Chalcones Enhance TRAIL-Induced Apoptosis in Prostate Cancer Cells


Abstract
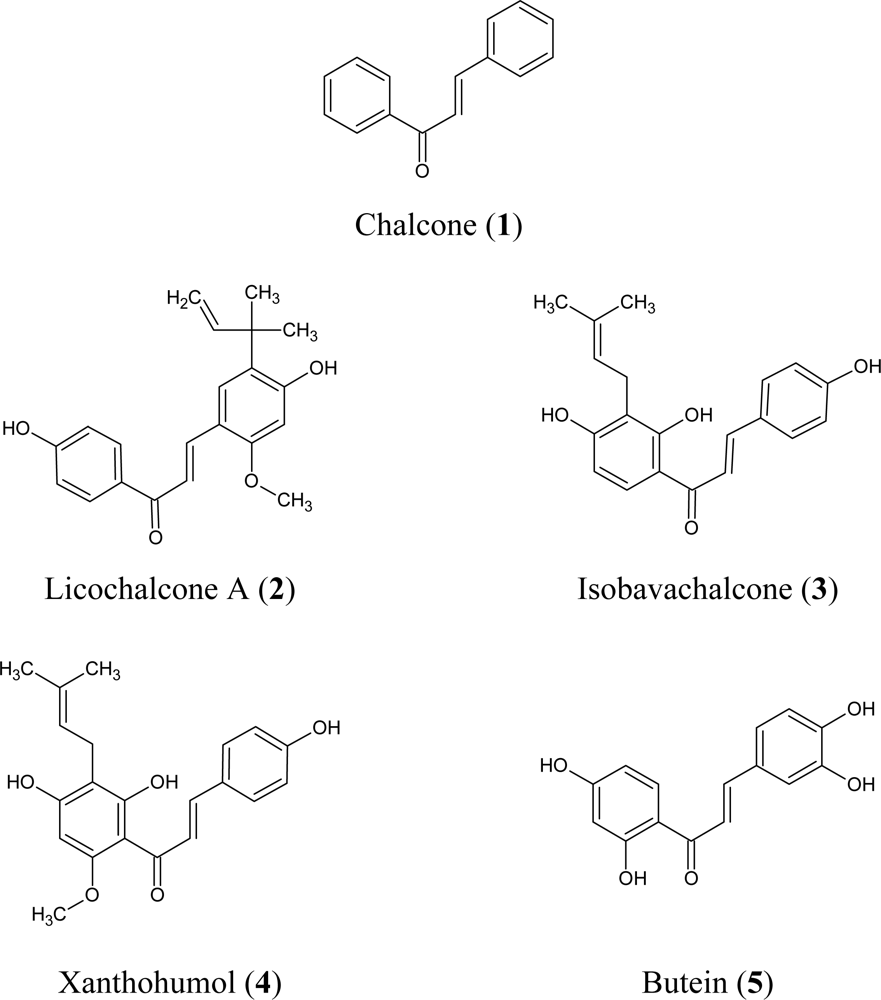
:1. Introduction
2. Results and Discussion
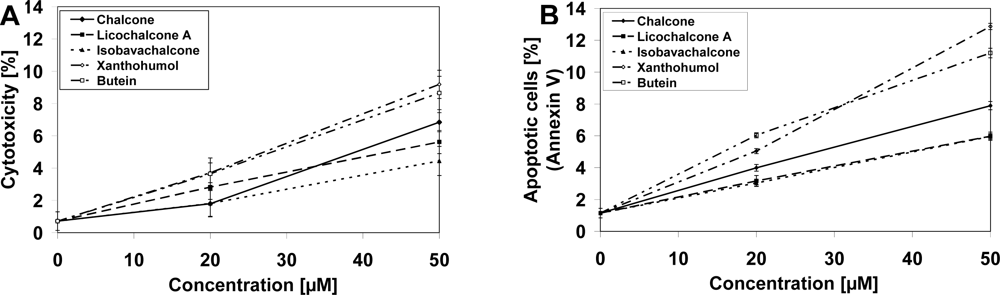
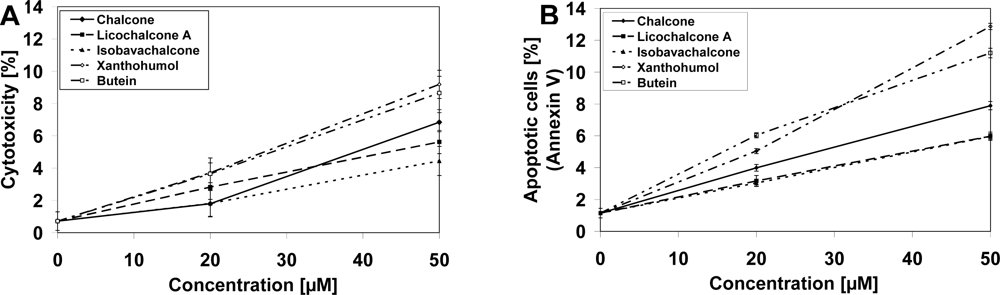
2.1. Cytotoxic and Apoptotic Effects of Chalcones in Prostate Cancer Cells
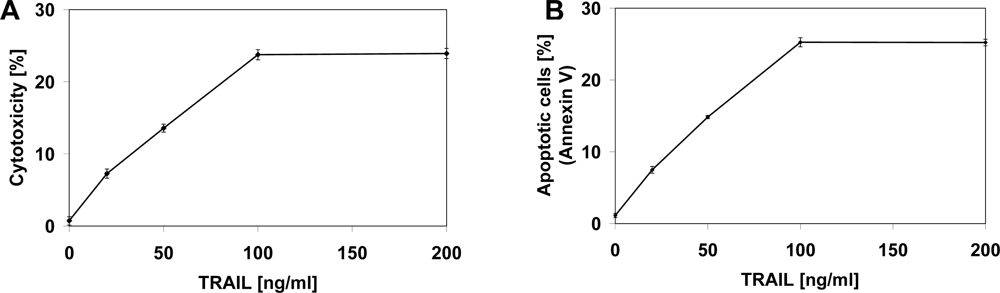
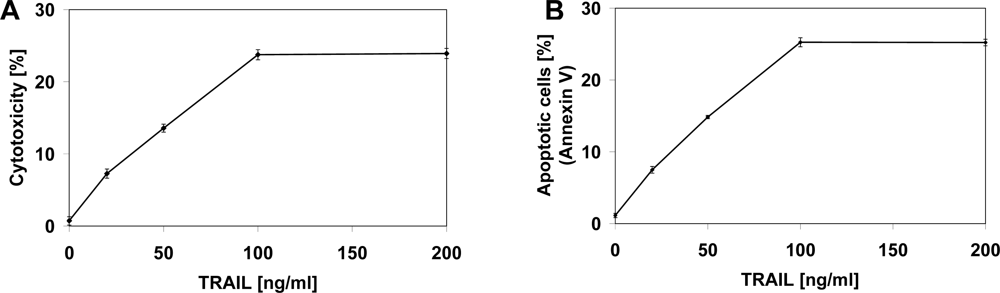
2.2. Cytotoxic and Apoptotic Effects of TRAIL in Prostate Cancer Cells
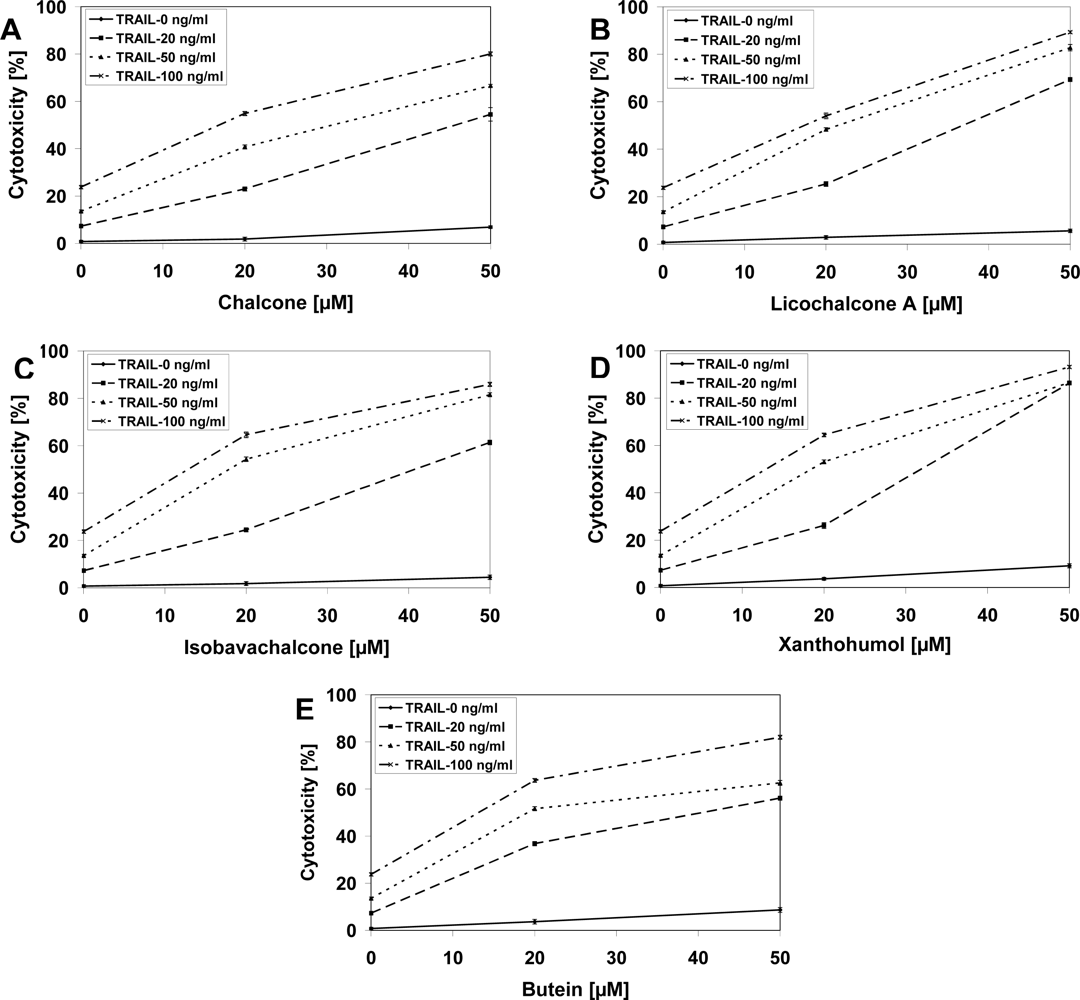
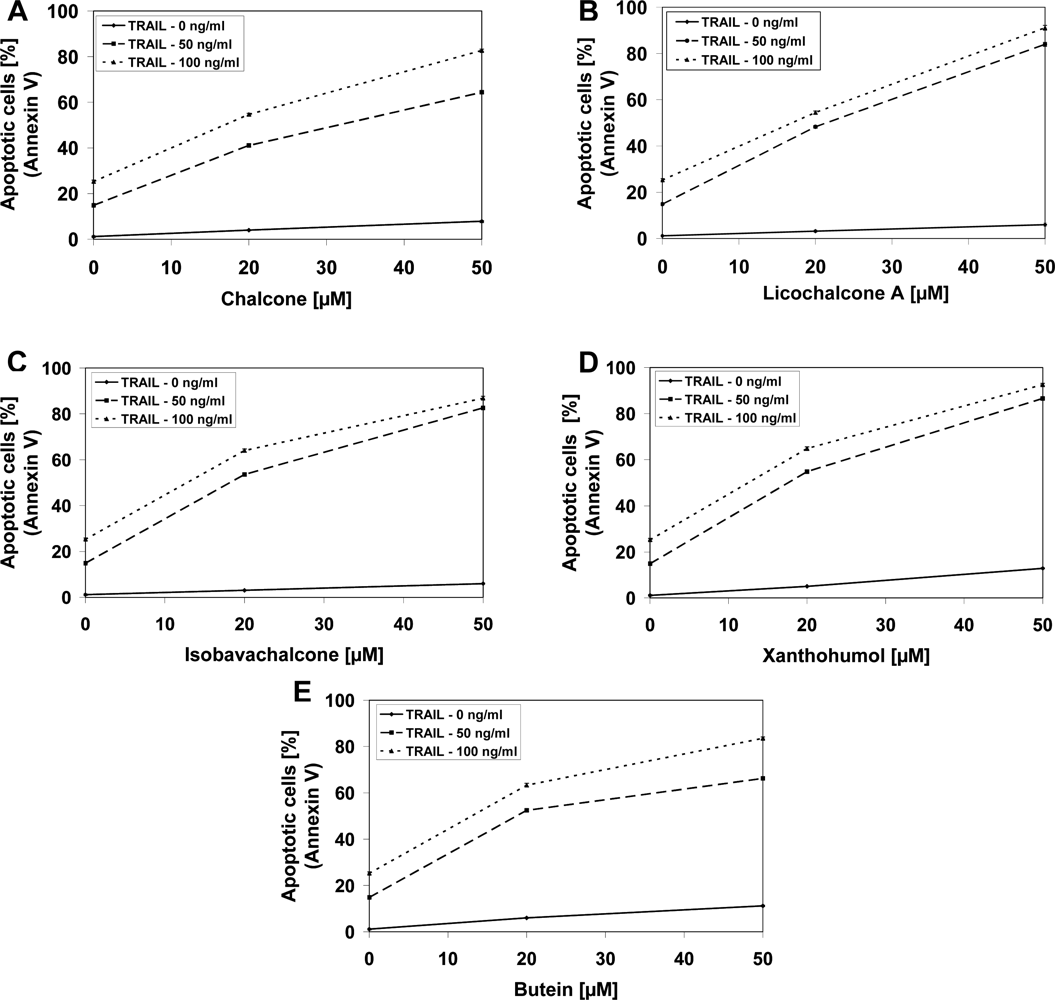
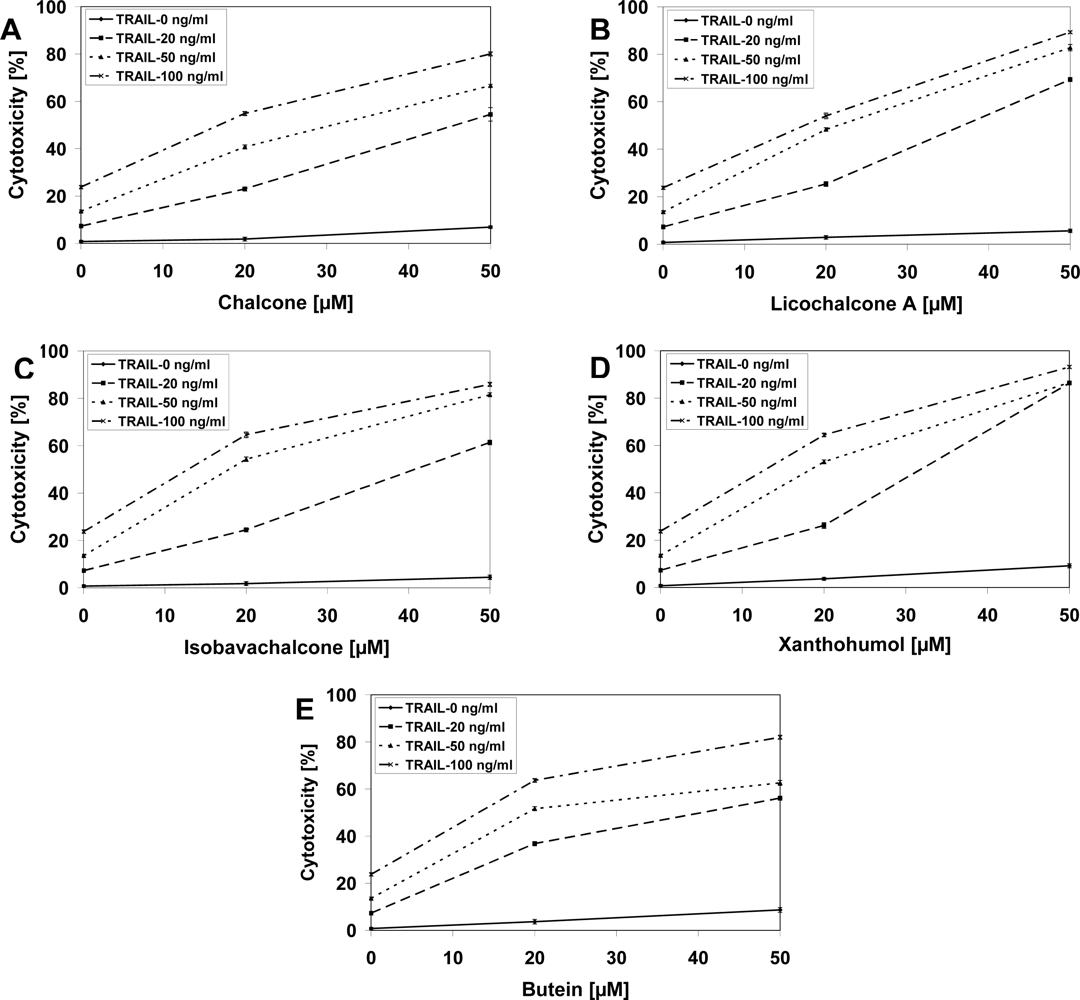
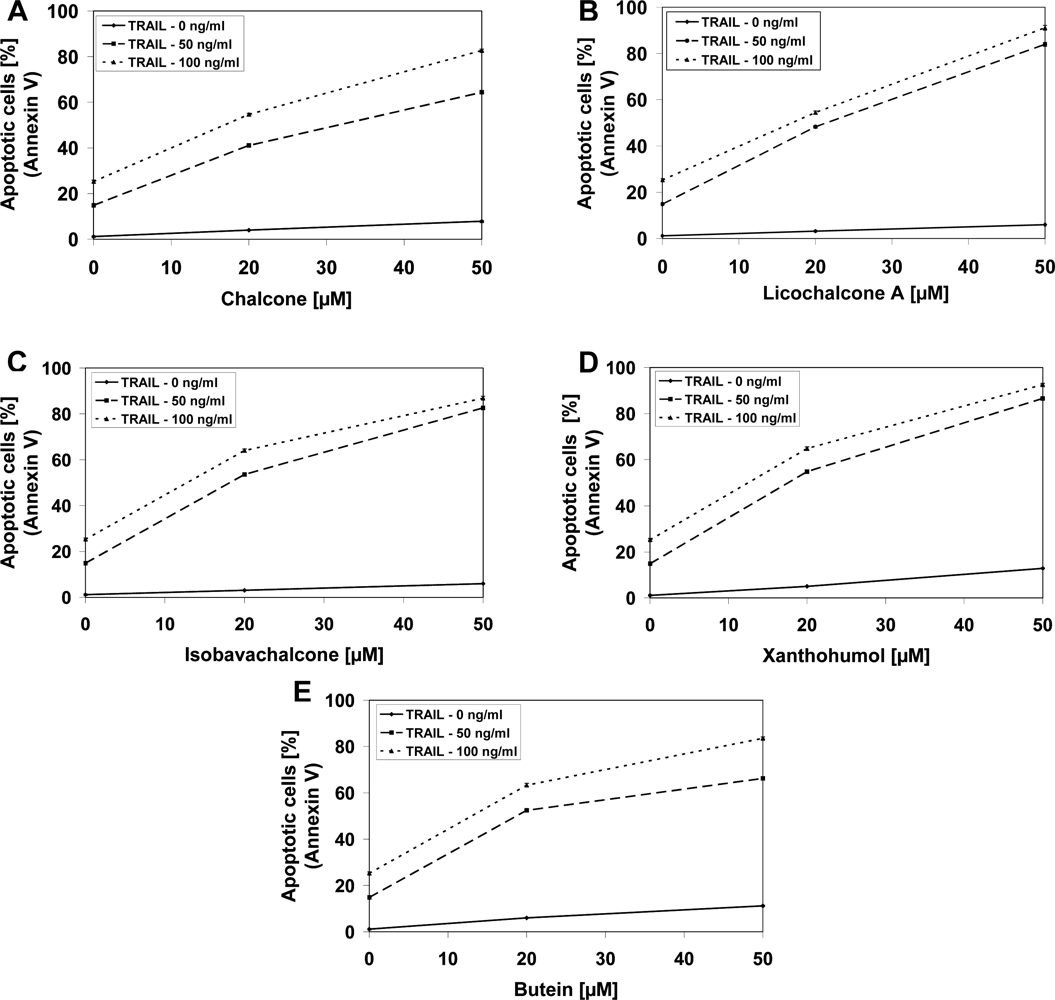
2.3. Cytotoxic and Apoptotic Effects of TRAIL in Combination with Chalcones in Prostate Cancer Cells
3. Experimental Section
3.1. Chemicals
3.1.1. Chalcones
3.1.2. TRAIL
3.2. Cell Culture
3.3. Cytotoxicity Assay
3.4. Lactate Dehydrogenase Release Assay
3.5. Determination of Apoptotic Cell Death by Annexin V-FITC Staining
3.6. Statistical Analysis
4. Conclusions
References
- Dicarlo, G; Mascolo, N; Izzo, AA; Capasso, F. Flavonoids: Old and new aspects of class of natural therapeutic drugs. Life Sci 1999, 65, 337–353. [Google Scholar]
- Dimmock, JR; Elias, DW; Beazely, MA; Kandepu, NM. Bioactivities of chalcones. Curr. Med. Chem 1999, 6, 1125–1149. [Google Scholar]
- Go, ML; Wu, X; Liu, XL. Chalcones: An update on cytotoxic and chemopreventive properties. Curr. Med. Chem 2005, 12, 481–499. [Google Scholar]
- Echeverria, C; Santibanez, JS; Donoso-Tauda, O; Escobar, CA; Ramirez-Tagle, R. Structural antitumoral activity relationships of synthetic chalcones. Int. J. Mol. Sci 2009, 10, 221–231. [Google Scholar]
