The Fibrillar Collagen Family

Abstract
:
1. Introduction
2. Fibrillar Collagen Family
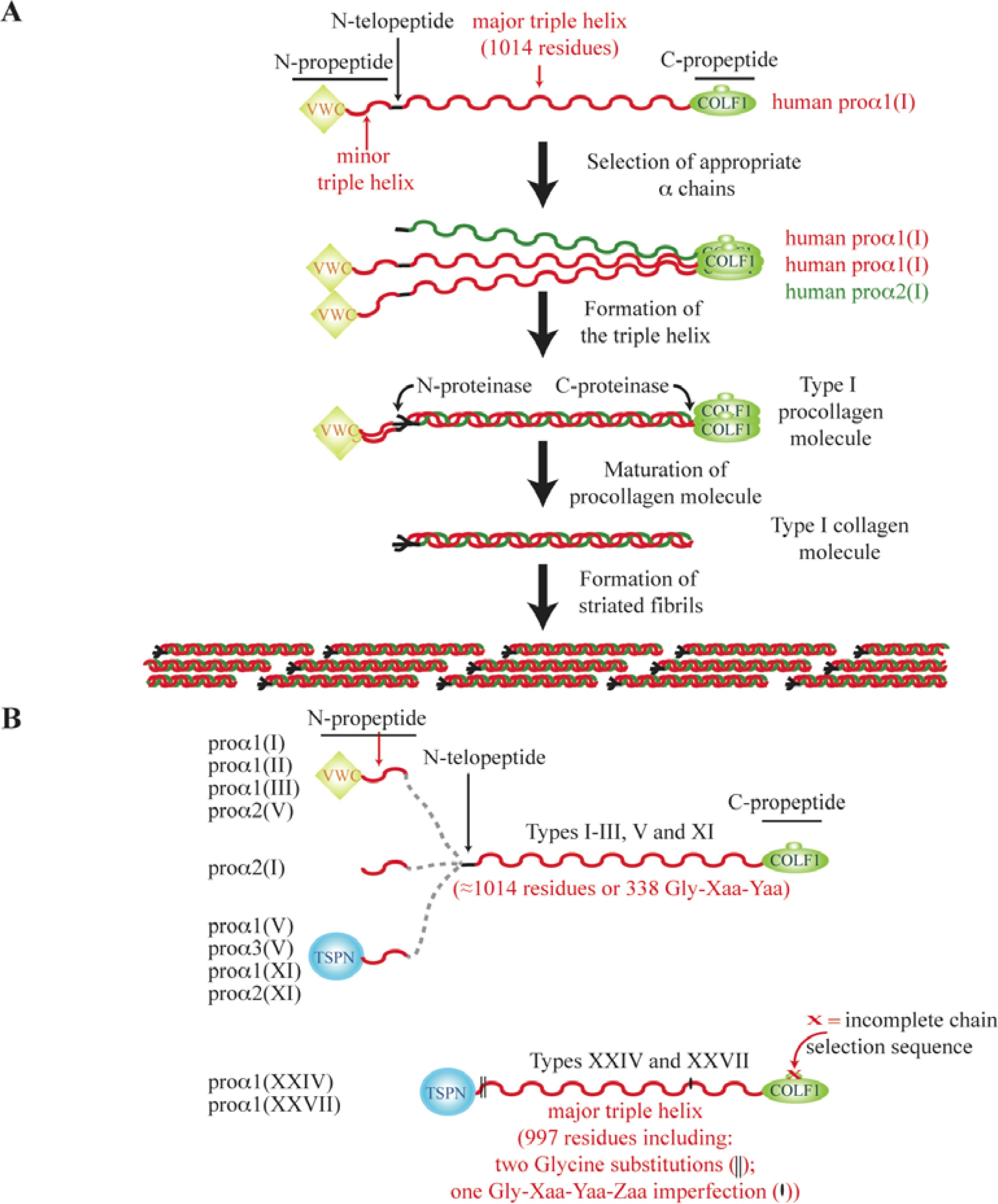
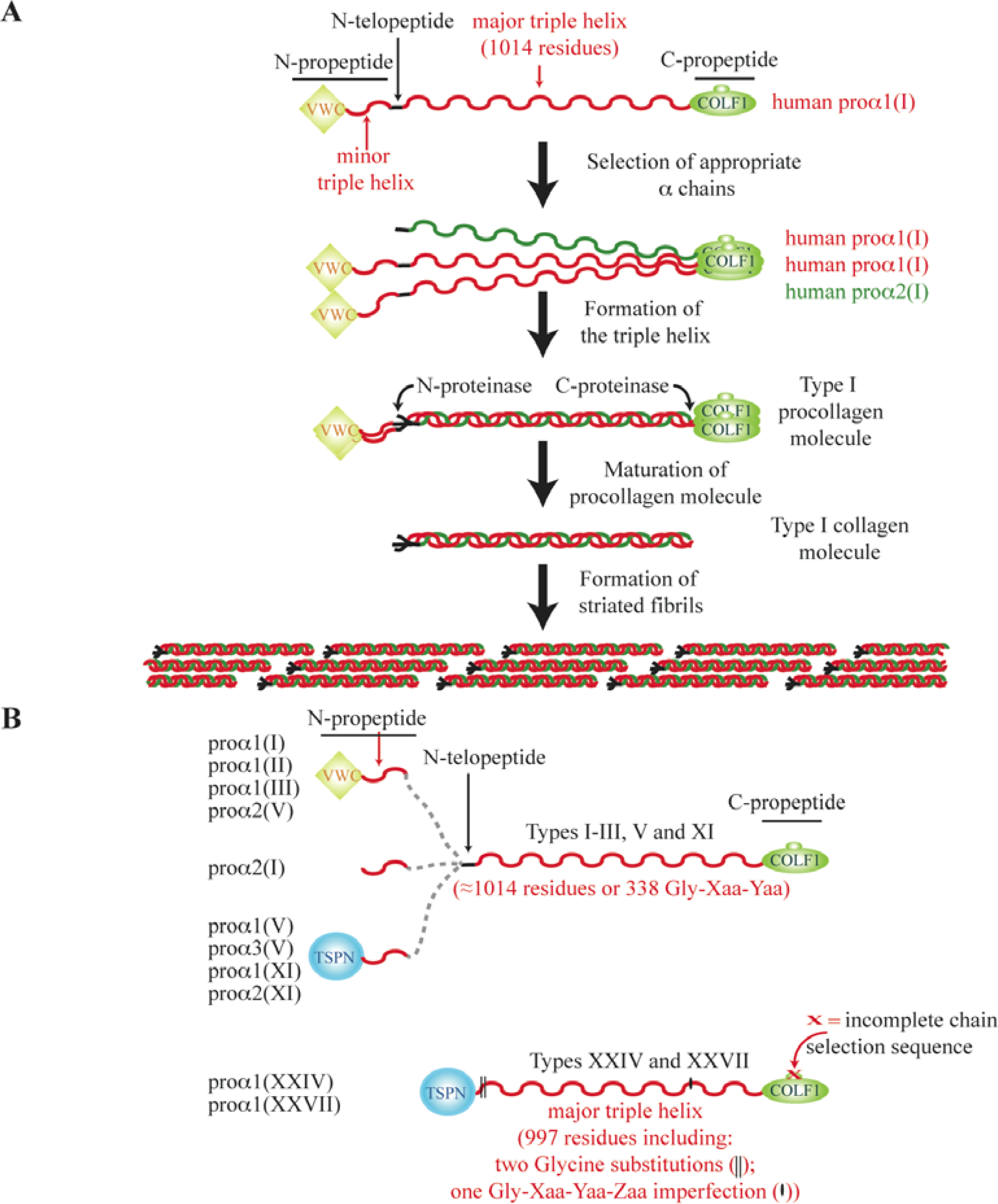
3. The N-Propeptide Region of Fibrillar Procollagens
4. The C-Propeptide of Fibrillar Collagens
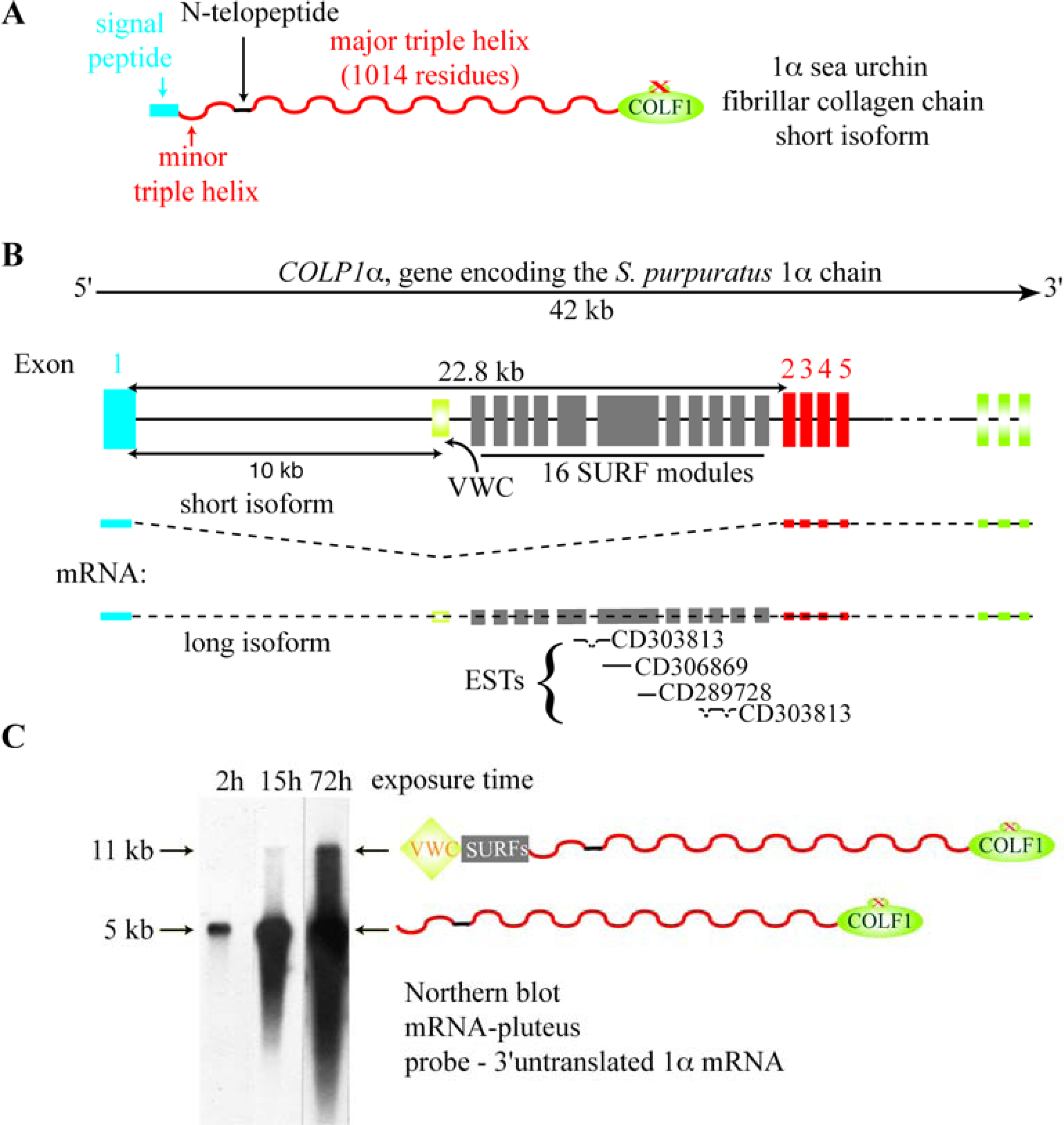
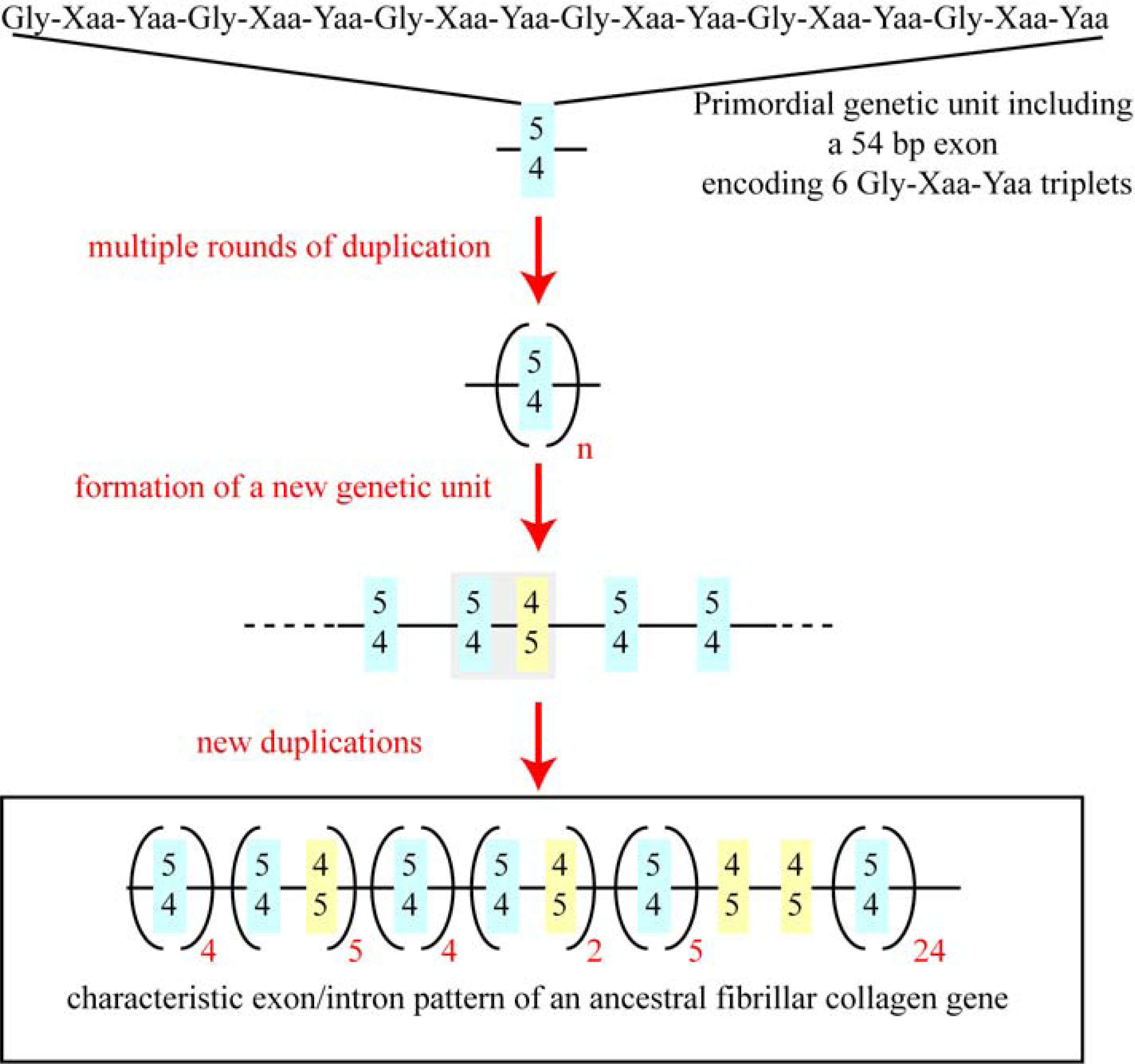
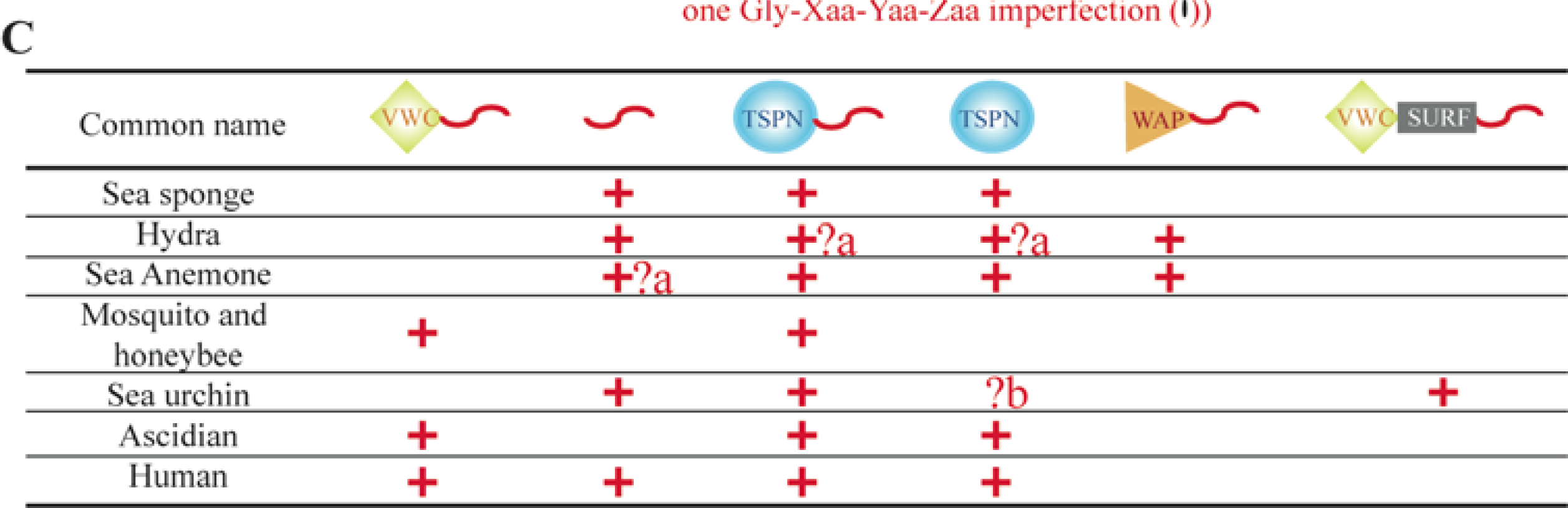
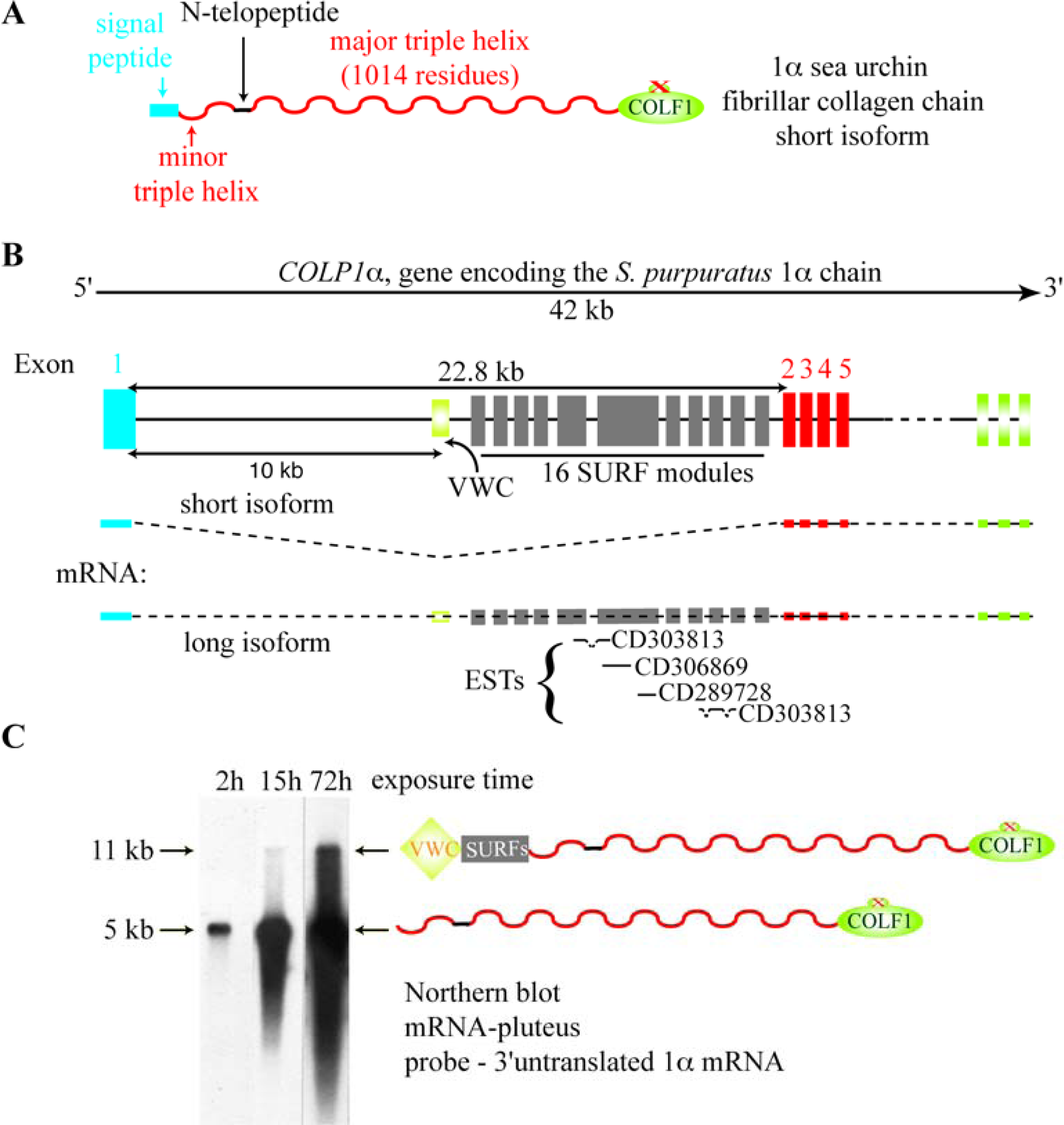
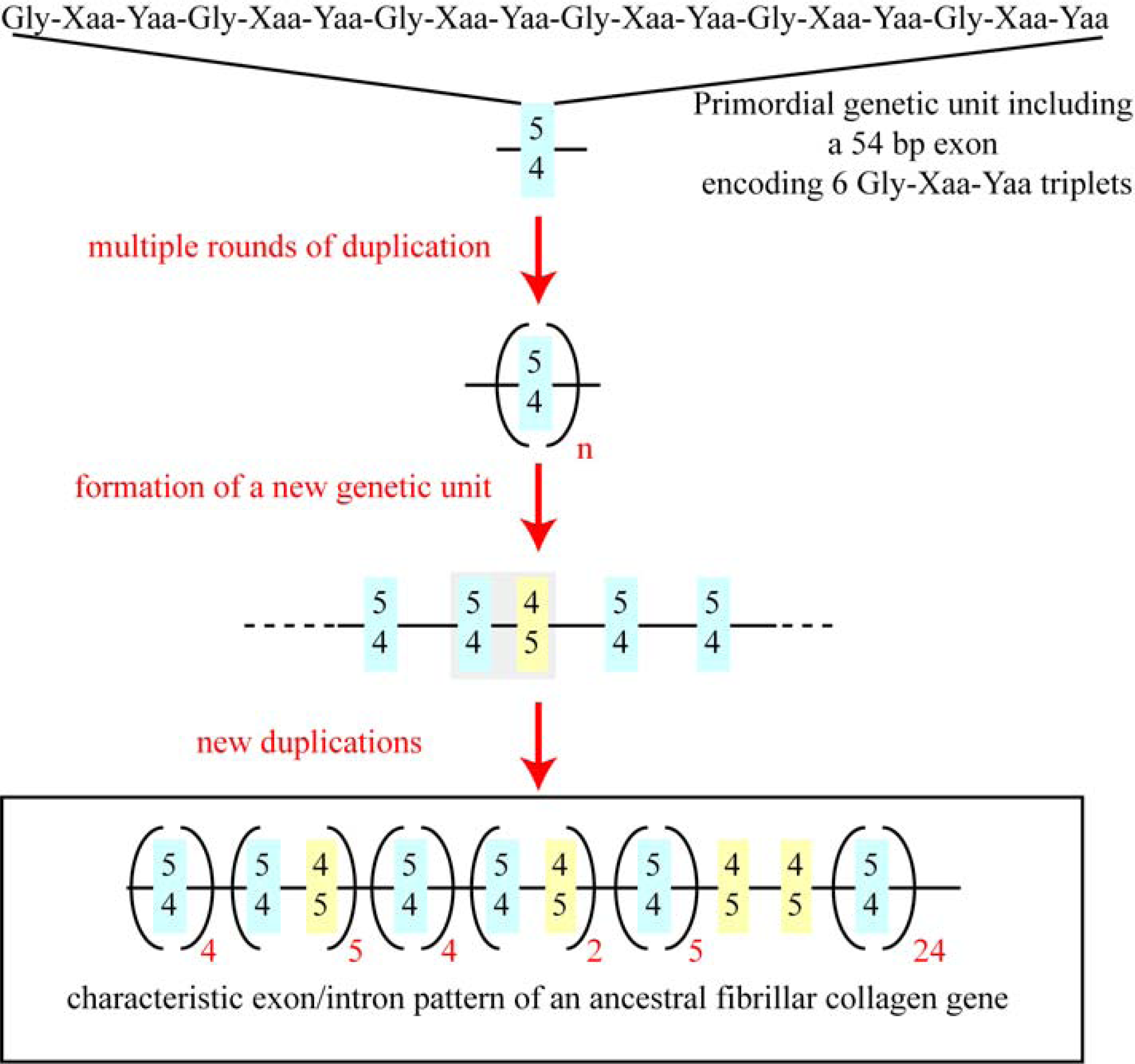
5. The Major Triple Helical Sequences and the Formation of an Ancestral Gene
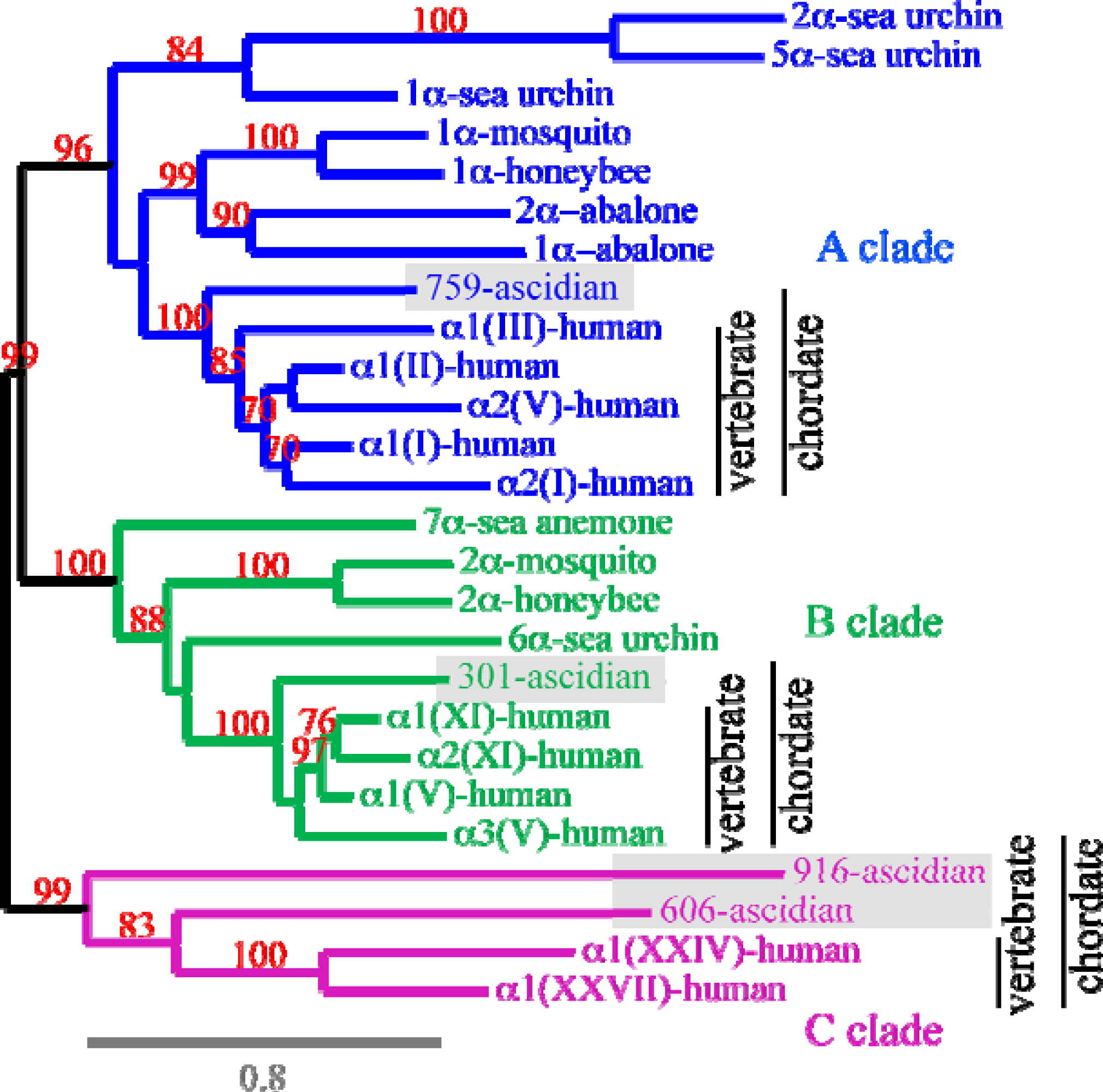
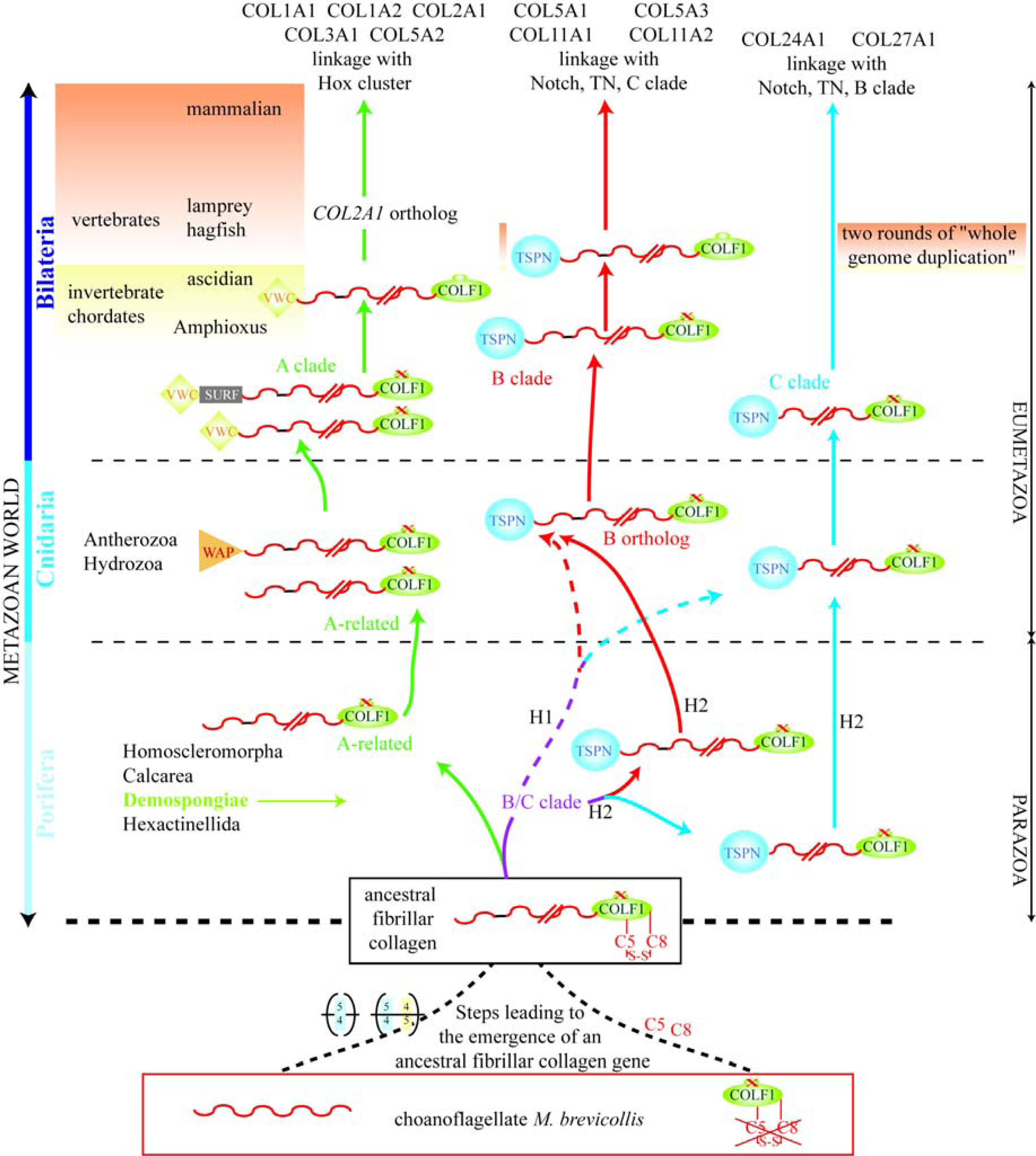
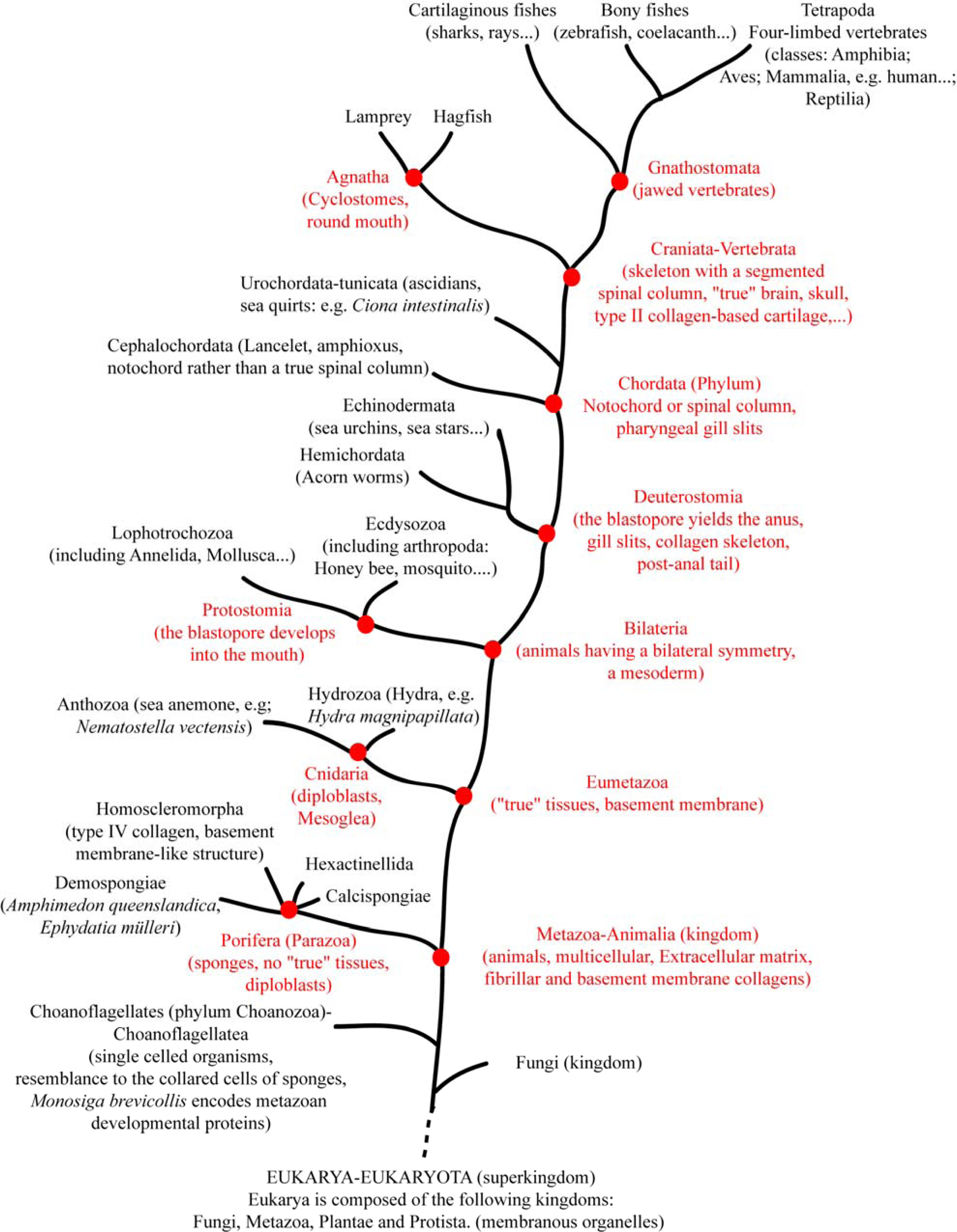
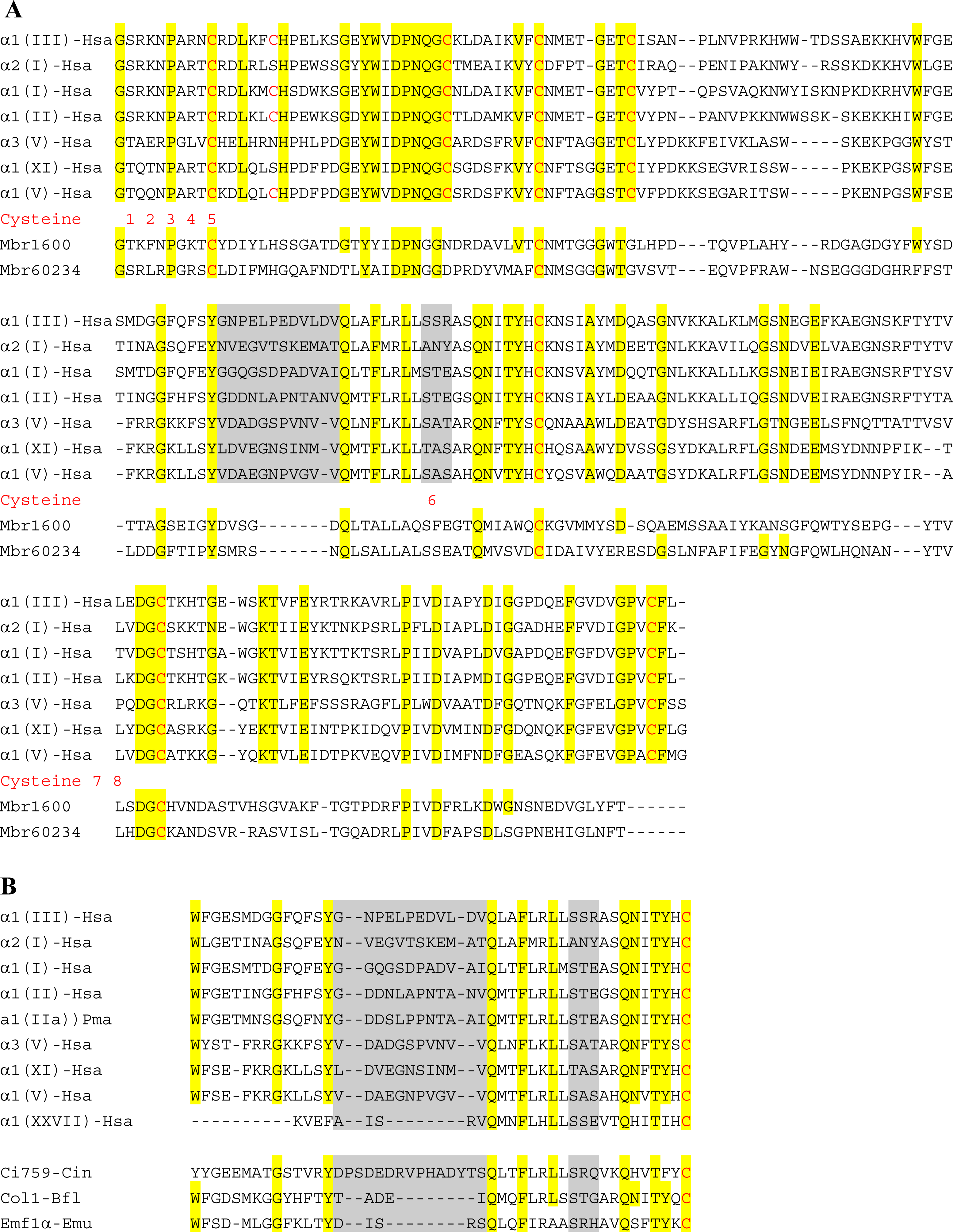
6. Using Triple Helical Sequences to Decipher Fibrillar Collagen Evolution
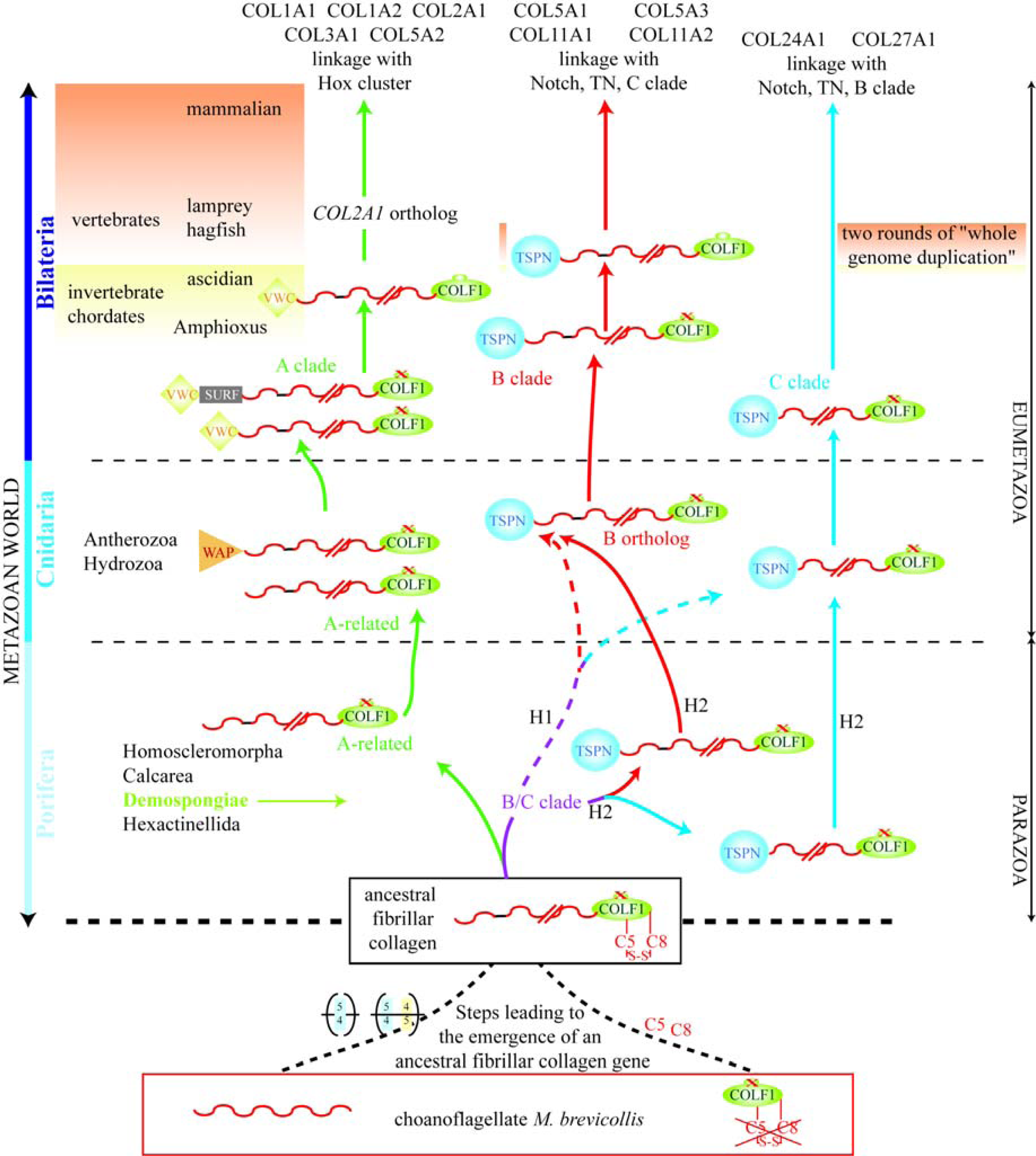
7. Suggested Evolutionary Model for the Fibrillar Collagen Family
8. Conclusions
Acknowledgments
References and Notes
- Myllyharju, J; Kivirikko, KI. Collagens, modifying enzymes and their mutations in humans, flies and worms. Trends Genet 2004, 20, 33–43. [Google Scholar]
- Exposito, JY; Cluzel, C; Garrone, R; Lethias, C. Evolution of collagens. Anat. Rec 2002, 268, 302–316. [Google Scholar]
- Rasmussen, M; Jacobsson, M; Björck, L. Genome-based identification and analysis of collagen-related structural motifs in bacterial and viral proteins. J. Biol. Chem 2003, 278, 32313–32316. [Google Scholar]
- Celerin, M; Ray, JM; Schisler, NJ; Day, AW; Stetler-Stevenson, WG; Laudenbach, DE. Fungal fimbriae are composed of collagen. EMBO J 1996, 15, 4445–4453. [Google Scholar]
- King, N; Westbrook, MJ; Young, SL; Kuo, A; Abedin, M; Chapman, J; Fairclough, S; Hellsten, U; Isogai, Y; Letunic, I; Marr, M; Pincus, D; Putnam, N; Rokas, A; Wright, KJ; Zuzow, R; Dirks, W; Good, M; Goodstein, D; Lemons, D; Li, W; Lyons, JB; Morris, A; Nichols, S; Richter, DJ; Salamov, A; Sequencing, JGI; Bork, P; Lim, WA; Manning, G; Miller, WT; McGinnis, W; Shapiro, H; Tjian, R; Grigoriev, IV; Rokhsar, D. The genome of the choanoflagellate Monosiga brevicollis and the origin of metazoans. Nature 2008, 451, 783–788. [Google Scholar]
