Inhibition of Citrinin-Induced Apoptotic Biochemical Signaling in Human Hepatoma G2 Cells by Resveratrol

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
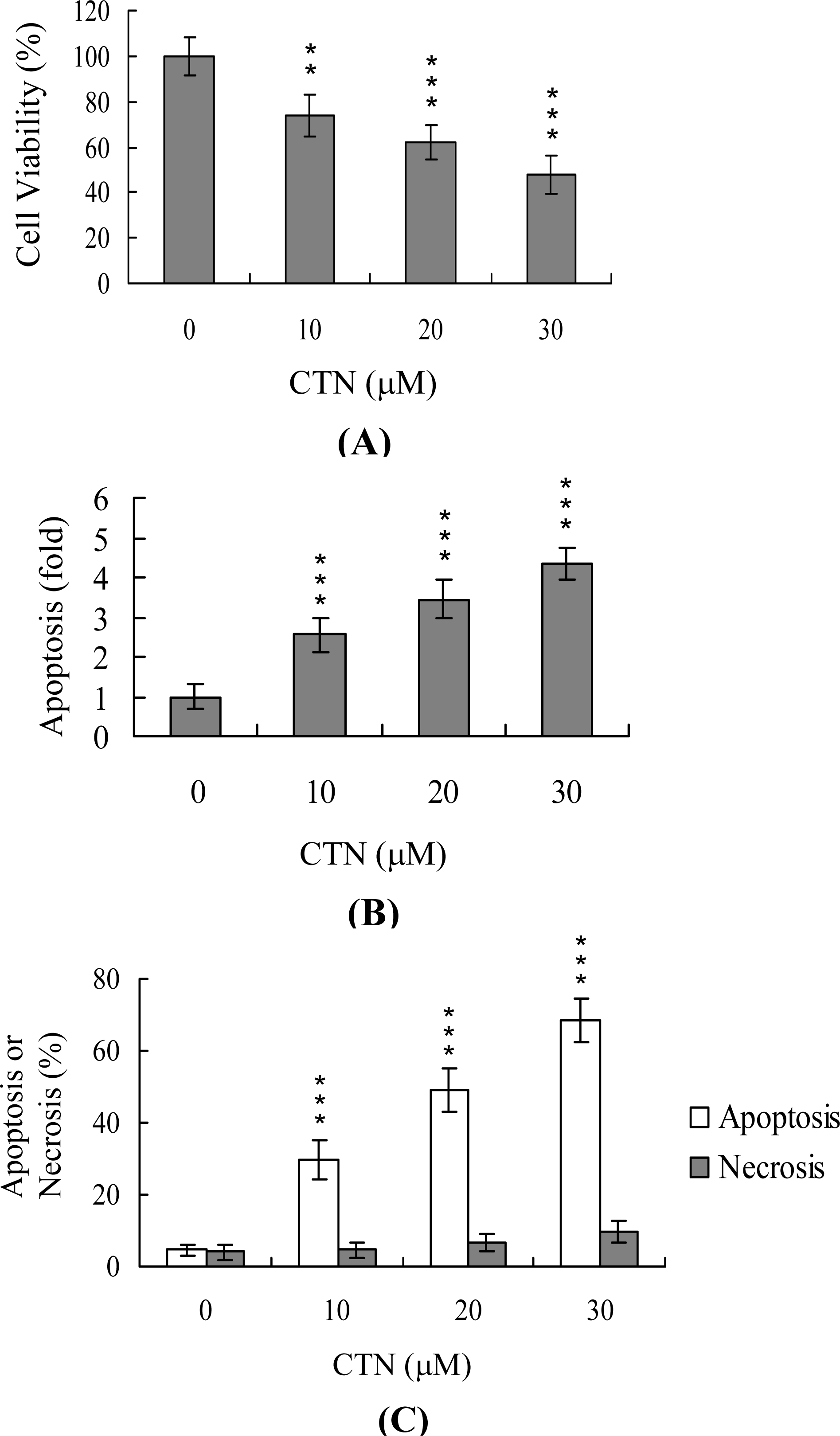
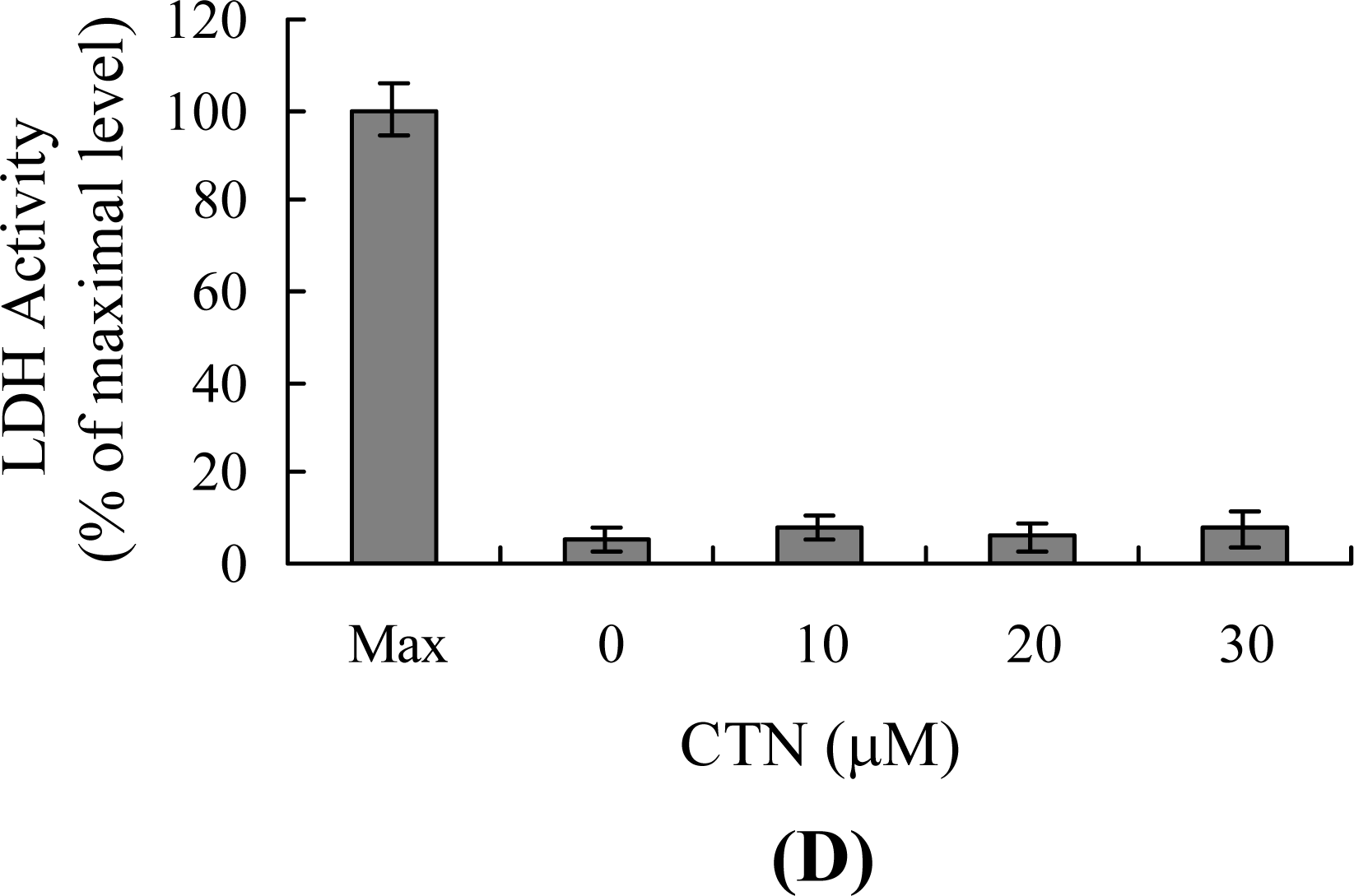
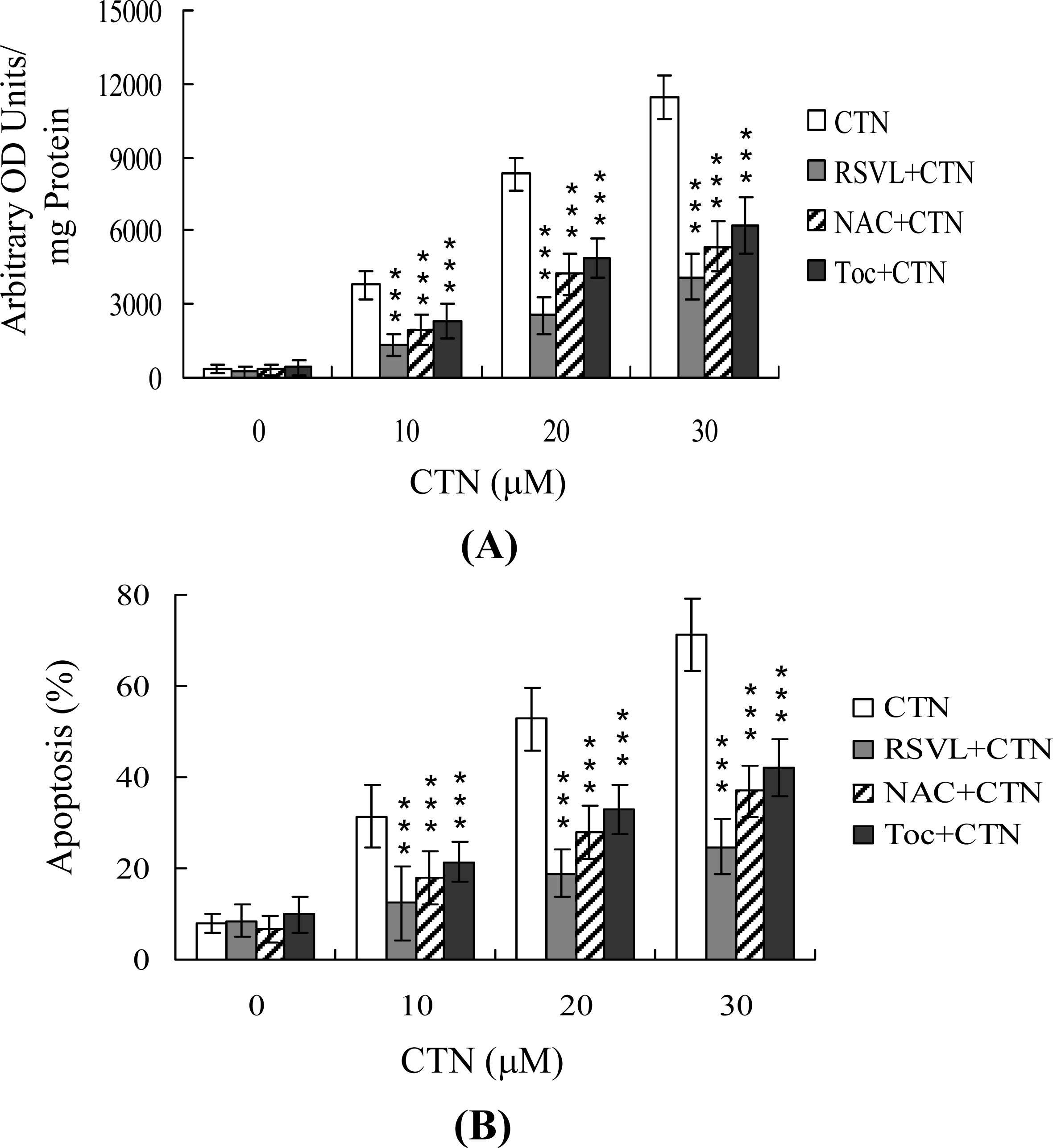
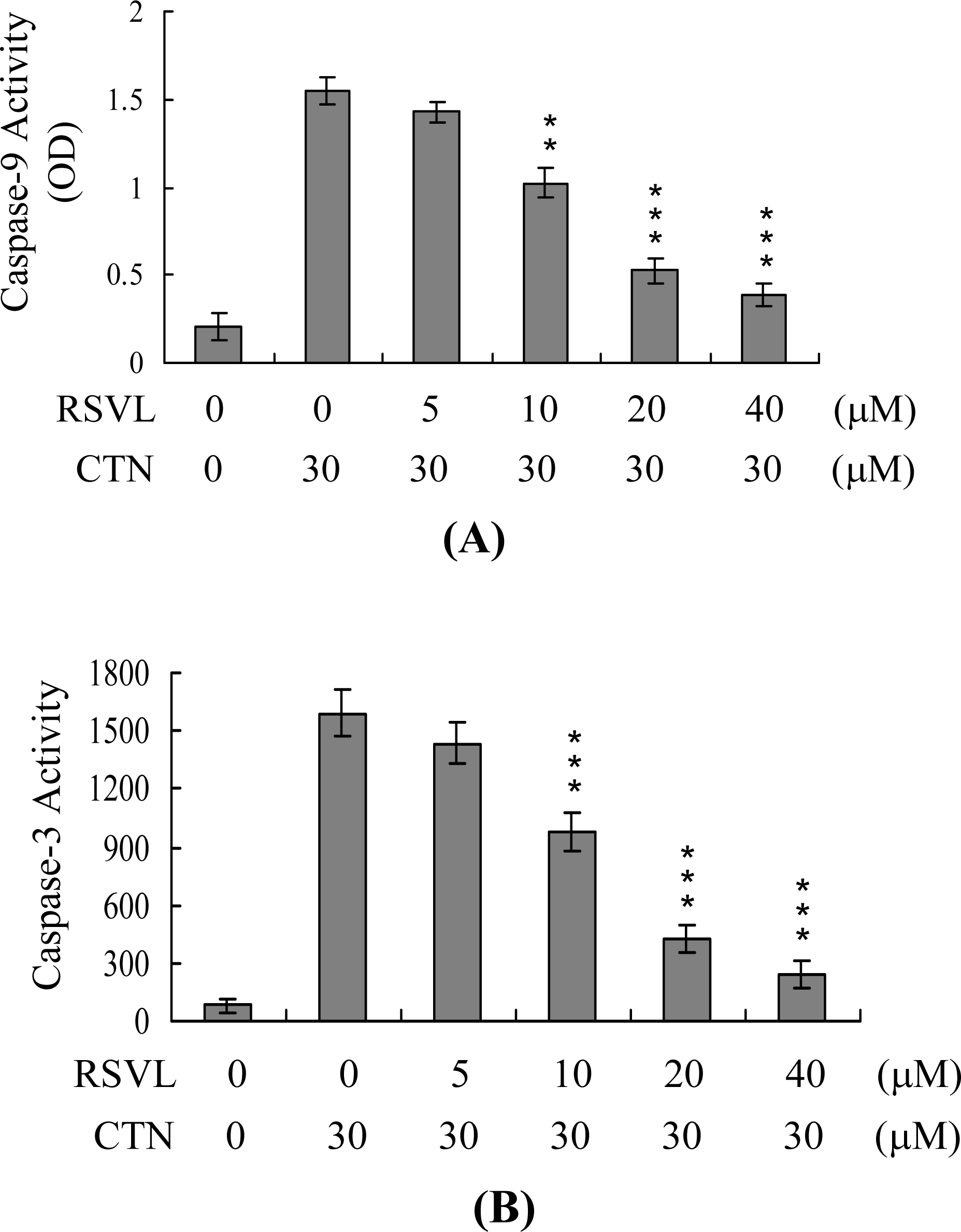
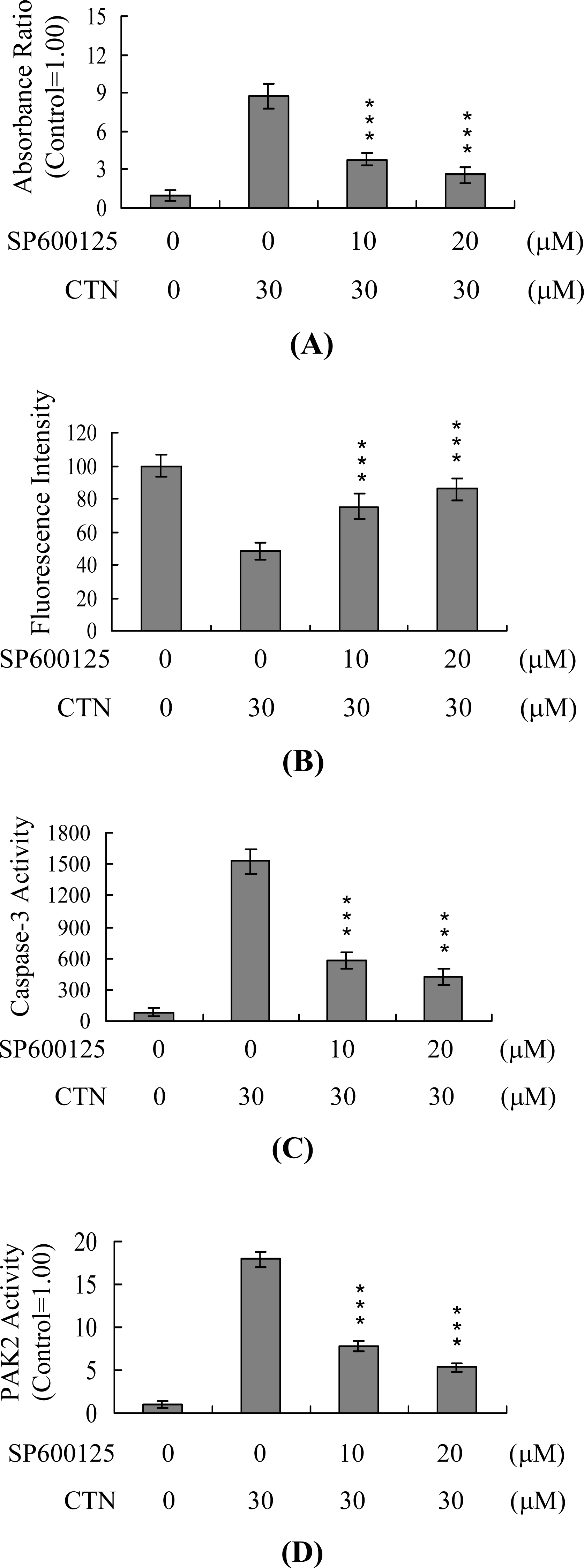
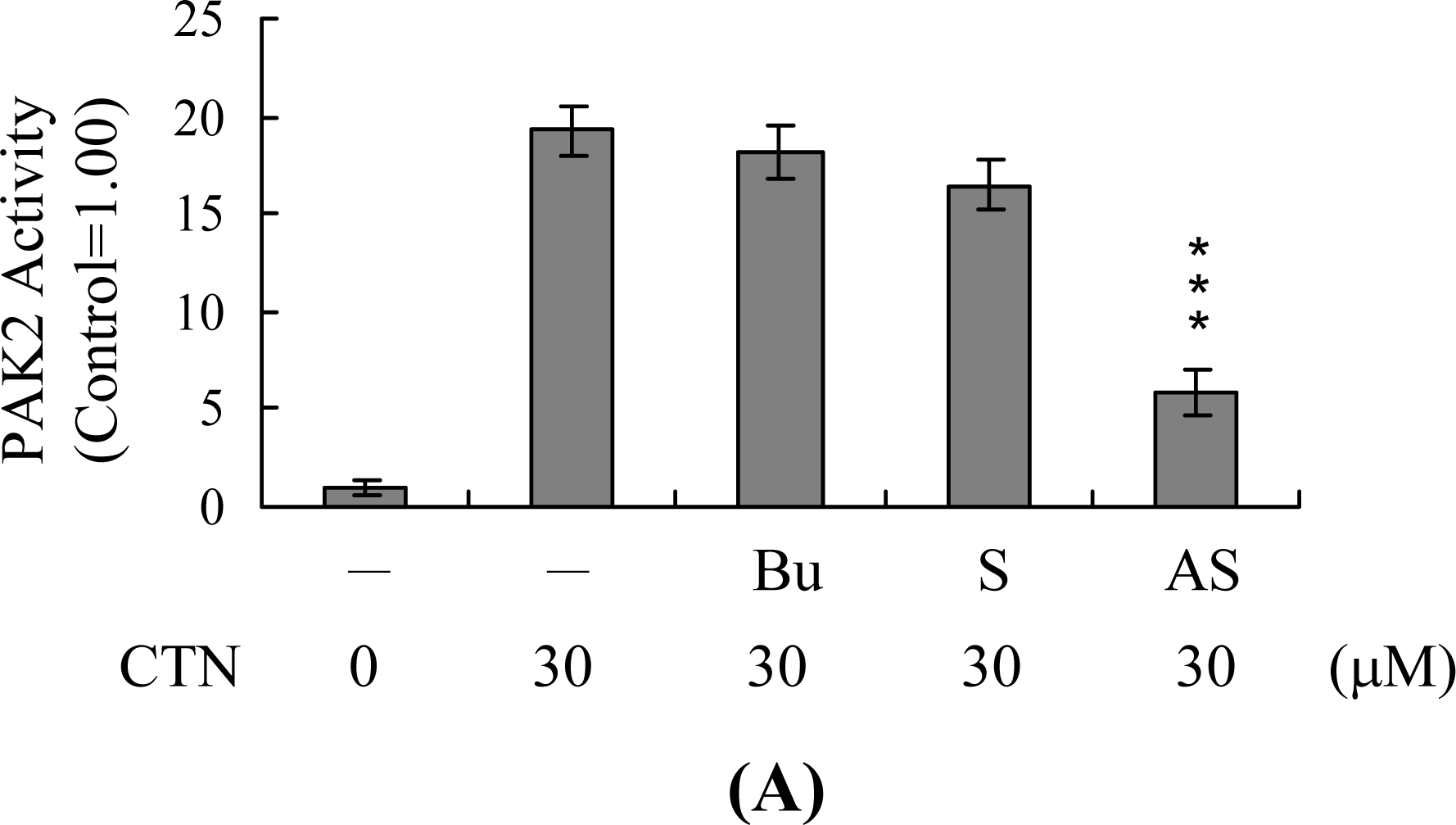
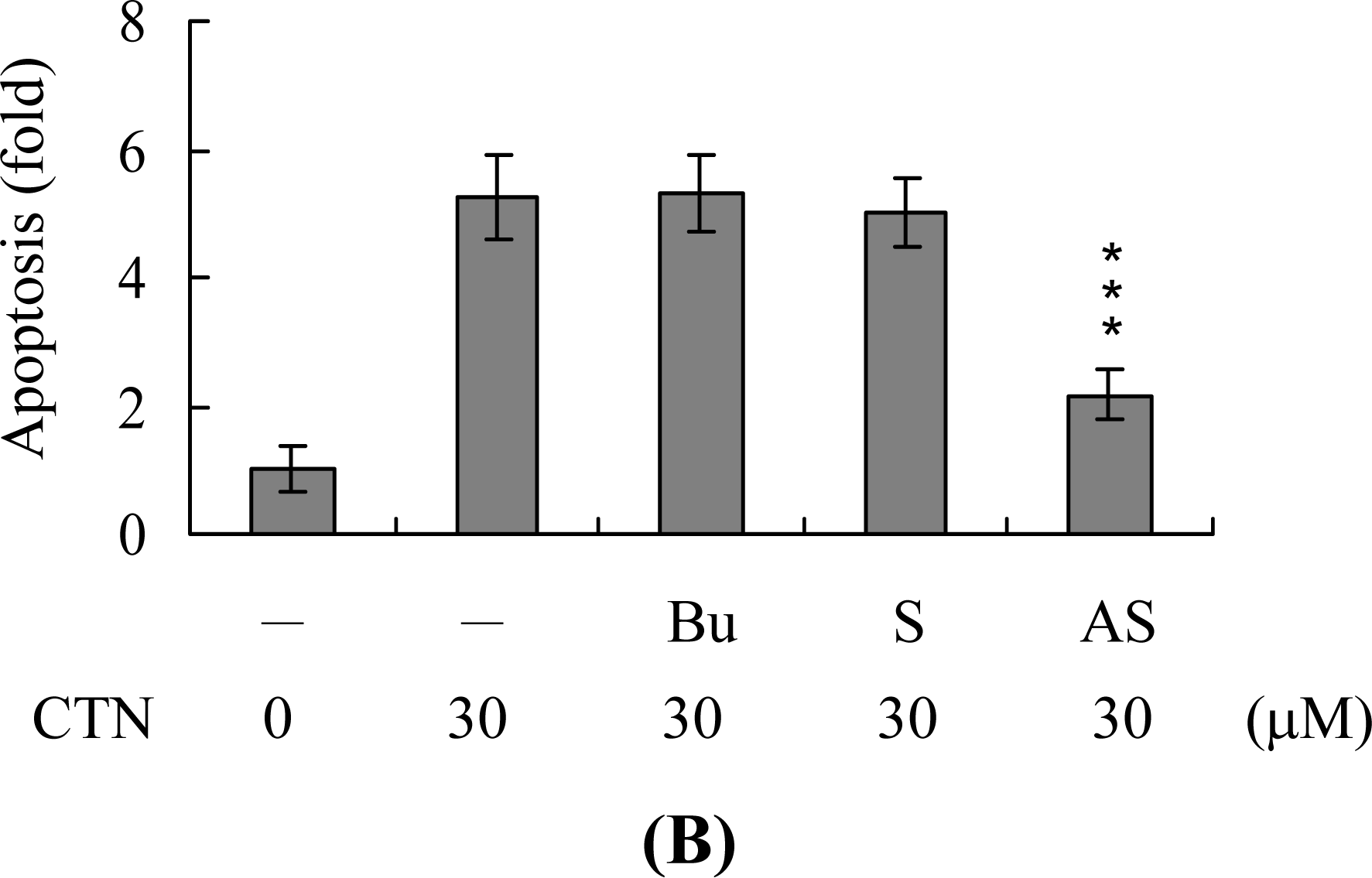
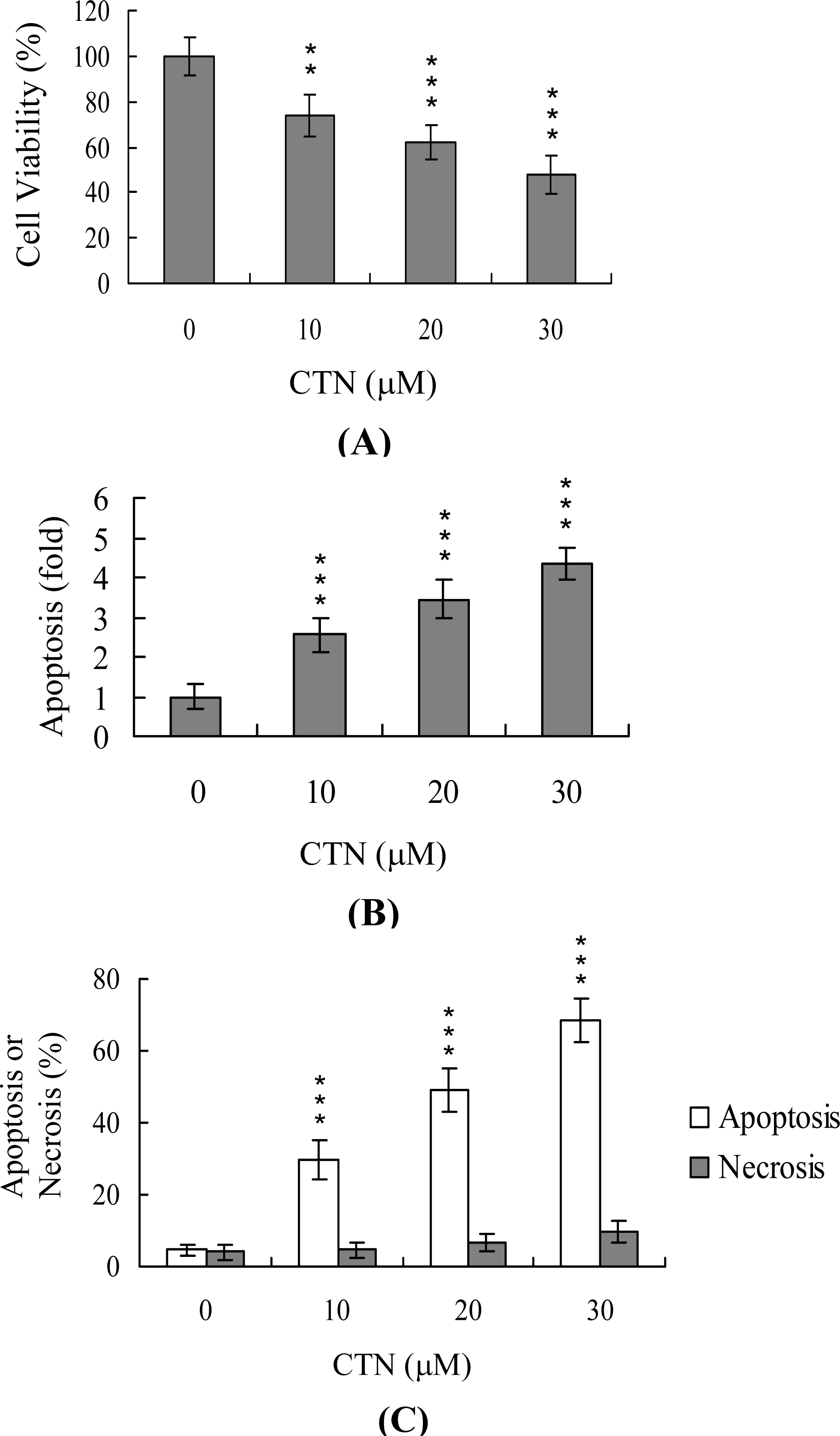
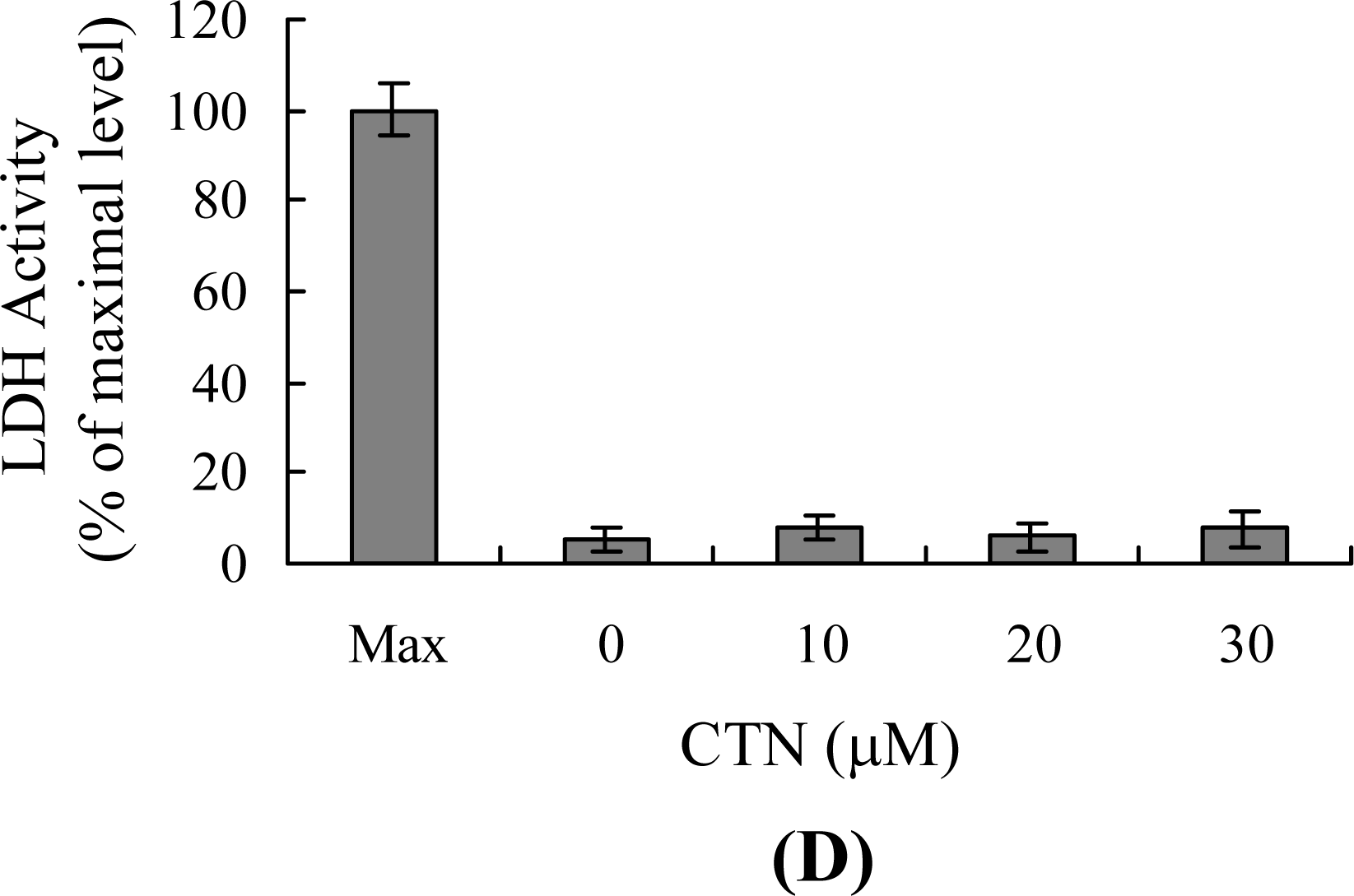
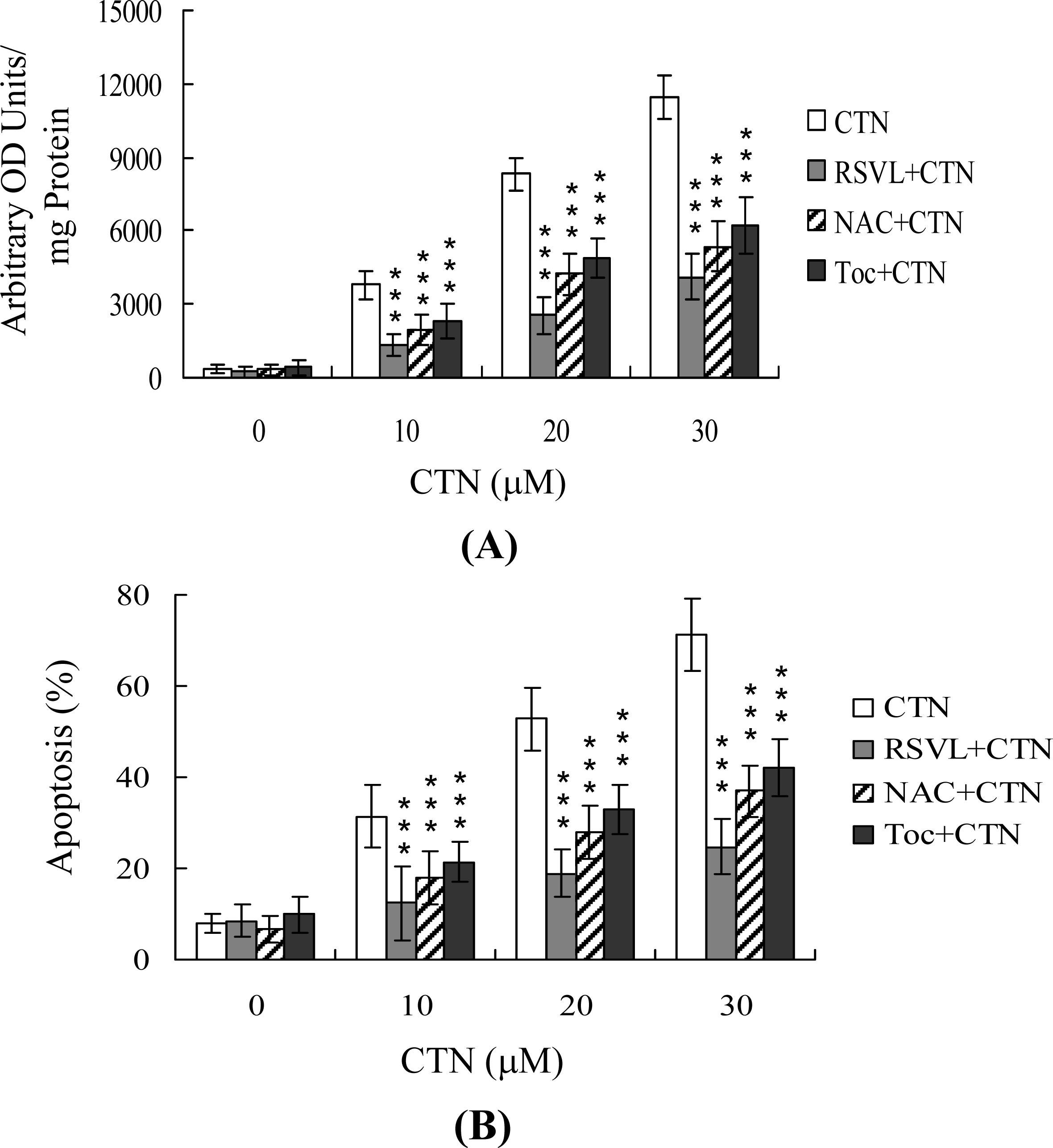
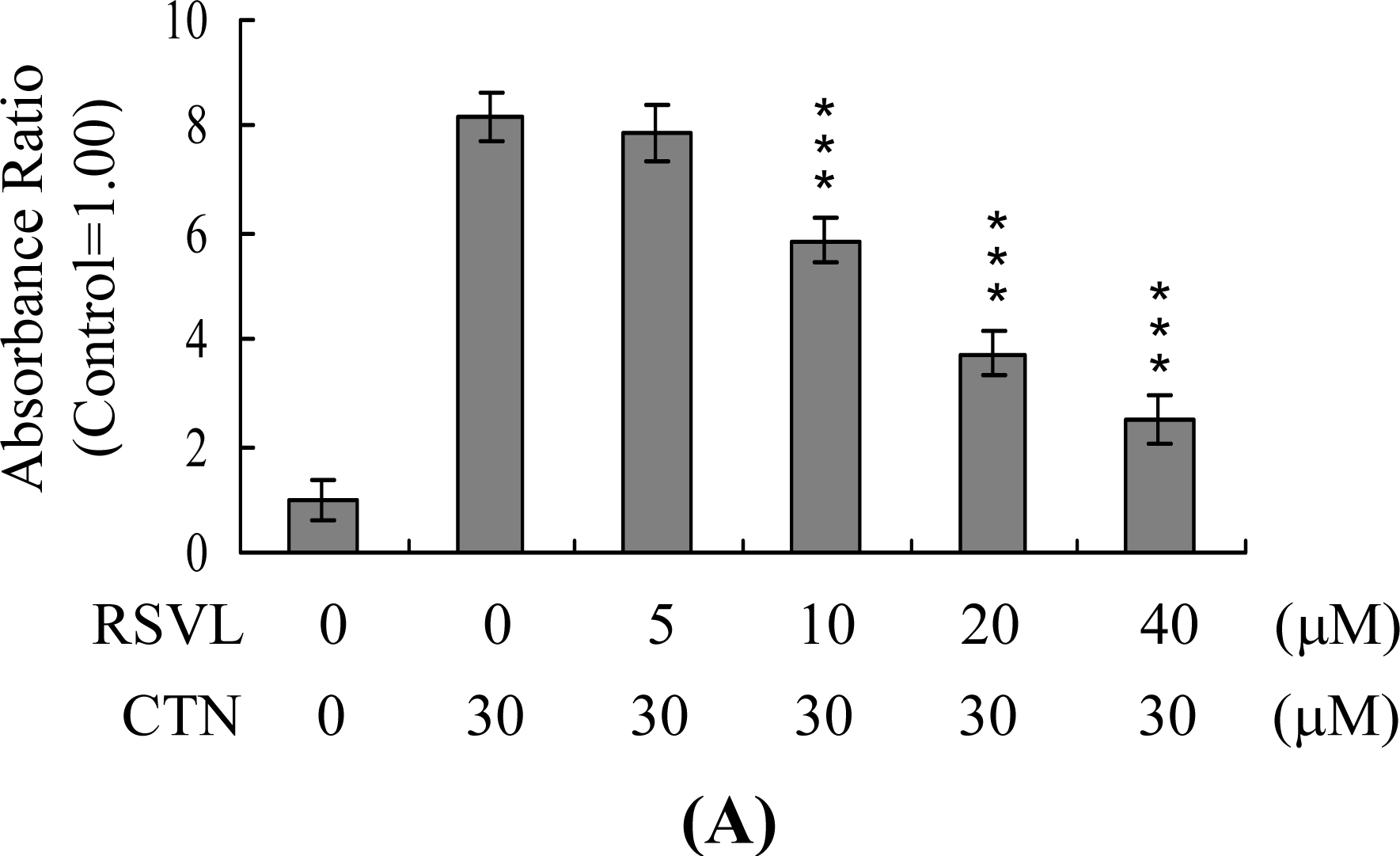
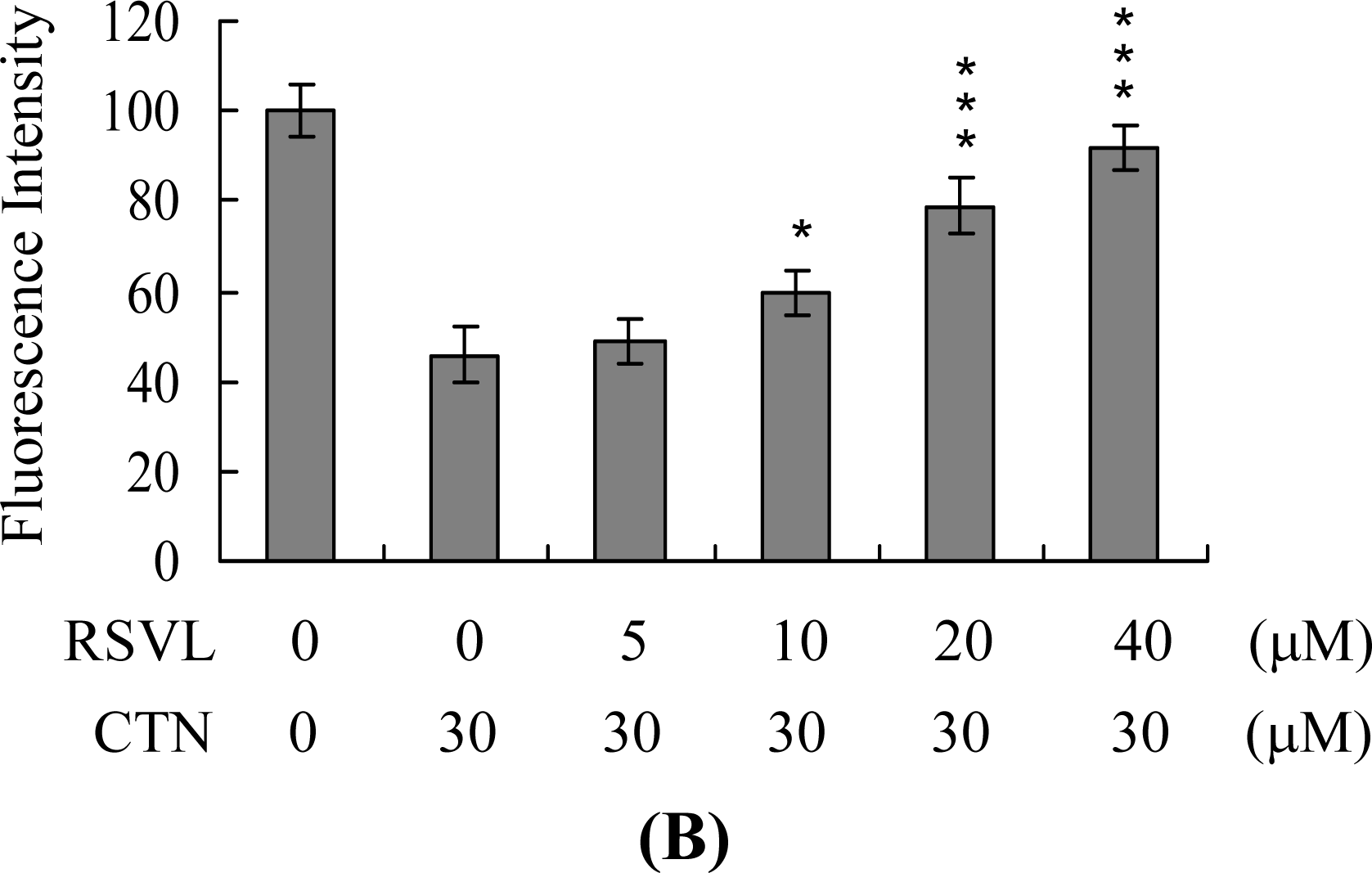
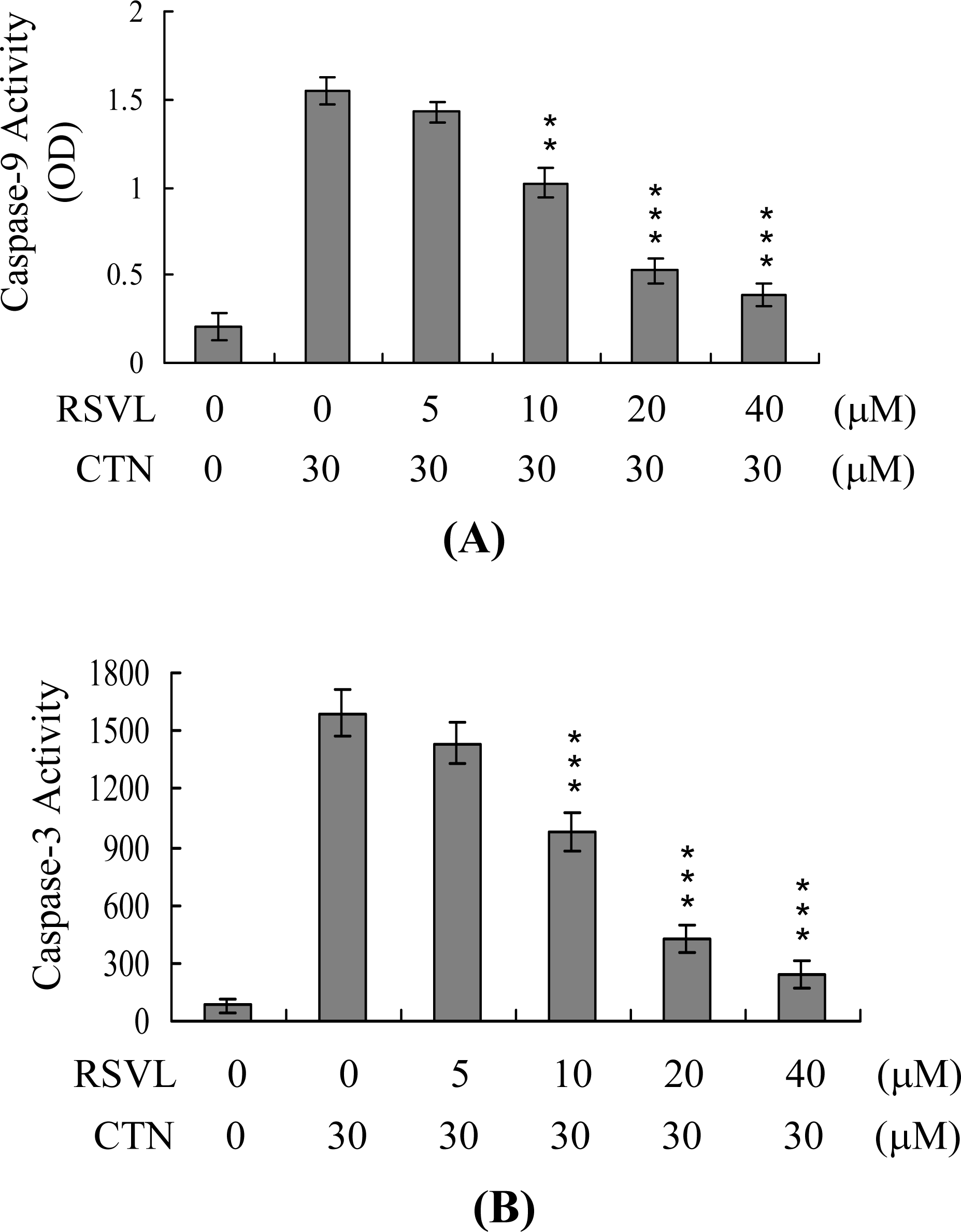
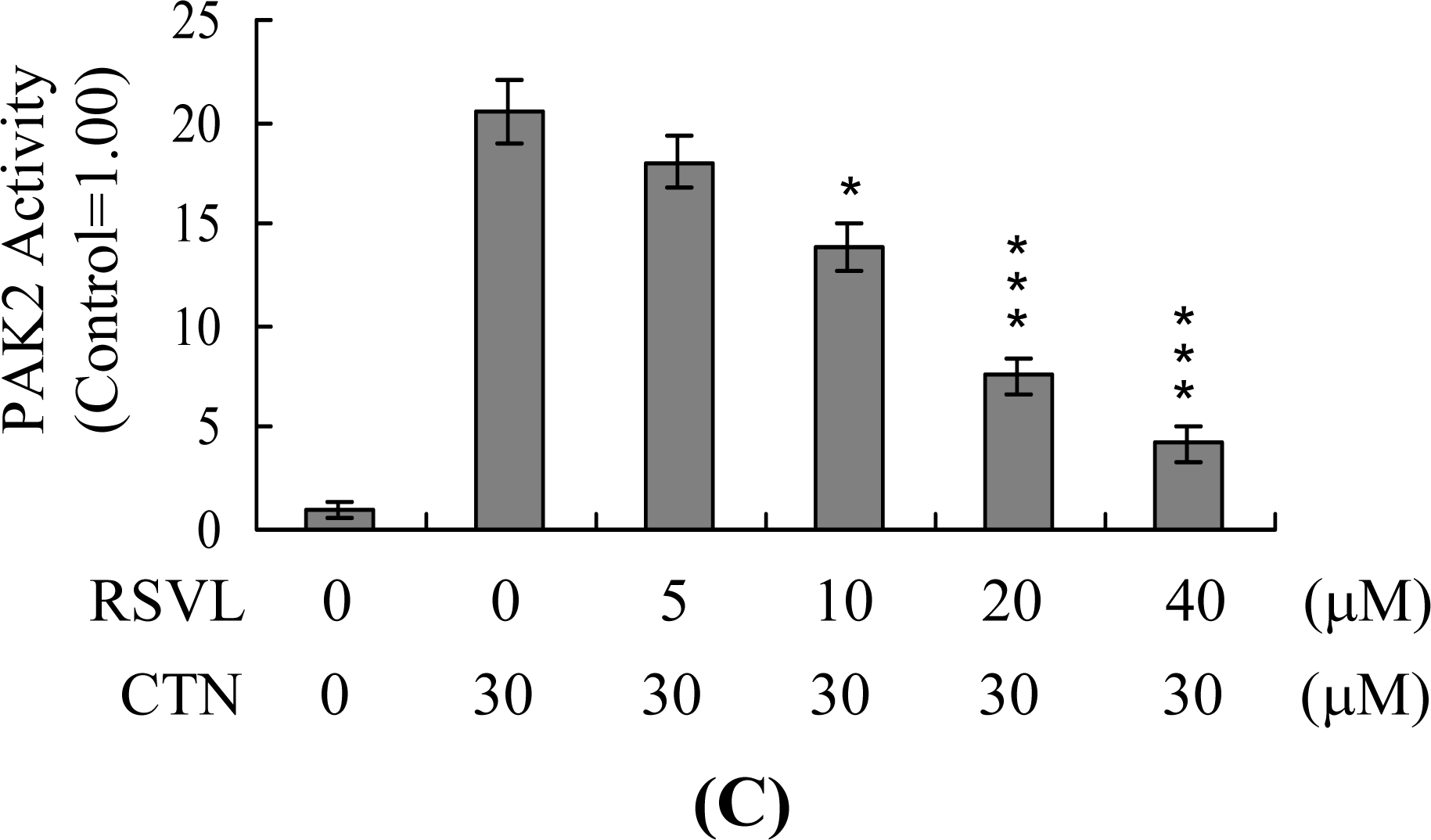
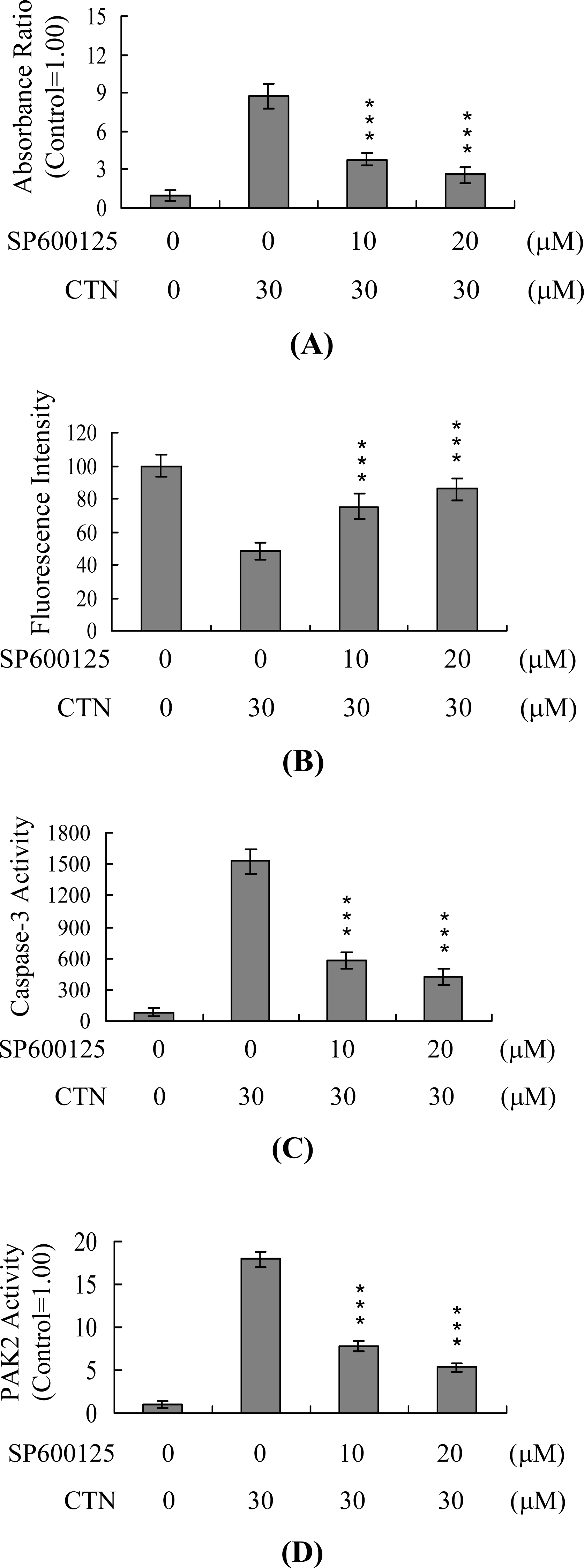
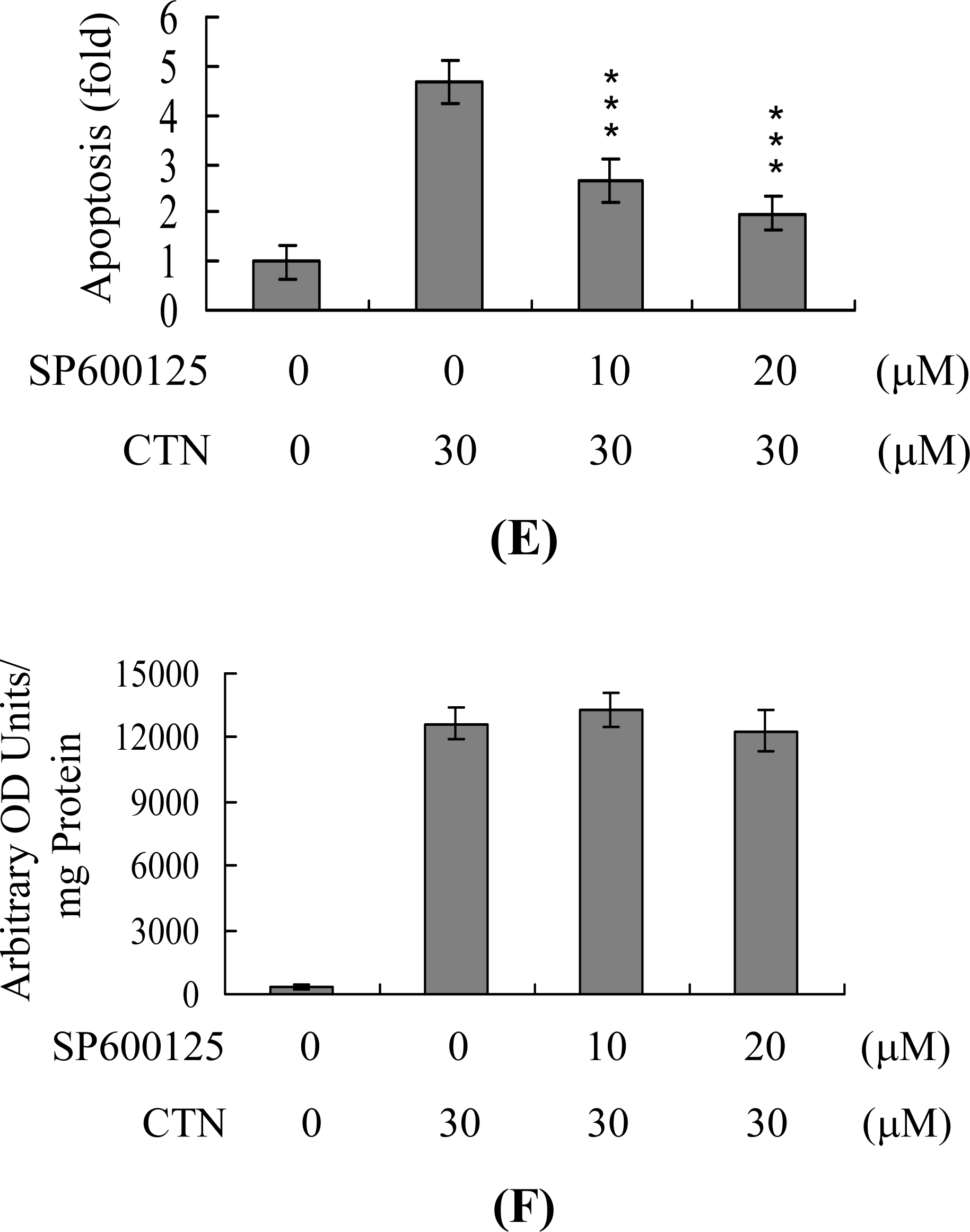
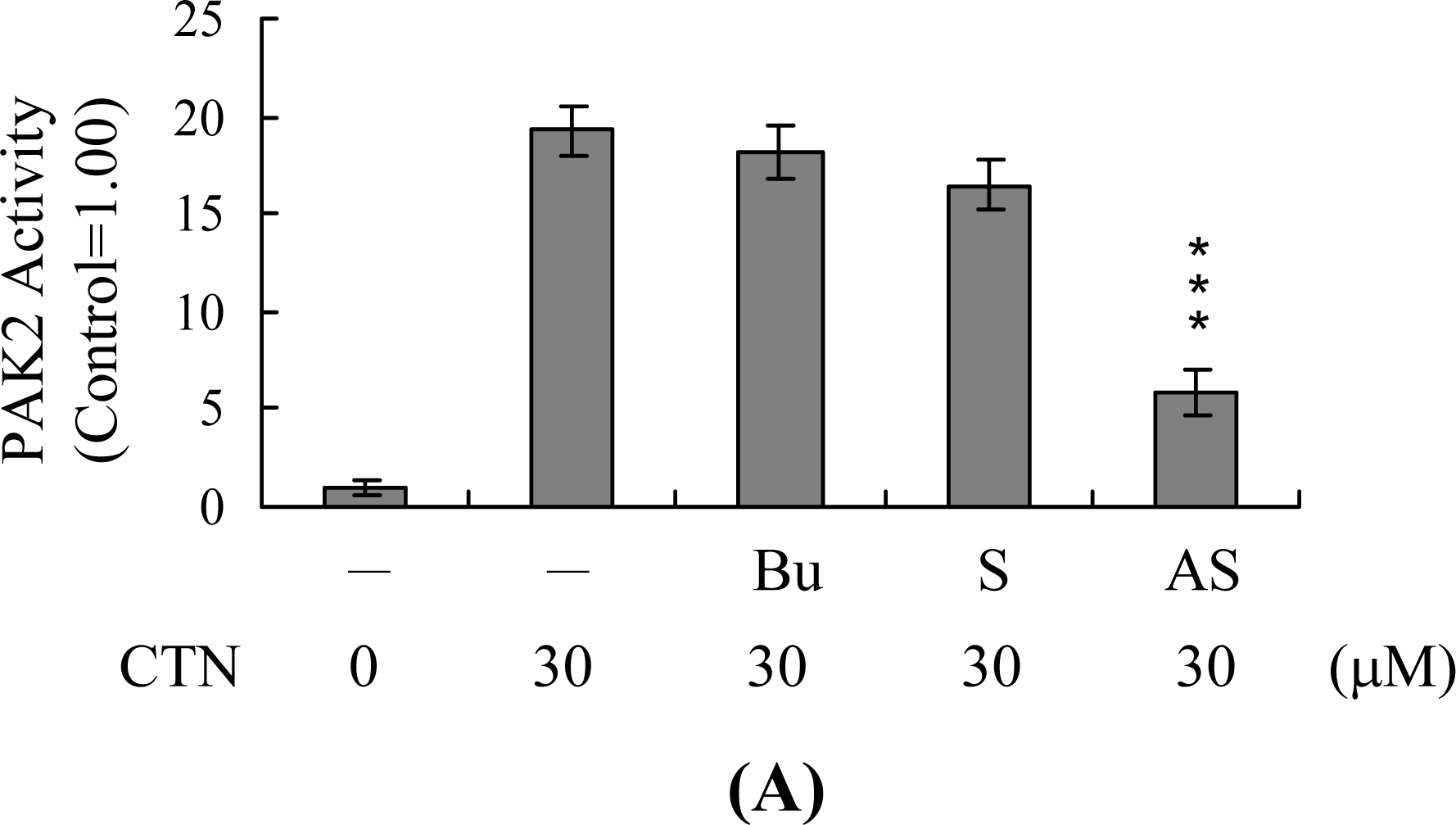
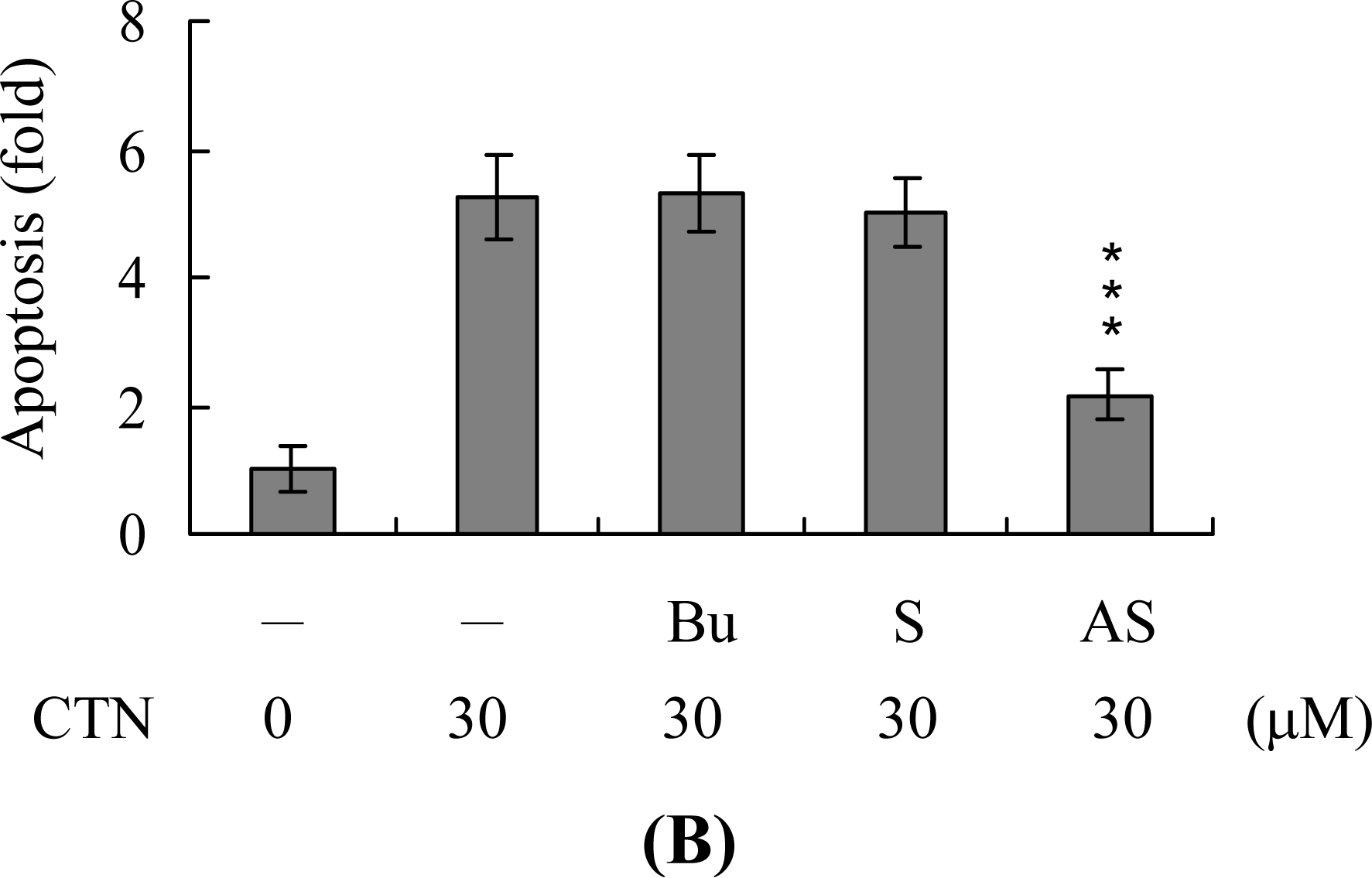
2. Results and Discussion
3. Experimental Section
3.1. Chemicals
3.2. Cell Culture and CTN Treatment
3.3. MTT Assay
3.4. Assessment of Apoptosis and Necrosis
3.5. ROS Assay
3.6. JNK Assays
3.7. Detection of Mitochondrial Membrane Potential (MMP)
3.8. Caspase Activity Assays
3.9. Immunoprecipitation and PAK2 Activity Assay
3.10. Inhibition of PAK2 by Anti-Sense Oligonucleotides
3.11. Statistics
4. Conclusions
Acknowledgments
References and Notes
- Blanc, PJ; Laussac, JP; Le Bars, J; Le Bars, P; Loret, MO; Pareilleux, A; Prome, D; Prome, JC; Santerre, AL; Goma, G. Characterization of monascidin A from Monascus as citrinin. Int. J. Food Microbiol 1995, 27, 201–213. [Google Scholar]
- CAST Mycotoxins: Risks in Plant, Animal, and Human Systems, Council of Agricultural Science and Technology, Task force report No. 139, CAST, Ames, IA, 2003.
- Aleo, MD; Wyatt, RD; Schnellmann, RG. The role of altered mitochondrial function in citrinin-induced toxicity to rat renal proximal tubule suspensions. Toxicol. Appl. Pharmacol 1991, 109, 455–463. [Google Scholar]
