Recent Progress of Flower Colour Modification by Biotechnology

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Recent Progress; Anthocyanin Biosynthesis and Flower Colour
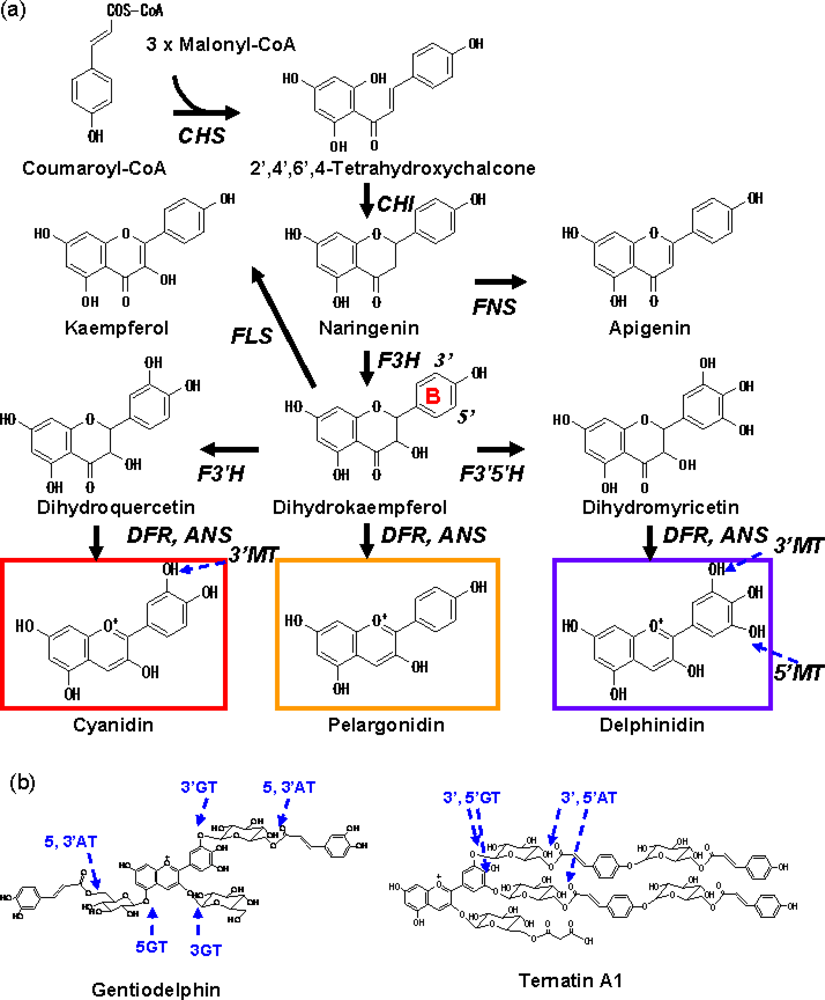
2.1. Anthocyanin Structure and Colour
2.2. Anthocyanin Modification
2.2.1. Glycosylation of Anthocyanins
2.2.2. Acylation of Anthocyanins
2.2.3. Methylation of Anthocyanins
2.3. Metal Ions
2.4. Regulation of Vacuolar pH
3. B-Ring Hydroxylation
3.1. F3′H and F3′5′H; Key Players of Flower Colour Modification
3.2. Natural Mutations of F3′H and F3′5′H
4. Flower Colour Modification Using Genetic Engineering
4.1. Lessons from Model Plant Experiments
4.2. Develpoment of the Moon® Series of Carnation
4.3. Regulatory Issues and Production of the Moon® Carnation
4.3.1. The Biology of the Parent Organism
4.3.2. The Modified Phenotype
4.3.3. Potential for Gene Flow from Transgenic Carnation to Native Plants
- Vegetative spread. Carnation does not spread vegetatively as it does not produce organs such as stolons, root-borne shoots, tubers, bulbs or runners. Cuttings must be struck in optimized conditions and roots do not form on discarded tissues. According to the numerous floras we have reviewed, including refuse sites [68] cultivated carnation has never been found growing wild, and efforts to locate wild carnation in production areas have been unsuccessful [59]. In experiments carried out in Colombia we have found that carnation deliberately planted in uncultivated habitats is unable to survive in competition with other plants.