- Acton, S; Resnick, D; Freeman, M; Ekkel, Y; Ashkenas, J; Krieger, M. The collagenous domains of macrophage scavenger receptors and complement component C1q mediate their similar, but not identical, binding specificities for polyanionic ligands. J. Biol. Chem 1993, 268, 3530–3537. [Google Scholar]
- Heino, J. The collagen family members as cell adhesion proteins. Bioessays 2007, 29, 1001–1010. [Google Scholar]
- Söderhäll, C; Marenholz, I; Kerscher, T; Rüschendorf, F; Esparza-Gordillo, J; Worm, M; Gruber, C; Mayr, G; Albrecht, M; Rohde, K; Schulz, H; Wahn, U; Hubner, N; Lee, YA. Variants in a novel epidermal collagen gene (COL29A1) are associated with atopic dermatitis. PLoS Biol 2007, 5, e242. [Google Scholar]
- Exposito, JY; Garrone, R. Characterization of a fibrillar collagen gene in sponges reveals the early evolutionary appearance of two collagen gene families. Proc. Natl. Acad. Sci. USA 1990, 87, 6669–6673. [Google Scholar]
- Boute, N; Exposito, JY; Boury-Esnault, N; Vacelet, J; Noro, N; Miyazaki, K; Yoshizato, K; Garrone, R. Type IV collagen in sponges, the missing link in basement membrane ubiquity. Biol. Cell 1996, 88, 37–44. [Google Scholar]
- Aouacheria, A; Geourjon, C; Aghajari, N; Navratil, V; Deléage, G; Lethias, C; Exposito, JY. Insights into early extracellular matrix evolution: spongin short chain collagen-related proteins are homologous to basement membrane type IV collagens and form a novel family widely distributed in invertebrates. Mol. Biol. Evol 2006, 23, 2288–2302. [Google Scholar]
- Wada, H; Okuyama, M; Satoh, N; Zhang, S. Molecular evolution of fibrillar collagen in chordates, with implications for the evolution of vertebrate skeletons and chordate phylogeny. Evol. Dev 2006, 8, 370–377. [Google Scholar]
- Wick, G; Kalischnig, G; Maurer, H; Mayerl, C; Müller, PU. Really old-palaeoimmunology: immunohistochemical analysis of extracellular matrix proteins in historic and pre-historic material. Exp. Gerontol 2001, 36, 1565–1579. [Google Scholar]
- Franc, S; Marzin, E; Boutillon, MM; Lafont, R; Lechéne de la Porte, P; Herbage, D. Immunohistochemical and biochemical analyses of 20,000–25,000-year-old fossil cartilage. Eur. J. Biochem 1995, 234, 125–131. [Google Scholar]