- Kogika, MM; Hagiwara, MK; Mirandola, RM. Experimental citrinin nephrotoxicosis in dogs: Renal function evaluation. Vet. Hum. Toxicol 1993, 35, 136–140. [Google Scholar]
- Arai, M; Hibino, T. Tumorigenicity of citrinin in male F344 rats. Cancer Lett 1983, 17, 281–287. [Google Scholar]
- Yu, FY; Liao, YC; Chang, CH; Liu, BH. Citrinin induces apoptosis in HL-60 cells via activation of the mitochondrial pathway. Toxicol. Lett 2006, 161, 143–151. [Google Scholar]
- Chan, WH. Citrinin induces apoptosis via a mitochondria-dependent pathway and inhibition of survival signals in embryonic stem cells, and causes developmental injury in blastocysts. Biochem. J 2007, 404, 317–326. [Google Scholar]
- Chan, WH; Shiao, NH. Effect of citrinin on mouse embryonic development in vitro and in vivo. Reprod. Toxicol 2007, 24, 120–125. [Google Scholar]
- Halliwell, B; Gutteridge, JM. Role of free radicals and catalytic metal ions in human disease: An overview. Methods Enzymol 1990, 186, 1–85. [Google Scholar]
- Chan, WH. Ginkgolide B induces apoptosis and developmental injury in mouse embryonic stem cells and blastocysts. Hum. Reprod 2006, 21, 2985–2995. [Google Scholar]
- Chan, WH; Shiao, NH; Lu, PZ. CdSe quantum dots induce apoptosis in human neuroblastoma cells via mitochondrial-dependent pathways and inhibition of survival signals. Toxicol. Lett 2006, 167, 191–200. [Google Scholar]
- Martins, LM; Kottke, T; Mesner, PW; Basi, GS; Sinha, S; Frigon, N, Jr; Tatar, E; Tung, JS; Bryant, K; Takahashi, A; Svingen, PA; Madden, BJ; McCormick, DJ; Earnshaw, WC; Kaufmann, SH. Activation of multiple interleukin-1beta converting enzyme homologues in cytosol and nuclei of HL-60 cells during etoposide-induced apoptosis. J. Biol. Chem 1997, 272, 7421–7430. [Google Scholar]
- Nicholson, DW; Thornberry, NA. Caspases: Killer proteases. Trends Biochem. Sci 1997, 22, 299–306. [Google Scholar]
- Tsujimoto, Y; Shimizu, S. Bcl-2 family: Life-or-death switch. FEBS Lett 2000, 466, 6–10. [Google Scholar]
- Anderson, P. Kinase cascades regulating entry into apoptosis. Microbiol. Mol. Biol. Rev 1997, 61, 33–46. [Google Scholar]