- Formation of seed by a recipient plant, fertilized by transgenic pollen. Introgression of genes from cultivated crops to wild relatives is well known [69] and in the case of transgenic carnation, which carries a herbicide resistance gene, it has been necessary to evaluate the potential of introgression to wild Dianthus populations. This evaluation has concluded that there is no realistic potential for introgression because;
- In nature, Dianthus is insect-pollinated, and pollen is not spread by wind. Hybridization of Dianthus is only carried out by the Lepidoptera (butterflies, moths) which have proboscis long enough (up to 2.5 cm) to reach the nectaries, which are located right at the base of the flower in all Dianthus species [70]. Cultivated carnations are double-flowered and the many petals present on the flowers make it impossible for insects to access any pollen. No hybrid between carnation and any other Dianthus species has ever been recorded in the wild.
- Assuming, theoretically, an insect were to access a carnation flower in production, or in a vase in someone’s home, the probability of subsequently fertilizing a recipient, flowering, Dianthus plant is limited. The Dianthus genus is not native to the production areas in South America, and though Dianthus species related to carnation are occasionally found they are rare—no Dianthus species were found in surveys of flower growing areas [59].
- Formation and dispersal of seed. The process of seed development takes at least 5 weeks in carnation [73,74] and as cut flowers only survive 3 to 4 weeks at most in the hands of the consumer, seed development on a cut flower imported for consumption is not possible. In the production areas, the same argument can be raised, because the purpose of production is of course flower harvest, which in its own right eliminates the possibility of seed formation in the crop.
4.3.4. Comparative Trials
4.4. Transgenic Rose
4.4.1. Delphinidin Production and Flower Colour Modification
4.4.2. Regulatory Procedures of Transgenic Rose
4.5. Genetic Modification for Red Flowers
5. Concluding Remarks
Acknowledgments
References
- Tanaka, Y; Sasaki, N; Ohmiya, A. Plant pigments for coloration: Anthocyanins, betalains and carotenoids. Plant J 2008, 54, 733–749. [Google Scholar]
- Grotewold, E. The genetics and biochemistry of floral pigments. Annu. Rev. Plant Biol 2006, 57, 761–780. [Google Scholar]
- Tanaka, Y; Ohmiya, A. Seeing is believing: Engineering anthocyanin and carotenoid biosynthetic pathways. Curr. Opin. Biotechnol 2008, 19, 190–197. [Google Scholar]
- Yoshida, K; Mori, M; Kondo, T. Blue flower color development by anthocyanins: From chemical structure to cell physiology. Nat. Prod. Rep 2009, 26, 884–915. [Google Scholar]
- Veitch, NC; Grayer, RJ. Flavonoids and their glycosides, including anthocyanins. Nat. Prod. Rep 2008, 25, 555–611. [Google Scholar]
- Honda, T; Saito, N. Recent progress in the chemistry of polyacylated anthocyanins as flower color pigment. Heterocycles 2002, 56, 633–692. [Google Scholar]
- Yoshida, K; Toyama, Y; Kameda, K; Kondo, T. Contribution of each caffeoyl residue of the pigment molecules of gentiodelphin to blue color development. Phytochemistry 2000, 54, 85–92. [Google Scholar]