- Asara, JM; Schweitzer, MH; Freimark, LM; Phillips, M; Cantley, LC. Protein sequences from mastodon and Tyrannosaurus rex revealed by mass spectrometry. Science 2007, 316, 280–285. [Google Scholar]
- Schweitzer, MH; Zheng, W; Organ, CL; Avci, R; Suo, Z; Freimark, LM; Lebleu, VS; Duncan, MB; Vander Heiden, MG; Neveu, JM; Lane, WS; Cottrell, JS; Horner, JR; Cantley, LC; Kalluri, R; Asara, JM. Biomolecular characterization and protein sequences of the Campanian hadrosaur B. canadensis. Science 2009, 324, 626–631. [Google Scholar]
- Bern, M; Phinney, BS; Goldberg, D. Reanalysis of Tyrannosaurus rex Mass Spectra. J. Proteome Res 2009, 8, 4328–4332. [Google Scholar]
- Koch, M; Laub, F; Zhou, P; Hahn, RA; Tanaka, S; Burgeson, RE; Gerecke, DR; Ramirez, F; Gordon, MK. Collagen XXIV, a vertebrate fibrillar collagen with structural features of invertebrate collagens: selective expression in developing cornea and bone. J. Biol. Chem 2003, 278, 43236–43244. [Google Scholar]
- Boot-Handford, RP; Tuckwell, DS; Plumb, DA; Rock, CF; Poulsom, R. A novel and highly conserved collagen [pro(α)1(XXVII)] with a unique expression pattern and unusual molecular characteristics establishes a new clade within the vertebrate fibrillar collagen family. J. Biol. Chem 2003, 278, 31067–31077. [Google Scholar]
- Pace, JM; Corrado, M; Missero, C; Byers, PH. Identification, characterization and expression analysis of a new fibrillar collagen gene, COL27A1. Matrix Biol 2003, 22, 3–14. [Google Scholar]
- Kleman, JP; Aeschlimann, D; Paulsson, M; van der Rest, M. Transglutaminase-catalyzed cross-linking of fibrils of collagen V/XI in A204 rhabdomyosarcoma cells. Biochemistry 1995, 34, 13768–13775. [Google Scholar]
- Burgeson, RE; Hollister, DW. Collagen heterogeneity in human cartilage: identification of several new collagen chains. Biochem. Biophys. Res. Commun 1979, 87, 1124–1131. [Google Scholar]
- Reese, CA; Mayne, R. Minor collagens of chicken hyaline cartilage. Biochemistry 1981, 20, 5443–5448. [Google Scholar]
- Lees, JF; Tasab, M; Bulleid, NJ. Identification of the molecular recognition sequence which determines the type-specific assembly of procollagen. EMBO J 1997, 16, 908–916. [Google Scholar]