- Nowakowska, Z. A review of anti-infective and anti-inflammatory chalcones. Eur. J. Med. Chem 2007, 42, 125–137. [Google Scholar]
- Miranda, CL; Stevens, JF; Helmrich, A; Henderson, MC; Rodriguez, RJ; Yang, YH; Deinzer, ML; Barnes, DW; Buhler, DR. Antiproliferative and cytotoxic effects of prenylated flavonoids from hops (Humulus lupus) in human cancer cell lines. Food Chem. Toxicol 1999, 37, 271–285. [Google Scholar]
- Shah, A; Khan, AM; Qureshi, R; Ansari, FL; Nazar, MF; Shah, SS. Redox behavior of anticancer chalcone on a glassy carbon electrode and evaluation of its interaction parameters with DNA. Int. J. Mol. Sci 2008, 9, 1424–1434. [Google Scholar]
- Boumendjel, A; Ronot, X; Boutonnat, J. Chalcones derivatives acting cell cycle blockers: Potential anticancer drugs? Curr. Drug Targets 2009, 10, 363–371. [Google Scholar]
- Katsori, AM; Hadjipavlou-Latina, D. Chalcones in cancer: Understanding their role in terms of QSAR. Curr. Med. Chem 2009, 16, 1062–1081. [Google Scholar]
- Heidenreich, A; Aus, G; Bolla, M; Joniau, S; Matveev, VB; Schmid, HP; Zattoni, F. European association of urology. EAU guidelines on prostate cancer. Eur. Urol 2008, 53, 68–80. [Google Scholar]
- Syed, DN; Suh, Y; Afag, F; Mukhtar, H. Dietary agents for chemoprevention of prostate cancer. Cancer Lett 2008, 265, 167–176. [Google Scholar]
- D’Archivio, M; Santangelo, C; Scazzocchio, B; Vari, R; Filesi, C; Masella, R; Giovannini, C. Modulatory effects of polyphenols on apoptosis induction: Relevance for cancer prevention. Int. J. Mol. Sci 2008, 9, 213–228. [Google Scholar]
