Influence of Thiazolidine-2,4-Dione Derivatives with Azolidine or Thiosemicarbazone Moieties on Haemophilus spp. Planktonic or Biofilm-Forming Cells


Abstract
:1. Introduction
2. Results and Discussion
2.1. Chemistry
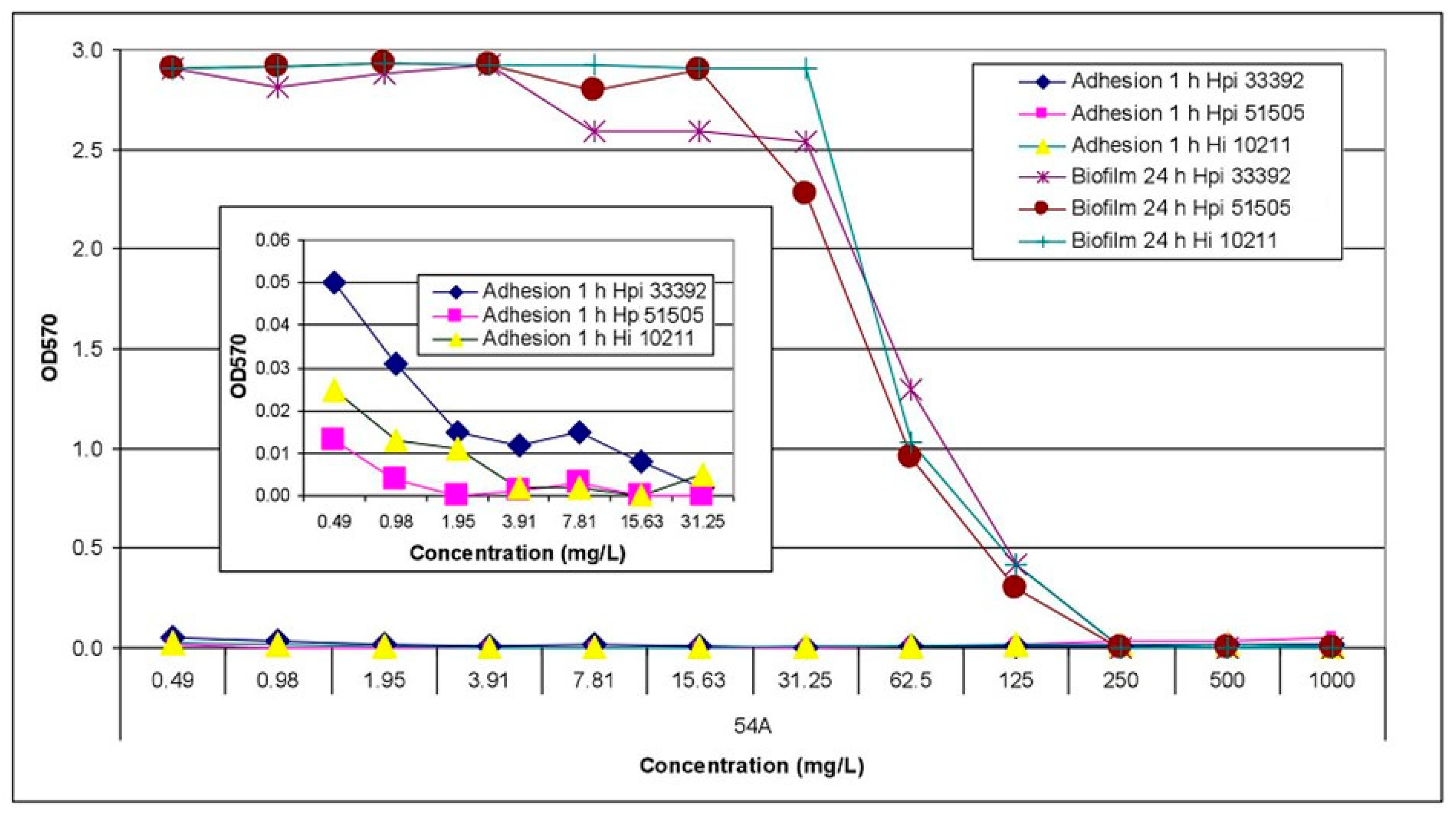
2.2. Activity Assay against Planktonic and Biofilm-Forming Cells
3. Materials and Methods
3.1. Chemistry
3.2. Bacterial Strains
3.3. Determination of the Minimal Inhibitory Concentration (MIC) of Tested TZD-Based Azolidine and Chlorophenyl Thiosemicarbazone Hybrids
3.4. Determination of the Minimal Biofilm Inhibitory Concentration (MBIC) of Tested TZD-Based Azolidine and Chlorophenyl Thiosemicarbazone Hybrids
3.5. Determination of the Anti-Adhesive Properties of Tested TZD-Based Azolidine and Chlorophenylthiosemicarbazone Hybrids
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Kaplan, J.B. Biofilm dispersal: Mechanisms, clinical implications, and potential therapeutic uses. J. Dent. Res. 2010, 89, 205–219. [Google Scholar] [CrossRef] [PubMed]
- Chernysh, S.; Gordya, N.; Tulin, D.; Yakovlev, A. Biofilm infections between Scylla and Charybdis: Interplay of host antimicrobial peptides and antibiotics. Infect. Drug Resist. 2018, 11, 501–514. [Google Scholar] [CrossRef] [PubMed]
- Jamal, M.; Ahmad, W.; Andleeb, S.; Jalil, F.; Imran, M.; Nawaz, M.A.; Hussain, T.; Ali, M.; Rafiq, M.; Kamil, M.A. Baterial biofilm and associated infections. J. Chin. Med. Assoc. 2018, 81, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Dickson, R.P.; Huang, Y.J.; Martinez, F.J.; Huffnagle, G.B. The lung microbiome and viral-induced exacerbations of chronic obstructive pulmonary disease: New observations, novel approaches. Am. J. Respir. Crit. Care Med. 2013, 188, 1185–1186. [Google Scholar] [CrossRef]