- Aouacheria, A; Cluzel, C; Lethias, C; Gouy, M; Garrone, R; Exposito, JY. Invertebrate data predict an early emergence of vertebrate fibrillar collagen clades and an anti-incest model. J. Biol. Chem 2004, 279, 47711–47719. [Google Scholar]
- Thom, JR; Morris, NP. Biosynthesis and proteolytic processing of type XI collagen in embryonic chick sterna. J. Biol. Chem 1991, 266, 7262–7269. [Google Scholar]
- Linsenmayer, TF; Gibney, E; Igoe, F; Gordon, MK; Fitch, JM; Fessler, LI; Birk, DE. Type V collagen: molecular structure and fibrillar organization of the chicken α1(V) NH2-terminal domain, a putative regulator of corneal fibrillogenesis. J. Cell Biol 1993, 121, 1181–1189. [Google Scholar]
- Plumb, DA; Dhir, V; Mironov, A; Ferrara, L; Poulsom, R; Kadler, KE; Thornton, DJ; Briggs, MD; Boot-Handford, RP. Collagen XXVII is developmentally regulated and forms thin fibrillar structures distinct from those of classical vertebrate fibrillar collagens. J. Biol. Chem 2007, 282, 12791–12795. [Google Scholar]
- Hjorten, R; Hansen, U; Underwood, RA; Telfer, HE; Fernandes, RJ; Krakow, D; Sebald, E; Wachsmann-Hogiu, S; Bruckner, P; Jacquet, R; Landis, WJ; Byers, PH; Pace, JM. Type XXVII collagen at the transition of cartilage to bone during skeletogenesis. Bone 2007, 41, 535–542. [Google Scholar]
- Exposito, JY; Larroux, C; Cluzel, C; Valcourt, U; Lethias, C; Degnan, BM. Demosponge and sea anemone fibrillar collagen diversity reveals the early emergence of A/C clades and the maintenance of the modular structure of type V/XI collagens from sponge to human. J. Biol. Chem 2008, 283, 28226–28235. [Google Scholar]
- Exposito, JY; D’Alessio, M; Ramirez, F. Novel amino-terminal propeptide configuration in a fibrillar procollagen undergoing alternative splicing. J. Biol. Chem 1992, 267, 17404–17408. [Google Scholar]
- Cluzel, C; Lethias, C; Garrone, R; Exposito, JY. Distinct maturations of N-propeptide domains in fibrillar procollagen molecules involved in the formation of heterotypic fibrils in adult sea urchin collagenous tissues. J. Biol. Chem 2004, 279, 9811–9817. [Google Scholar]
- Exposito, JY; D’Alessio, M; Solursh, M; Ramirez, F. Sea urchin collagen evolutionarily homologous to vertebrate pro-α2(I) collagen. J. Biol. Chem 1992, 267, 15559–15562. [Google Scholar]