- Zhou, J; Geng, G; Batist, G; Wu, JH. Syntheses and potential anti-prostate cancer activities of ionone-based chalcones. Bioorg. Med. Chem. Lett 2009, 19, 1183–1186. [Google Scholar]
- Yun, JM; Kweon, MH; Kwon, H; Hwang, JK; Mukhtar, H. Induction of apoptosis and cell cycle arrest by a chalcone panduratin A isolated from Kaempferia pandurata in androgen-independent prostate cancer cells PC3 and DU145. Carcinogenesis 2006, 27, 1454–1564. [Google Scholar]
- Fu, Y; Hsieh, TC; Guo, J; Kunicki, J; Lee, MY; Darzynkiewicz, Z; Wu, JM. Licochalcone-A, a novel flavonoid isolated from licorice root (Glycyrrhiza glabra) causes G2 and late-G1 arrests in androgen-independent PC3 prostate cancer cells. Biochem. Biophys. Res. Commun 2004, 322, 263–270. [Google Scholar]
- Lee, YM; Lim, Y; Choi, HJ; Jung, JI; Chung, WY; Park, JH. Induction of cell cycle arrest in prostate cancer cells by the dietary compound isoliquiritigenin. J. Med. Food 2009, 12, 8–14. [Google Scholar]
- Desmulle, L; Bellahcene, A; Dhooge, W; Comhaire, F; Roelens, F; Huvaere, K; Heyerick, A; Castronovo, V; Dekeukeleire, D. Antiproliferative properties of prenylated flavonoids from hops (Humulus lupulus L.) in prostate cancer cell lines. Phytomedicine 2006, 13, 732–734. [Google Scholar]
- Almasan, A; Ashkenazi, A. Apo2L/TRAIL: Apoptosis signaling, biology and potential for cancer therapy. Cytokine Growth Factor Rev 2003, 14, 337–348. [Google Scholar]