- Xia, Z; Dickens, M; Raingeaud, J; Davis, RJ; Greenberg, ME. Opposing effects of ERK and JNK-p38 MAP kinases on apoptosis. Science 1995, 270, 1326–1331. [Google Scholar]
- Verheij, M; Bose, R; Lin, XH; Yao, B; Jarvis, WD; Grant, S; Birrer, MJ; Szabo, E; Zon, LI; Kyriakis, JM; Haimovitz-Friedman, A; Fuks, Z; Kolesnick, RN. Requirement for ceramide-initiated SAPK/JNK signalling in stress-induced apoptosis. Nature 1996, 380, 75–79. [Google Scholar]
- Seimiya, H; Mashima, T; Toho, M; Tsuruo, T. c-Jun NH2-terminal kinase-mediated activation of interleukin-1beta converting enzyme/CED-3-like protease during anticancer drug-induced apoptosis. J. Biol. Chem 1997, 272, 4631–4636. [Google Scholar]
- Hsuuw, YD; Chang, CK; Chan, WH; Yu, JS. Curcumin prevents methylglyoxal-induced oxidative stress and apoptosis in mouse embryonic stem cells and blastocysts. J. Cell. Physiol 2005, 205, 379–386. [Google Scholar]
- Chan, WH. Citrinin induces apoptosis in mouse embryonic stem cells. IUBMB Life 2008, 60, 171–179. [Google Scholar]
- Rudel, T; Bokoch, GM. Membrane and morphological changes in apoptotic cells regulated by caspase-mediated activation of PAK2. Science 1997, 276, 1571–1574. [Google Scholar]
- Lee, N; MacDonald, H; Reinhard, C; Halenbeck, R; Roulston, A; Shi, T; Williams, LT. Activation of hPAK65 by caspase cleavage induces some of the morphological and biochemical changes of apoptosis. Proc. Natl. Acad. Sci. USA 1997, 94, 13642–13647. [Google Scholar]
- Tang, TK; Chang, WC; Chan, WH; Yang, SD; Ni, MH; Yu, JS. Proteolytic cleavage and activation of PAK2 during UV irradiation-induced apoptosis in A431 cells. J. Cell. Biochem 1998, 70, 442–454. [Google Scholar]
- Chan, WH; Yu, JS; Yang, SD. PAK2 is cleaved and activated during hyperosmotic shock-induced apoptosis via a caspase-dependent mechanism: evidence for the involvement of oxidative stress. J. Cell. Physiol 1999, 178, 397–408. [Google Scholar]