- Ogata, J; Kanno, Y; Itoh, Y; Tsugawa, H; Suzuki, M. Plant biochemistry: Anthocyanin biosynthesis in roses. Nature 2005, 435, 757–758. [Google Scholar]
- Tanaka, Y; Katsumoto, Y; Fukui, Y; Mizutani, M; Togami, J. Flavonoid glycosyltransferase and utilization thereof. . Patent Publication Number WO/2007/094521, August 23 2007. [Google Scholar]
- Kogawa, K; Kato, N; Kazuma, K; Noda, N; Suzuki, M. Purification and characterization of UDP-glucose: Anthocyanin 3′,5′-O-glucosyltransferase from Clitoria Ternatea. Planta 2007, 227, 1501–1509. [Google Scholar]
- Noda, N; Kato, N; Kogawa, K; Kazuma, K; Suzuki, M. Cloning and characterization of the gene encoding anthocyanin 3′,5′-O-glucosyltransferase involved in ternatin biosynthesis from blue petals of butterfly pea (Clitoria Ternatea). Plant Cell Physiol 2004, 45, s132. [Google Scholar]
- Fukuchi-Mizutani, M; Okuhara, H; Fukui, Y; Nakao, M; Katsumoto, Y; Yonekura-Sakakibra, K; Kusumi, T; Hase, T; Tanaka, Y. Biochemical and molecular characterization of a novel UDP-glucose: Anthocyanin 3′-O-glucosyltransferase, a key enzyme for blue anthocyanin biosynthesis, from gentian. Plant J 2003, 132, 1652–1663. [Google Scholar]
- Fujiwara, H; Tanaka, Y; Yonekura-Sakakibara, K; Fukuchi-Mizutani, M; Nakao, M; Fukui, Y; Yamaguchi, M; Ashikari, T; Kusumi, T. cDNA cloning, gene expression and subcellular localization of anthocyanin 5-aromatic acyltransferase from Gentiana Triflora. Plant J 1998, 16, 421–431. [Google Scholar]
- Tanaka, Y; Katsumoto, Y; Mizutani, M; Fukui, Y; Togami, J. Method of stabilizing, and bluing, of anthocyanin pigment using gene coding for transferase of aromatic acyl to 3′-position of anthocyanin. . Patent Publication Number WO/2006/046780, May 4 2006. [Google Scholar]
- Yonekura-Sakakibara, K; Tanaka, Y; Fukuchi-Mizutani, M; Fujiwara, H; Fukui, Y; Ashikari, T; Murakami, Y; Yamaguchi, M; Kusumi, T. Molecular and biochemical characterization of hydroxycinnamoyl-CoA: Anthocyanin 3-O-glucoside-6″-O-hydroxycinnamoyltransferase from Perilla Frutescens. Plant Cell Physiol 2000, 132, 1652–1663. [Google Scholar]
- Luo, J; Nishiyama, Y; Fuell, C; Taguchi, G; Elliott, K; Hill, L; Tanaka, Y; Kitayama, M; Yamazaki, M; Bailey, P; Parr, A; Michael, AJ; Saito, K; Martin, C. Convergent evolution in the BAHD family of acyl transferases: Identification and characterization of anthocyanin acyl transferases from Arabidopsis Thaliana. Plant J 2007, 50, 678–695. [Google Scholar]
- D’Auria, JC. Acyltransferases in plants: A good time to be BAHD. Curr. Opin. Plant. Biol 2006, 9, 331–340. [Google Scholar]
- Fraser, CM; Thompson, MG; Shirley, AM; Ralph, J; Schoenherr, JA; Sinlapadech, T; Hall, MC; Chapple, C. Related arabidopsis serine carboxypeptidase-like sinapoylglucose acyltransferases display distinct but overlapping substrate specificities. Plant Physiol 2007, 144, 19860–19999. [Google Scholar]
- Kazuma, K; Noda, N; Suzuki, M. Flavonoid composition related to petal color in different lines of Clitoria Ternatea. Phytochemistry 2003, 64, 1133–1139. [Google Scholar]
- Noda, N; Kazuma, K; Sasaki, T; Furukawa, K; Suzumi, M. . (Aomori prefecture) novel aromatic acyl group transferase gene. Patent Publication Number WO/2007/046148, April 26 2007.