- Thorburn, A; Behbakht, K; Ford, H. TRAIL receptor-targeted therapeutics: Resistance mechanisms and strategies to avoid them. Drug Resist. Updat 2008, 11, 17–24. [Google Scholar]
- Zhang, L; Fang, B. Mechanisms of resistance to TRAIL-induced apoptosis in cancer. Cancer Gene Ther 2005, 12, 228–237. [Google Scholar]
- Shankar, S; Singh, TR; Srivastava, RK. Ionizing radiation enhances the therapeutic potential of TRAIL in prostate cancer in vitro and in vivo: Intracellular mechanisms. The Prostate 2004, 61, 35–49. [Google Scholar]
- Horinaka, M; Yoshida, T; Shiraishi, T; Nakata, S; Wakada, M; Sakai, T. The dietary flavonoid apigenin sensitizes malignant tumor cells to tumor necrosis factor-related apoptosis-inducing ligand. Mol. Cancer Ther 2006, 5, 945–951. [Google Scholar]
- Shankar, S; Chen, Q; Siddiqui, I; Sarva, K; Srivastava, RK. Sensitization of TRAIL-resistant LNCaP cells by resveratrol (3,4′,5-trihydroxystilbene): Molecular mechanisms and therapeutic potential. J. Mol. Signal 2007, 2, 27–36. [Google Scholar]
- Szliszka, E; Bronikowska, J; Majcher, A; Miszkiewicz, J; Krol, W. Enhanced sensitivity of hormone-refractory prostate cancer cells to tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) mediated cytotoxicity by taxanes. CEJ Urol 2009, 62, 29–34. [Google Scholar]
- Szliszka, E; Czuba, ZP; Bronikowska, J; Mertas, A; Paradysz, A; Krol, W. Ethanolic extract of propolis (EEP) augments TRAIL-induced apoptotic death in prostate cancer cells. Evid Based Complement Alternat Med 2009. [Google Scholar]