- Suzuki, HR; Reiter, RS; D’Alessio, M; Di Liberto, M; Ramirez, F; Exposito, JY; Gambino, R; Solursh, M. Comparative analysis of fibrillar and basement membrane collagen expression in embryos of the sea urchin, Strongylocentrotus purpuratus. Zoolog. Sci 1997, 14, 449–454. [Google Scholar]
- Dion, AS; Myers, JC. COOH-terminal propeptides of the major human procollagens. Structural, functional and genetic comparisons. J. Mol. Biol 1987, 193, 127–143. [Google Scholar]
- Boot-Handford, RP; Tuckwell, DS. Fibrillar collagen: the key to vertebrate evolution? A tale of molecular incest. Bioessays 2003, 25, 142–151. [Google Scholar]
- Yamada, Y; Avvedimento, VE; Mudryj, M; Ohkubo, H; Vogeli, G; Irani, M; Pastan, I; de Crombrugghe, B. The collagen gene: evidence for its evolutinary assembly by amplification of a DNA segment containing an exon of 54 bp. Cell 1980, 22, 887–892. [Google Scholar]
- Exposito, JY; van der Rest, M; Garrone, R. The complete intron/exon structure of Ephydatia mülleri fibrillar collagen gene suggests a mechanism for the evolution of an ancestral gene module. J. Mol. Evol 1993, 37, 254–259. [Google Scholar]
- Takahara, K; Hoffman, GG; Greenspan, DS. Complete structural organization of the human α1(V) collagen gene (COL5A1): divergence from the conserved organization of other characterized fibrillar collagen genes. Genomics 1995, 29, 588–597. [Google Scholar]
- Vuoristo, MM; Pihlajamaa, T; Vandenberg, P; Prockop, DJ; Ala-Kokko, L. The human COL11A2 gene structure indicates that the gene has not evolved with the genes for the major fibrillar collagens. J. Biol. Chem 1995, 270, 22873–22881. [Google Scholar]
- Exposito, JY; Cluzel, C; Lethias, C; Garrone, R. Tracing the evolution of vertebrate fibrillar collagens from an ancestral α chain. Matrix Biol 2000, 19, 275–279. [Google Scholar]
- Miura, S; Kimura, S. Jellyfish mesogloea collagen. Characterization of molecules as α1α2α3 heterotrimers. J. Biol. Chem 1985, 260, 15352–15356. [Google Scholar]
- Tillet, E; Franc, JM; Franc, S; Garrone, R. The evolution of fibrillar collagens: a sea-pen collagen shares common features with vertebrate type V collagen. Comp. Biochem. Physiol. B: Biochem. Mol. Biol 1996, 113, 239–246. [Google Scholar]