- Chan, WH; Yu, JS; Yang, SD. Apoptotic signalling cascade in photosensitized human epidermal carcinoma A431 cells: Involvement of singlet oxygen, c-Jun N-terminal kinase, caspase-3 and p21-activated kinase 2. Biochem. J 2000, 351, 221–232. [Google Scholar]
- Chan, WH; Wu, HJ. Anti-apoptotic effects of curcumin on photosensitized human epidermal carcinoma A431 cells. J. Cell. Biochem 2004, 92, 200–212. [Google Scholar]
- Zhang, S; Han, J; Sells, MA; Chernoff, J; Knaus, UG; Ulevitch, RJ; Bokoch, GM. Rho family GTPases regulate p38 mitogen-activated protein kinase through the downstream mediator Pak1. J. Biol. Chem 1995, 270, 23934–23936. [Google Scholar]
- Frost, JA; Xu, S; Hutchison, MR; Marcus, S; Cobb, MH. Actions of Rho family small G proteins and p21-activated protein kinases on mitogen-activated protein kinase family members. Mol. Cell. Biol 1996, 16, 3707–3713. [Google Scholar]
- Huang, YT; Lai, CY; Lou, SL; Yeh, JM; Chan, WH. Activation of JNK and PAK2 is essential for citrinin-induced apoptosis in a human osteoblast cell line. Environ. Toxicol 2008, 24, 343–356. [Google Scholar]
- Chan, WH; Wu, HJ; Shiao, NH. Apoptotic signaling in methylglyoxal-treated human osteoblasts involves oxidative stress, c-Jun N-terminal kinase, caspase-3, and p21-activated kinase 2. J. Cell. Biochem 2007, 100, 1056–1069. [Google Scholar]
- Jang, M; Cai, L; Udeani, GO; Slowing, KV; Thomas, CF; Beecher, CW; Fong, HH; Farnsworth, NR; Kinghorn, AD; Mehta, RG; Moon, RC; Pezzuto, JM. Cancer chemopreventive activity of resveratrol, a natural product derived from grapes. Science 1997, 275, 218–220. [Google Scholar]
- Mgbonyebi, OP; Russo, J; Russo, IH. Antiproliferative effect of synthetic resveratrol on human breast epithelial cells. Int. J. Oncol 1998, 12, 865–869. [Google Scholar]
- Huang, C; Ma, WY; Goranson, A; Dong, Z. Resveratrol suppresses cell transformation and induces apoptosis through a p53-dependent pathway. Carcinogenesis 1999, 20, 237–242. [Google Scholar]
- Gusman, J; Malonne, H; Atassi, G. A reappraisal of the potential chemopreventive and chemotherapeutic properties of resveratrol. Carcinogenesis 2001, 22, 1111–1117. [Google Scholar]