- Abe, Y; Tera, M; Sasaki, N; Okamura, M; Umemoto, N; Momose, M; Kawahara, N; Kamakura, H; Goda, Y; Nagasawa, K; Ozeki, Y. Detection of 1-O-malylglucose: Pelargonidin 3-O-glucose-6-O-malyltransferase activity in carnation (Dianthus Caryophyllus). Biochem. Biophys. Res. Commun 2008, 373, 473–477. [Google Scholar]
- Umemoto, N; Momose, M. Novel protein having acyltransferase activity and gene encoding the same. . Patent Publication Number WO/2008/082006, July 10 2008. [Google Scholar]
- Brugliera, F; Linda, D; Koes, R; Tanaka, Y. Genetic sequences having methyltransferase activity and uses therefor. . Patent Publication Number WO/03/062428, July 31 2003. [Google Scholar]
- Hugueney, P; Provenzano, S; Verries, C; Ferrandino, A; Meudec, E; Batelli, G; Merdinoglu, D; Cheynier, V; Schubert, A; Ageorges, A. A novel cation-dependent O-methyltransferase involved in anthocyanin methylation in grapevine. Plant Physiol 2009, 150, 2057–2070. [Google Scholar]
- Takeda, K. Blue metal complex pigments involved in blue flower color. Proc. Jpn. Acad.: Ser. B 2006, 82, 142–154. [Google Scholar]
- Kondo, T; Yoshida, K; Nakagawa, A; Kawai, T; Tamura, H; Goto, T. Structural basis of blue-colour development in flower petals: Structure determination of commelinin from Commelina communis. Nature 1992, 358, 515–518. [Google Scholar]
- Shiono, M; Matsugaki, N; Takeda, K. Structure of commelinin, a blue complex pigment from the blue flowers of Commelina Communis. Proc. Jpn. Acad.: Ser. B 2008, 84, 452–456. [Google Scholar]
- Yoshida, K; Toyama-Kato, Y; Kameda, K; Kondo, T. Sepal color variation of Hydrangea macrophylla and vacuolar pH measured with a proton-selective microelectrode. Plant Cell Physiol 2003, 44, 262–268. [Google Scholar]
- Shoji, K; Miki, N; Nakajima, N; Monomoi, K; Kato, C; Yoshida, K. Perianth bottom-specific blue color development in Tulip cv. Murasakizuisho requires ferrc ions. Plant Cell Physiol 2007, 48, 243–251. [Google Scholar]
- Momonoi, K; Yoshida, K; Mano, S; Takahashi, H; Nakamori, C; Shoji, K; Nitta, A; Nishimura, M. A vacuolar iron transporter in tulip, TgVit1, is responsible for blue coloration in petal cells through iron accumulation. Plant J 2009, 59, 437–447. [Google Scholar]
- Yoshida, K; Kondo, T; Okazaki, Y; Katou, K. Cause of blue petal colour. Nature 1995, 373, 291. [Google Scholar]
- Fukada-Tanaka, S; Inagaki, Y; Yamaguchi, T; Saito, N; Iida, S. Colouring-enhancing protein in blue petals. Nature 2000, 407, 581. [Google Scholar]
- Yoshida, K; Miki, N; Nomonoi, K; Kawachi, M; Katou, K; Okazaki, Y; Uozumi, N; Maeshima, M; Kondo, T. Synchrony between flower opening and petal-color change from red to blue in morning glory, Ipomoea tricolor cv. Heavenly blue. Proc. Jpn. Acad. Ser. B 2009, 85, 187–197. [Google Scholar]
- Quattrocchio, F; Verweij, W; Kroon, A; Spelt, C; Mol, J; Koes, R. PH4 of petunia is an R2R3 MYB protein that activates vacuolar acidification through interactions with basic-helix-loop-helix transcription factors of the anthocyanin pathway. Plant Cell 2006, 18, 1274–1291. [Google Scholar]
- Verweij, W; Spelt, C; Di Sansebastiano, G-P; Vermeer, J; Reale, L; Ferranti, F; Koes, R; Quattrocchio, F. An H+ P-ATPase on the tonoplast determines vacuolar pH and flower colour. Nature Cell Biol 2008, 10, 1456–1462. [Google Scholar]
- Tanaka, Y. Flower colour and cytochromes P450. Phyochem. Rev 2006, 5, 283–291. [Google Scholar]
- Seitz, C; Eder, C; Deiml, B; Kellner, S; Martens, S; Forkmann, G. Cloning, functional identification and sequence analysis of flavonoid 3′-hydroxylase and flavonoid 3,5′-hydroxylase cDNA reveals independent evolution of flavonoid 3′,5′-hydroxylase in the Asteraceae family. Plant Mol. Biol 2006, 61, 365–381. [Google Scholar]
- Seitz, C; Ameres, S; Forkmann, G. Identification of the molecular basis for the functional difference between flavonoid 3′-hydroxylase and flavonoid 3′,5′-hydroxylase. FEBS Lett 2007, 581, 3429–3434. [Google Scholar]