- Sicot, FX; Exposito, JY; Masselot, M; Garrone, R; Deutsch, J; Gaill, F. Cloning of an annelid fibrillar-collagen gene and phylogenetic analysis of vertebrate and invertebrate collagens. Eur. J. Biochem 1997, 246, 50–58. [Google Scholar]
- Yandell, M; Mungall, CJ; Smith, C; Prochnik, S; Kaminker, J; Hartzell, G; Lewis, S; Rubin, GM. Large-scale trends in the evolution of gene structures within 11 animal genomes. PLoS Comput. Biol 2006, 2, e15. [Google Scholar]
- Roy, SW; Gilbert, W. Resolution of a deep animal divergence by the pattern of intron conservation. Proc. Natl. Acad. Sci. USA 2005, 102, 4403–4408. [Google Scholar]
- Csurös, M; Rogozin, IB; Koonin, EV. Extremely intron-rich genes in the alveolate ancestors inferred with a flexible maximum-likelihood approach. Mol. Biol. Evol 2008, 25, 903–911. [Google Scholar]
- Irimia, M; Roy, SW. Spliceosomal introns as tools for genomic and evolutionary analysis. Nucleic Acids Res 2008, 36, 1703–1712. [Google Scholar]
- Chevenet, F; Brun, C; Banuls, AL; Jacq, B; Chisten, R. TreeDyn: Towards dynamic graphics and annotations for analyses of trees. BMC Bioinformatics 2006, 7. [Google Scholar]
- Zhang, X; Boot-Handford, RP; Huxley-Jones, J; Forse, LN; Mould, AP; Robertson, DL; Li, L; Athiyal, M; Sarras, MP, Jr. The collagens of hydra provide insight into the evolution of metazoan extracellular matrices. J. Biol. Chem 2007, 282, 6792–6802. [Google Scholar]
- Rychel, AL; Smith, SE; Shimamoto, HT; Swalla, BJ. Evolution and development of the chordates: collagen and pharyngeal cartilage. Mol. Biol. Evol 2006, 23, 541–549. [Google Scholar]
- DeSalle, R; Schierwater, B. An even “newer” animal phylogeny. Bioessays 2008, 30, 1043–1047. [Google Scholar]
- Zhang, G; Cohn, MJ. Genome duplication and the origin of the vertebrate skeleton. Curr. Opin. Genet. Dev 2008, 18, 387–393. [Google Scholar]
- Degnan, BM; Vervoort, M; Larroux, C; Richards, GS. Early evolution of metazoan transcription factors. Curr. Opin. Genet. Dev 2009, 19, 591–599. [Google Scholar]
- Garcia-Fernàndez, J; Benito-Gutiérrez, E. It’s a long way from amphioxus: descendants of the earliest chordate. Bioessays 2009, 31, 665–675. [Google Scholar]