- Thomasset, SC; Berry, TP; Garcea, G; Marczylo, T; Steward, WP; Gescher, AJ. Dietary polyphenolic phytochemicals—Promising cancer chemopreventive agents in humans? A review of their clinical properties. Int. J. Cancer 2007, 120, 451–458. [Google Scholar]
- Khan, N; Adhami, VM; Mukhtar, H. Apoptosis by dietary agents for prevention and treatment of cancer. Biochem. Pharmacol 2008, 76, 1333–1339. [Google Scholar]
- Szliszka, E; Czuba, ZP; Jernas, K; Krol, W. Dietary flavonoids sensitize HeLa cells to tumor necrosis factor-related apoptosis-inducing ligand (TRAIL). Int. J. Mol. Sci 2008, 9, 56–64. [Google Scholar]
- Han, X; Shen, T; Lou, H. Dietary polyphenols and their biological significance. Int. J. Mol. Sci 2007, 8, 950–988. [Google Scholar]
- Ozmen, A; Madlener, S; Buer, S; Krasteva, S; Vonach, C; Giessrigl, B; Gridling, M; Viola, K; Stark, N; Saiko, P; Michel, B; Fritzer-Szekeres, M; Szekeres, T; Askin-Celik, T; Krenn, L; Krupitza, G. In vitro anti-leukemic activity of the ethno-pharmacological plant Scutellaria orientalis ssp carica endemic to western Turkey. Phytomedicine 2009. [Google Scholar]
- Ferguson, LR; Philpott, M. Cancer prevention by dietary bioactive components that target the immune response. Curr. Cancer Drug Targets 2007, 7, 459–464. [Google Scholar]
- Jakóbisiak, M; Lasek, W; Gołąb, J. Natural mechanisms protecting against cancer. Immunol. Lett 2003, 90, 103–122. [Google Scholar]
- Yoshida, T; Horinaka, M; Takara, M; Tsuchihashi, M; Mukai, N; Wakada, M; Sakai, T. Combination of isoliquiritigenin and tumor necrosis factor-related apoptosis-inducing ligand induces apoptosis in colon cancer HT29 cells. Environ. Health Prev. Med 2008, 13, 281–287. [Google Scholar]