- Hoshino, A; Morita, Y; Choi, JD; Saito, N; Toki, K; Tanaka, Y; Iida, S. Spontaneous mutations of the flavonoid 3′-hydroxylase gene conferring reddish flowers in the three morning glory species. Plant Cell Physiol 2003, 44, 990–1001. [Google Scholar]
- Zufall, RA; Rausher, MD. Genetic changes associated with floral adaptation restrict future evolutionary potential. Nature 2004, 428, 847–850. [Google Scholar]
- Nakatsuka, T; Nishihara, M; Mishiba, K; Hirano, H; Yamamura, S. Two different transposable elements inserted in flavonoid 3′,5′-hydroxylase gene contribute to pink flower coloration in Gentiana Scabra. Mol. Genet. Genomics 2006, 275, 231–241. [Google Scholar]
- Forkmann, G; Ruhau, B. Distinct substrate specificity of dihydroflavonol 4-reductase from flowers of Petunia Hybrida. Z. Naturforsch 1987, 42C, 1146–1148. [Google Scholar]
- Meyer, P; Heidemann, I; Forkmann, G; Saedler, H. A new petunia flower colour generated by transformation of a mutant with a maize gene. Nature 1987, 330, 677–678. [Google Scholar]
- Nakamura, N; Fukuchi-Mizutani, M; Suzuki, K; Miyazaki, K; Tanaka, Y. RNAi suppression of the anthocyanidin synthase gene in Torenia Hybrida yields white flowers with higher frequency and better stability than antisense and sense suppression. Plant Biotechnol 2006, 23, 13–17. [Google Scholar]
- Shukla, VK; Doyon, Y; Miller, JC; DeKelver, RC; Moehle, EA; Worden, SE; Mitchell, JC; Arnold, NL; Gopalan, S; Meng, X; Choi, VM; Rock, JM; Wu, YY; Katibah, GE; Zhifang, G; McCaskill, D; Simpson, MA; Blakeslee, B; Greenwalt, SA; Butler, HJ; Hinkley, SJ; Zhang, L; Rebar, EJ; Gregory, PD; Urnov, FD. Precise genome modification in the crop species Zea Mays using zinc-finger nucleases. Nature 2009, 459, 437–441. [Google Scholar]
- Townsend, JA; Wright, DA; Winfrey, RJ; Fu, F; Maeder, ML; Joung, JK; Voytas, DF. High-frequency modification of plant genes using engineered zinc-finger nucleases. Nature 2009, 459, 442–445. [Google Scholar]
- Holton, TA; Tanaka, Y. Transgenic flowering plants. . Patent Publication Number WO/94/28140, 1994. [Google Scholar]
- Holton, TA. Transgenic plants exhibiting altered flower color & methods for producing same. . Patent Publication Number WO/96/36716, 1996. [Google Scholar]
- Mol, J; Cornish, E; Mason, J; Koes, R. Novel coloured flowers. Curr. Opin. Biotechnol 1999, 10, 198–201. [Google Scholar]
- de Vetten, N; Ter Horst, J; Vab Schaik, H-P; de Boer, A; Mol, J; Koes, R. A cytochrome b5 is required for full activity of flavonoid 3′,5′-hydroxylase, a cytochrome P450 involved in the formation of blue flower colors. Proc. Natl. Acad. Sci. USA 1999, 96, 778–783. [Google Scholar]
- Brugliera, F; Tull, D; Holton, TA; Karan, M; Treloar, N; Simpson, K; Skurczynska, J; Mason, JG. Introduction of a cytochrome b5 enhances the activity of flavonoid 3′,5′-hydroxylase (a cytochrome P450) in transgenic carnation. Int Plant Mol Biol Rep (Suppl) 2000, 18, S6–S8. [Google Scholar]
- Tanaka, Y; Katsumoto, Y; Brugliera, F; John, M. Genetic engineering in floriculture. Plant Cell Tissue Organ Cult 2005, 80, 1–24. [Google Scholar]
- EFSA. Opinion of the Scientific Panel on Genetically Modified Organisms on a request from the Commission related to the notification (Reference C/NL/04/02) for the placing on the market of the genetically modified carnation Moonlite 123.2.38 with a modified colour, for import of cut flowers for ornamental use, under Part C of Directive 2001/18/EC from Florigene. EFSA J 2006, 362, 1–19. [Google Scholar]
- EFSA. Opinion of the Scientific Panel on Genetically Modified Organisms on a request from the Commission related to the notification (Reference C/NL/06/01) for the placing on the market of the genetically modified carnation Moonaqua 123.8.12 with a modified colour, for import of cut flowers for ornamental use, under Part C of Directive 2001/18/EC from Florigene. EFSA J 2008, 662, 1–25. [Google Scholar]
- Florigene Pty Ltd.