- Frankel, EN; Waterhouse, AL; Kinsella, JE. Inhibition of human LDL oxidation by resveratrol. Lancet 1993, 341, 1103–1104. [Google Scholar]
- Pace-Asciak, CR; Rounova, O; Hahn, SE; Diamandis, EP; Goldberg, DM. Wines and grape juices as modulators of platelet aggregation in healthy human subjects. Clin. Chim. Acta 1996, 246, 163–182. [Google Scholar]
- Subbaramaiah, K; Chung, WJ; Michaluart, P; Telang, N; Tanabe, T; Inoue, H; Jang, M; Pezzuto, JM; Dannenberg, AJ. Resveratrol inhibits cyclooxygenase-2 transcription and activity in phorbol ester-treated human mammary epithelial cells. J. Biol. Chem 1998, 273, 21875–21882. [Google Scholar]
- Clement, MV; Hirpara, JL; Chawdhury, SH; Pervaiz, S. Chemopreventive agent resveratrol, a natural product derived from grapes, triggers CD95 signaling-dependent apoptosis in human tumor cells. Blood 1998, 92, 996–1002. [Google Scholar]
- Hsieh, TC; Wu, JM. Differential effects on growth, cell cycle arrest, and induction of apoptosis by resveratrol in human prostate cancer cell lines. Exp. Cell. Res 1999, 249, 109–115. [Google Scholar]
- Ahmad, N; Adhami, VM; Afaq, F; Feyes, DK; Mukhtar, H. Resveratrol causes WAF-1/p21-mediated G(1)-phase arrest of cell cycle and induction of apoptosis in human epidermoid carcinoma A431 cells. Clin. Cancer Res 2001, 7, 1466–1473. [Google Scholar]
- Surh, YJ; Hurh, YJ; Kang, JY; Lee, E; Kong, G; Lee, SJ. Resveratrol, an antioxidant present in red wine, induces apoptosis in human promyelocytic leukemia (HL-60) cells. Cancer Lett 1999, 140, 1–10. [Google Scholar]
- Madan, E; Prasad, S; Roy, P; George, J; Shukla, Y. Regulation of apoptosis by resveratrol through JAK/STAT and mitochondria mediated pathway in human epidermoid carcinoma A431 cells. Biochem. Biophys. Res. Commun 2008, 377, 1232–1237. [Google Scholar]
- Filomeni, G; Graziani, I; Rotilio, G; Ciriolo, MR. trans-Resveratrol induces apoptosis in human breast cancer cells MCF-7 by the activation of MAP kinases pathways. Genes Nutr 2007, 2, 295–305. [Google Scholar] [Green Version]