- Kim, N. Butein sensitizes human leukemia cells to apoptosis induced by tumor necrosis factor-related apoptosis inducing ligand (TRAIL). Arch. Pharm. Res 2008, 31, 1179–1186. [Google Scholar]
- Szliszka, E; Czuba, ZP; Domino, M; Mazur, B; Zydowicz, G; Krol, W. Ethanolic extract of propolis (EEP) enhances the apoptosis-inducing potential of TRAIL in cancer cells. Molecules 2009, 14, 738–754. [Google Scholar]
- Moreira, L; Dias, LG; Pereira, JA; Estevinho, L. Antioxidant properties, total phenols and pollen analysis of propolis sample from Portugal. Food Chem. Toxicol 2008, 46, 3482–3485. [Google Scholar]
- Kim, KT; Yoo, KM; Lee, JW; Eom, SH; Hwang, IK; Lee, CY. Protective effect of steamed American ginseng (Panax quinquefolius L.) on V79-4 cells induced by oxidative stress. J. Ethnopharmacol 2007, 111, 443–450. [Google Scholar]
- Krol, W; Czuba, Z; Scheller, S; Gabrys, S; Shani, J. Antioxidant property of ethanolic extract of propolis (EEP) as evaluated by inhibiting the chemiluminescence oxidation of luminol. Biochem. Inter 1990, 21, 593–597. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Post-hoc test | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| p= | (1) | (2) | (3) | (4) | (5) | (6) | (7) | (8) | (9) | (10) | (11) |
| Control: LNCaP cells (1) | 2E-05 | 1E-05 | 8E-06 | 1E-05 | 2E-05 | 3E-05 | 3E-05 | 2E-05 | 1E-05 | 2E-05 | |
| Chalcone 20 [μM] (2) | 2E-05 | 2E-05 | 2E-05 | 8E-06 | 8E-06 | 2E-05 | 9E-06 | 3E-05 | 2E-05 | 3E-05 | |
| Chalcone 50 [μM] (3) | 1E-05 | 2E-05 | 3E-05 | 2E-05 | 1E-05 | 8E-06 | 2E-05 | 2E-05 | 9E-06 | 9E-06 | |