- Hughes, S. Carnations and Pinks, the Complete Guide; The Crowood Press: Marlborough, UK, 1991. [Google Scholar]
- Allwood, MC. Carnations, Pinks and All Dianthus, 4th ed; Allwood Bros Ltd: Sussex, UK, 1954. [Google Scholar]
- Holley, WD; Baker, R. Carnation Production: Including the History, Breeding, Culture and Marketing of Carnations; WC Brown Co. Ltd: Dubuque, IA, USA, 1963. [Google Scholar]
- Chandler, SF; Artunduaga, IRS; Jaramillo, LEL; Osorio, CAP. The production of transgenic carnation and rose in Colombia; A survey for wild populations and related species. 10th International Symposium on the Biosafety of Genetically Modified Organisms, Wellington, New Zealand, November 16–21, 2008.
- Sanchez-Guerrero, IM; Escudero, AI; Bartolom, B; Palacios, R. Occupational allergy to carnation. J. Allergy Clin. Immuno 1999, 104, 181–185. [Google Scholar]
- Vidal, C; Polo, F. Occupational allergy caused by Dianthus Caryophyllus, Gypsophila paniculara and Lilium longiflorum. Allergy 1998, 53, 995–998. [Google Scholar]
- Wu, X; Beecher, GR; Holden, JM; Haytowitz, DB; Gebhardt, SE; Prior, RL. Concentrations of anthocyanins in common foods in the United States and estimation of normal consumption. J. Agri. Food Chem 2006, 54, 4069–4075. [Google Scholar]
- Heinonen, M. Review: Antioxidant activity and antimicrobial effect of berry phenolics—A finnish perspective. Mol. Nutr. Food Res 2007, 51, 684–691. [Google Scholar]
- Catalano, G; Fossen, T; Andersen, ØM. Petunidin 3-O-β-rhamnopyranoside-5-O-β-glucopyranoside and other anthocyanins from flowers of Vicia Villosa. J. Agri. Food Chem 1998, 49, 4568–4570. [Google Scholar]
- Ando, T; Saito, N; Tatsuzawau, F; Kakefudau, T; Yamakageu, EK; Ohtani, T; Koshi-ishiu, M; MatsusakeuH, Y; Kokubunu, I; Watanabeu, H; Tsukamotou, T; Uedau, Y; Hashimoto, G; Marchesid, E; Asakura, K; Harar, R; Sekir, F. Floral anthocyanins in wild taxa of Petunia. Biochem. Syst. Ecol 1999, 27, 623–650. [Google Scholar]
- Chandler, SF; Dunfell, J. Gene flow, risk-assessment and the environmental release of transgenic plants. CRC Crit. Rev. Plant Sci 2008, 27, 25–49. [Google Scholar]
- Wilkinson, MJ; Ford, CS. Estimating the potential for ecological harm from gene flow to crop wild relatives. Collect. Biosafety Rev 2007, 3, 42–63. [Google Scholar]
- Kim, KD. Plant invasion and management in turf-dominated waste landfills in South Korea. Land Degrad. Develop 2002, 13, 257–267. [Google Scholar]
- Ellstrand, NC. Current knowledge of gene flow in plants: Implications for transgene flow. Phil. Trans. R. Soc. Lond. B 2003, 358, 1163–1170. [Google Scholar]
- Hickey, M; King, C. 100 Families of Flowering Plants; Cambridge University Press: Cambridge, UK, 1981. [Google Scholar]
- Mehlquist, GAL; Ober, D; Sagawa, Y. Somatic mutations in the carnation, Dianthus Caryophyllus. Proc. Natl. Acad. Sci. USA 1954, 40, 432–436. [Google Scholar]
- Kho, YO; Baer, J. The effect of temperature on pollen production in carnations. Euphytica 1973, 22, 467–470. [Google Scholar]
- Sparnaaij, LD; Beeger, GW. The improvement of seed production for breeding purposes in the glasshouse carnation (Dianthus Caryophyllus). Euphytica 1973, 22, 274–278. [Google Scholar]