- Schlegel, J; Peters, I; Orrenius, S; Miller, DK; Thornberry, NA; Yamin, TT; Nicholson, DW. CPP32/apopain is a key interleukin 1 beta converting enzyme-like protease involved in Fas-mediated apoptosis. J. Biol. Chem 1996, 271, 1841–1844. [Google Scholar]
- Erhardt, P; Cooper, GM. Activation of the CPP32 apoptotic protease by distinct signaling pathways with differential sensitivity to Bcl-xL. J. Biol. Chem 1996, 271, 17601–17604. [Google Scholar]
- Chan, WH; Wu, HY; Chang, WH. Dosage effects of curcumin on cell death types in a human osteoblast cell line. Food Chem. Toxicol 2006, 44, 1362–1371. [Google Scholar]
- Manser, E; Leung, T; Salihuddin, H; Zhao, ZS; Lim, L. A brain serine/threonine protein kinase activated by Cdc42 and Rac1. Nature 1994, 367, 40–46. [Google Scholar]
- Martin, GA; Bollag, G; McCormick, F; Abo, A. A novel serine kinase activated by rac1/CDC42Hs-dependent autophosphorylation is related to PAK65 and STE20. Embo J 1995, 14, 1970–1978. [Google Scholar]
- Bennett, BL; Sasaki, DT; Murray, BW; O'Leary, EC; Sakata, ST; Xu, W; Leisten, JC; Motiwala, A; Pierce, S; Satoh, Y; Bhagwat, SS; Manning, AM; Anderson, DW. SP600125, an anthrapyrazolone inhibitor of Jun N-terminal kinase. Proc. Natl. Acad. Sci. USA 2001, 98, 13681–13686. [Google Scholar]
- Vrabcheva, T; Usleber, E; Dietrich, R; Martlbauer, E. Co-occurrence of ochratoxin A and citrinin in cereals from Bulgarian villages with a history of Balkan endemic nephropathy. J. Agric. Food Chem 2000, 48, 2483–2488. [Google Scholar]
- Liu, BH; Wu, TS; Su, MC; Chung, CP; Yu, FY. Evaluation of citrinin occurrence and cytotoxicity in Monascus fermentation products. J. Agric. Food Chem 2005, 53, 170–175. [Google Scholar]
- Sabater-Vilar, M; Maas, RF; Fink-Gremmels, J. Mutagenicity of commercial Monascus fermentation products and the role of citrinin contamination. Mutat. Res 1999, 444, 7–16. [Google Scholar]