| Licochalcone-A 20 [μM] (4) | 8E-06 | 2E-05 | 3E-05 | 2E-05 | 0.1632 | 2E-05 | 8E-06 | 1E-05 | 3E-05 | 1E-05 | |
| Licochalcone-A 50 [μM] (5) | 1E-05 | 8E-06 | 2E-05 | 2E-05 | 3E-05 | 0.8617 | 2E-05 | 2E-05 | 0.5419 | 8E-06 | |
| Isobavachalcone 20 [μM] (6) | 2E-05 | 8E-06 | 1E-05 | 0.1632 | 3E-05 | 2E-05 | 2E-05 | 1E-05 | 3E-05 | 1E-05 | |
| Isobavachalcone 50 [μM] (7) | 3E-05 | 2E-05 | 8E-06 | 2E-05 | 0.8617 | 2E-05 | 9E-06 | 2E-05 | 0.7127 | 2E-05 | |
| Xanthohumol 20 [μM] (8) | 3E-05 | 9E-06 | 2E-05 | 8E-06 | 2E-05 | 2E-05 | 9E-06 | 3E-05 | 8E-06 | 2E-05 | |
| Xanthohumol 50 [μM] (9) | 2E-05 | 3E-05 | 2E-05 | 1E-05 | 2E-05 | 1E-05 | 2E-05 | 3E-05 | 8E-06 | 9E-06 | |
| Butein 20 [μM] (10) | 1E-05 | 2E-05 | 9E-06 | 3E-05 | 0.5419 | 3E-05 | 0.7127 | 8E-06 | 8E-06 | 2E-05 | |
| Butein 50 [μM] (11) | 2E-05 | 3E-05 | 9E-06 | 1E-05 | 8E-06 | 1E-05 | 2E-05 | 2E-05 | 9E-06 | 2E-05 | |
| Post - hoc test. | TRAIL - 50 ng/ml | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Chalcones 20 μM | Chalcones 50 μM | |||||||||
| p= | (1) | (2) | (3) | (4) | (5) | (1) | (2) | (3) | (4) | (5) |
| Chalcone (1) | 0.000112 | 0.000161 | 0.000129 | 0.00012 | 0.000161 | 0.00012 | 0.000129 | 0.000112 | ||
| Licochalcone (2) | 0.000112 | 0.00012 | 0.000161 | 0.000112 | 0.000161 | 0.000112 | 0.000112 | 0.00012 | ||
| Isobavachalcone (3) | 0.000161 | 0.00012 | 0.000126 | 0.000257 | 0.00012 | 0.000112 | 0.00012 | 0.000112 | ||
| Xanthohumol (4) | 0.000129 | 0.000161 | 0.000126 | 0.00012 | 0.000129 | 0.000112 | 0.00012 | 0.000161 | ||
