DM; Rosner, JL. Potentiation of susceptibility to aminoglycosides by salicylate in Escherichia coli. Antimicrob. Agents Chemother 1990, 34, 786–791. [Google Scholar]
- Domenico, P; Hopkins, T; Schoch, PE; Cunha, BA. Potentiation of aminoglycoside inhibition and reduction of capsular polysaccharide production in Klebsiella pneumoniae by sodium salicylate. J. Antimicrob. Chemother 1990, 25, 903–914. [Google Scholar]
- Damper, PD; Epstein, W. Role of the membrane potential in bacterial resistance to aminoglycoside antibiotics. Antimicrob. Agents Chemother 1981, 20, 803–808. [Google Scholar]
- Bryan, LE; Kwan, S. Roles of ribosomal binding, membrane potential, and electron transport in bacterial uptake of streptomycin and gentamicin. Antimicrob. Agents Chemother 1983, 23, 835–845. [Google Scholar]
- Tsukagoshi, N; Aono, R. Entry into and release of solvents by Escherichia coli in an organic-aqueous two-liquid-phase system and substrate specificity of the AcrAB-TolC solvent-extruding pump. J. Bacteriol 2000, 182, 4803–4810. [Google Scholar]
- Ferhat, M; Atlan, D; Vianney, A; Lazzaroni, JC; Doublet, P; Gilbert, C. The TolC protein of Legionella pneumophila plays a major role in multi-drug resistance and the early steps of host invasion. PLOS one 2009, 4, e7732. [Google Scholar]
- Ricci, V; Piddock, LJV. Only for substrate antibiotics are a functional AcrAB-TolC efflux pump and RamA required to select multidrug-resistant Salmonella Typhimurium. J. Antimicrob. Chemother 2009, 63, 654–657. [Google Scholar]
- Olofsson, L; Ankarloo, J; Andersson, PO; Nicholls, IA. Bacteriophage viability in organic solvents. Chem. Biol 2001, 8, 661–671. [Google Scholar]
- Berglund, J; Nicholls, IA; Lindbladh, C; Mosbach, K. Selection of phage display peptides with affinity for a yohimbine imprinted methacrylate polymer. Anal. Commun 1998, 35, 3–7. [Google Scholar]
- Söderberg, P; Olofsson, L; Ankarloo, J; Nicholls, IA. Phage display screening in low dielectric media. J. Mol. Recognit 2008, 29, 329–336. [Google Scholar]
© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Ankarloo, J.; Wikman, S.; Nicholls, I.A. Escherichia coli mar and acrAB Mutants Display No Tolerance to Simple Alcohols. Int. J. Mol. Sci. 2010, 11, 1403-1412. https://doi.org/10.3390/ijms11041403
Ankarloo J, Wikman S, Nicholls IA. Escherichia coli mar and acrAB Mutants Display No Tolerance to Simple Alcohols. International Journal of Molecular Sciences. 2010; 11(4):1403-1412. https://doi.org/10.3390/ijms11041403
Chicago/Turabian StyleAnkarloo, Jonas, Susanne Wikman, and Ian A. Nicholls. 2010. "Escherichia coli mar and acrAB Mutants Display No Tolerance to Simple Alcohols" International Journal of Molecular Sciences 11, no. 4: 1403-1412. https://doi.org/10.3390/ijms11041403