- Hejnol, A; Obst, M; Stamatakis, A; Ott, M; Rouse, GW; Edgecombe, GD; Martinez, P; Baguñà, J; Bailly, X; Jondelius, U; Wiens, M; Müller, WE; Seaver, E; Wheeler, WC; Martindale, MQ; Giribet, G; Dunn, CW. Assessing the root of bilaterian animals with scalable phylogenomic methods. Proc. Biol. Sci 2009, 276, 4261–4270. [Google Scholar]
- Philippe, H; Derelle, R; Lopez, P; Pick, K; Borchiellini, C; Boury-Esnault, N; Vacelet, J; Renard, E; Houliston, E; Quéinnec, E; Da Silva, C; Wincker, P; Le Guyader, H; Leys, S; Jackson, DJ; Schreiber, F; Erpenbeck, D; Morgenstern, B; Wörheide, G; Manuel, M. Phylogenomics revives traditional views on deep animal relationships. Curr. Biol 2009, 19, 706–712. [Google Scholar]
- van de Peer, Y; Maere, S; Meyer, A. The evolutionary significance of ancient genome duplications. Nat. Rev. Genet 2009, 10, 725–732. [Google Scholar]
- Weil, D; Bernard, M; Gargano, S; Ramirez, F. The proα2(V) collagen gene is evolutionarily related to the major fibrillar-forming collagens. Nucleic Acids Res 1987, 15, 181–198. [Google Scholar]
- Bernard, M; Yoshioka, H; Rodriguez, E; van der Rest, M; Kimura, T; Ninomiya, Y; Olsen, BR; Ramirez, F. Cloning and sequencing of pro-α1(XI) collagen cDNA demonstrates that type XI belongs to the fibrillar class of collagens and reveals that the expression of the gene is not restricted to cartilagenous tissue. J. Biol. Chem 1988, 263, 17159–17166. [Google Scholar]
- Bulleid, NJ; Wilson, R; Lees, JF. Type-III procollagen assembly in semi-intact cells: chain association, nucleation and triple-helix folding do not require formation of inter-chain disulphide bonds but triple-helix nucleation does require hydroxylation. Biochem. J 1996, 317, 195–202. [Google Scholar]
- Zhang, G; Miyamoto, MM; Cohn, MJ. Lamprey type II collagen and Sox9 reveal an ancient origin of the vertebrate collagenous skeleton. Proc. Natl. Acad. Sci. USA 2006, 103, 3180–3185. [Google Scholar]
- Zhang, G; Cohn, MJ. Hagfish and lancelet fibrillar collagens reveal that type II collagen-based cartilage evolved in stem vertebrates. Proc. Natl. Acad. Sci. USA 2006, 103, 16829–16833. [Google Scholar]