| Butein (5) | 0.00012 | 0.000112 | 0.000257 | 0.00012 | 0.000112 | 0.00012 | 0.000112 | 0.000161 | ||
| TRAIL - 100 ng/ml | ||||||||||
| Chalcones 20 μM | Chalcones 50 μM | |||||||||
| p= | (1) | (2) | (3) | (4) | (5) | (1) | (2) | (3) | (4) | (5) |
| Chalcone (1) | 0.402463 | 0.00012 | 0.000161 | 0.000112 | 0.000161 | 0.00012 | 0.000129 | 0.010564 | ||
| Licochalcone (2) | 0.402463 | 0.000161 | 0.000129 | 0.00012 | 0.000161 | 0.000112 | 0.000138 | 0.00012 | ||
| Isobavachalcone (3) | 0.00012 | 0.000161 | 0.00315 | 0.019365 | 0.00012 | 0.000112 | 0.00012 | 0.000112 | ||
| Xanthohumol (4) | 0.000161 | 0.000129 | 0.00315 | 0.000122 | 0.000129 | 0.000138 | 0.00012 | 0.000161 | ||
| Butein (5) | 0.000112 | 0.00012 | 0.019365 | 0.000122 | 0.010564 | 0.00012 | 0.000112 | 0.000161 | ||
© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Szliszka, E.; Czuba, Z.P.; Mazur, B.; Sedek, L.; Paradysz, A.; Krol, W. Chalcones Enhance TRAIL-Induced Apoptosis in Prostate Cancer Cells. Int. J. Mol. Sci. 2010, 11, 1-13. https://doi.org/10.3390/ijms11010001
Szliszka E, Czuba ZP, Mazur B, Sedek L, Paradysz A, Krol W. Chalcones Enhance TRAIL-Induced Apoptosis in Prostate Cancer Cells. International Journal of Molecular Sciences. 2010; 11(1):1-13. https://doi.org/10.3390/ijms11010001
Chicago/Turabian StyleSzliszka, Ewelina, Zenon P. Czuba, Bogdan Mazur, Lukasz Sedek, Andrzej Paradysz, and Wojciech Krol. 2010. "Chalcones Enhance TRAIL-Induced Apoptosis in Prostate Cancer Cells" International Journal of Molecular Sciences 11, no. 1: 1-13. https://doi.org/10.3390/ijms11010001