- Wei, W; Li, C; Wang, Y; Su, H; Zhu, J; Kritchevsky, D. Hypolipidemic and anti-atherogenic effects of long-term Cholestin (Monascus purpureus-fermented rice, red yeast rice) in cholesterol fed rabbits. J. Nutr. Biochem 2003, 14, 314–318. [Google Scholar]
- Long, LH; Clement, MV; Halliwell, B. Artifacts in cell culture: rapid generation of hydrogen peroxide on addition of (−)-epigallocatechin, (−)-epigallocatechin gallate, (+)-catechin, and quercetin to commonly used cell culture media. Biochem. Biophys. Res. Commun 2000, 273, 50–53. [Google Scholar]
- Halliwell, B. Oxidative stress in cell culture: An under-appreciated problem? FEBS Lett 2003, 540, 3–6. [Google Scholar]
- Jang, JH; Surh, YJ. Protective effects of resveratrol on hydrogen peroxide-induced apoptosis in rat pheochromocytoma (PC12) cells. Mutat. Res 2001, 496, 181–190. [Google Scholar]
- Chan, WH. Effect of resveratrol on high glucose-induced stress in human leukemia K562 cells. J. Cell. Biochem 2005, 94, 1267–1279. [Google Scholar]
- Chan, WH; Chang, YJ. Dosage effects of resveratrol on ethanol-induced cell death in the human K562 cell line. Toxicol. Lett 2006, 161, 1–9. [Google Scholar]
- Martinez, J; Moreno, JJ. Effect of resveratrol, a natural polyphenolic compound, on reactive oxygen species and prostaglandin production. Biochem. Pharmacol 2000, 59, 865–870. [Google Scholar]
- Fulda, S; Debatin, KM. Sensitization for anticancer drug-induced apoptosis by the chemopreventive agent resveratrol. Oncogene 2004, 23, 6702–6711. [Google Scholar]
- Su, JL; Lin, MT; Hong, CC; Chang, CC; Shiah, SG; Wu, CW; Chen, ST; Chau, YP; Kuo, ML. Resveratrol induces FasL-related apoptosis through Cdc42 activation of ASK1/JNK-dependent signaling pathway in human leukemia HL-60 cells. Carcinogenesis 2005, 26, 1–10. [Google Scholar]
- Nicolini, G; Rigolio, R; Miloso, M; Bertelli, AA; Tredici, G. Anti-apoptotic effect of trans-resveratrol on paclitaxel-induced apoptosis in the human neuroblastoma SH-SY5Y cell line. Neurosci. Lett 2001, 302, 41–44. [Google Scholar]