- Ohtani, K; Yao, T; Kobayashi, M; Kusakabe, R; Kuratani, S; Wada, H. Expression of Sox and fibrillar collagen genes in lamprey larval chondrogenesis with implications for the evolution of vertebrate cartilage. J. Exp. Zool. B: Mol. Dev. Evol 2008, 310, 596–607. [Google Scholar]
- Wright, GM; Keeley, FW; Robson, P. The unusual cartilaginous tissues of jawless craniates, cephalochordates and invertebrates. Cell Tissue Res 2001, 304, 165–174. [Google Scholar]
- Kuraku, S; Meyer, A; Kuratani, S. Timing of genome duplications relative to the origin of the vertebrates: did cyclostomes diverge before or after? Mol. Biol. Evol 2009, 26, 47–59. [Google Scholar]
- Kuraku, S. Insights into cyclostome phylogenomics: Pre-2R or post-2R. Zoolog. Sci 2008, 25, 960–968. [Google Scholar]
- Rychel, AL; Swalla, BJ. Development and evolution of chordate cartilage. J. Exp. Zool. B: Mol. Dev. Evol 2007, 308, 325–335. [Google Scholar]
- Kadler, KE; Hill, A; Canty-Laird, EG. Collagen fibrillogenesis: fibronectin, integrins, and minor collagens as organizers and nucleators. Curr. Opin. Cell Biol 2008, 20, 495–501. [Google Scholar]
- Wenstrup, RJ; Florer, JB; Davidson, JM; Phillips, CL; Pfeiffer, BJ; Menezes, DW; Chervoneva, I; Birk, DE. Murine model of the Ehlers-Danlos syndrome col5a1 haploinsufficiency disrupts collagen fibril assembly at multiple stages. J. Biol. Chem 2006, 281, 12888–12895. [Google Scholar]
- Fernandes, RJ; Weis, M; Scott, MA; Seegmiller, RE; Eyre, DR. Collagen XI chain misassembly in cartilage of the chondrodysplasia (cho) mouse. Matrix Biol 2007, 26, 597–603. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Exposito, J.-Y.; Valcourt, U.; Cluzel, C.; Lethias, C. The Fibrillar Collagen Family. Int. J. Mol. Sci. 2010, 11, 407-426. https://doi.org/10.3390/ijms11020407
Exposito J-Y, Valcourt U, Cluzel C, Lethias C. The Fibrillar Collagen Family. International Journal of Molecular Sciences. 2010; 11(2):407-426. https://doi.org/10.3390/ijms11020407
Chicago/Turabian StyleExposito, Jean-Yves, Ulrich Valcourt, Caroline Cluzel, and Claire Lethias. 2010. "The Fibrillar Collagen Family" International Journal of Molecular Sciences 11, no. 2: 407-426. https://doi.org/10.3390/ijms11020407