- Gatt, MK; Hammett, KRW; Markham, KR; Murray, BG. Yellow pinks: Interspecific hybridization between Dianthus plumarius and related species with yellow flowers. Scientia Hort 1998, 77, 207–218. [Google Scholar]
- Brugliera, F; Tanaka, Y; Mason, J. Flavonoid 3′,5′-hydroxylase gene sequences and uses therefor. . Patent Publication Number US7612257, March 11 2004. [Google Scholar]
- Katsumoto, Y; Mizutani, M; Fukui, Y; Brugliera, F; Holton, T; Karan, M; Nakamura, N; Yonekura-Sakakibara, K; Togami, J; Pigeaire, A; Tao, G-Q; Nehra, N; Lu, C-Y; Dyson, B; Tsuda, S; Ashikari, T; Kusumi, T; Mason, J; Tanaka, Y. Engineering of the rose flavonoid biosynthetic pathway successfully generated blue-hued flowers accumulating delphinidin. Plant Cell Physiol 2007, 48, 1589–1600. [Google Scholar]
- Iwata, H. Protein and gene participating in perpetual blooming of angiosperm. . Patent Publication Number WO/2004/070036, 2004. [Google Scholar]
- Tsuda, S; Fukui, Y; Nakamura, N; Katsumoto, Y; Yonekura-Sakakibara, K; Fukuchi-Mizutani, M; Ohira, K; Ueyama, Y; Ohkawa, H; Holton, TA; Kusumi, T; Tanaka, Y. Flower color modification of Petunia Hybrida commercial varieties by metabolic engineering. Plant Biotechnol 2004, 21, 377–386. [Google Scholar]
- Nakatsuka, T; Abe, Y; Kakizaki, Y; Yamamura, S; Nishihara, M. Production of red-flowered plants by genetic engineering of multiple flavonoid biosynthetic genes. Plant Cell Rep 2007, 26, 1951–1959. [Google Scholar]
- Seitz, C; Vitten, M; Steinbach, P; Hartl, S; Hirsche, J; Rhthje, W; Treutter, D; Forkmann, G. Redirection of anthocyanin synthesis in Osteospermum Hybrida by a two-enzyme manipulation strategy. Phytochemistry 2007, 68, 824–833. [Google Scholar]
- Eason, JR; Debenham, M; McLachlan, A; Morgan, E. Novel red-flowered Gentiana: An emerging export cut flower crop from New Zealand. Acta Hort 2007, 755, 259–266. [Google Scholar]
- Mishiba, K; Nishihara, M; Nakatsuka, T; Abe, Y; Hirano, H; Yokoi, T; Kikuchi, A; Yamamura, S. Consistent transcriptional silencing of 35S-driven transgenes in gentian. Plant J 2005, 44, 541–556. [Google Scholar]
- Nakatsuka, T; Mishiba, KI; Kubota, A; Abe, Y; Yamamura, S; Nakamura, N; Tanaka, Y; Nishihara, M. Genetic engineering of novel flower colour by suppression of anthocyanin modification genes in gentian. J Plant Physiol 2009, in press.. [Google Scholar]
- Butelli, E; Titta, L; Giorgio, M; Mock, H-P; Matros, A; Peterek, S; Schijlen, EGWM; Hall, R; Bovy, AG; Luo, J; Martin, C. Enrichment of tomato fruit with health-promoting anthocyanins by expression of select transcription factors. Nature Biotechnol 2008, 26, 1301–1308. [Google Scholar]

© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Tanaka, Y.; Brugliera, F.; Chandler, S. Recent Progress of Flower Colour Modification by Biotechnology. Int. J. Mol. Sci. 2009, 10, 5350-5369. https://doi.org/10.3390/ijms10125350
Tanaka Y, Brugliera F, Chandler S. Recent Progress of Flower Colour Modification by Biotechnology. International Journal of Molecular Sciences. 2009; 10(12):5350-5369. https://doi.org/10.3390/ijms10125350
Chicago/Turabian StyleTanaka, Yoshikazu, Filippa Brugliera, and Steve Chandler. 2009. "Recent Progress of Flower Colour Modification by Biotechnology" International Journal of Molecular Sciences 10, no. 12: 5350-5369. https://doi.org/10.3390/ijms10125350