- Huang, LH; Shiao, NH; Hsuuw, YD; Chan, WH. Protective effects of resveratrol on ethanol-induced apoptosis in embryonic stem cells and disruption of embryonic development in mouse blastocysts. Toxicology 2007, 242, 109–122. [Google Scholar]
- Coso, OA; Chiariello, M; Yu, JC; Teramoto, H; Crespo, P; Xu, N; Miki, T; Gutkind, JS. The small GTP-binding proteins Rac1 and Cdc42 regulate the activity of the JNK/SAPK signaling pathway. Cell 1995, 81, 1137–1146. [Google Scholar]
- Honda, T; Shimizu, K; Kawakatsu, T; Fukuhara, A; Irie, K; Nakamura, T; Matsuda, M; Takai, Y. Cdc42 and Rac small G proteins activated by trans-interactions of nectins are involved in activation of c-Jun N-terminal kinase, but not in association of nectins and cadherin to form adherens junctions, in fibroblasts. Genes Cells 2003, 8, 481–491. [Google Scholar]
- Rudel, T; Zenke, FT; Chuang, TH; Bokoch, GM. p21-activated kinase (PAK) is required for Fas-induced JNK activation in Jurkat cells. J. Immunol 1998, 160, 7–11. [Google Scholar]
- Behl, C; Davis, JB; Lesley, R; Schubert, D. Hydrogen peroxide mediates amyloid beta protein toxicity. Cell 1994, 77, 817–827. [Google Scholar]
- Reimann, EM; Walsh, DA; Krebs, EG. Purification and properties of rabbit skeletal muscle adenosine 3′,5′-monophosphate-dependent protein kinases. J. Biol. Chem 1971, 246, 1986–1995. [Google Scholar]

© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Chen, C.-C.; Chan, W.-H. Inhibition of Citrinin-Induced Apoptotic Biochemical Signaling in Human Hepatoma G2 Cells by Resveratrol. Int. J. Mol. Sci. 2009, 10, 3338-3357. https://doi.org/10.3390/ijms10083338
Chen C-C, Chan W-H. Inhibition of Citrinin-Induced Apoptotic Biochemical Signaling in Human Hepatoma G2 Cells by Resveratrol. International Journal of Molecular Sciences. 2009; 10(8):3338-3357. https://doi.org/10.3390/ijms10083338
Chicago/Turabian StyleChen, Chia-Chi, and Wen-Hsiung Chan. 2009. "Inhibition of Citrinin-Induced Apoptotic Biochemical Signaling in Human Hepatoma G2 Cells by Resveratrol" International Journal of Molecular Sciences 10, no. 8: 3338-3357. https://doi.org/10.3390/ijms10083338