- Nørskov-Lauritsen, N. Classification, identification, and clinical significance of Haemophilus and Aggregatibacter species with host specificity for humans. Clin. Microbiol. Rev. 2014, 27, 214–240. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, A.; Murphy, T.F. Haemophilus influenzae infections in the H. influenzae type b conjugate vaccine era. J. Clin. Microbiol. 2011, 49, 3728–3732. [Google Scholar] [CrossRef] [PubMed]
- Choi, D.; Thermidor, M.; Cunha, B.A. Haemophilus parainfluenzae mitral prosthetic valve endocarditis in an intravenous drug abuser. Heart Lung 2005, 34, 152–154. [Google Scholar] [CrossRef]
- Mitchell, J.L.; Hill, S.L. Immune response to Haemophilus parainfluenzae in patients with chronic obstructive lung disease. Clin. Diagn. Lab. Immunol. 2000, 7, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Couturier, M.R.; Mehinovic, E.; Croft, A.C.; Fisher, M.A. Identification of HACEK clinical isolates by matrix-assisted laser desorption ionization–time of flight mass spectrometry. J. Clin. Microbiol. 2011, 49, 1104–1106. [Google Scholar] [CrossRef] [PubMed]
- Ebbing, R.; Robertson, C.F.; Robinson, P.J. Haemophilus influenzae and Haemophilus parainfluenzae in cystic fibrosis: 15 year experience. J. Med. Microbiol. Diagn. 2015, 5. [Google Scholar] [CrossRef]
- Kosikowska, U.; Rybojad, P.; Stępień-Pyśniak, D.; Żbikowska, A.; Malm, A. Changes in the prevalence and biofilm formation of Haemophilus influenzae and Haemophilus parainfluenzae from the respiratory microbiota of patients with sarcoidosis. BMC Infect. Dis. 2016, 16, 449. [Google Scholar] [CrossRef] [PubMed]
- Kosikowska, U.; Korona-Głowniak, I.; Niedzielski, A.; Malm, A. Nasopharyngeal and adenoid colonization by Haemophilus influenzae and Haemophilus parainfluenzae in children undergoing adenoidectomy and the ability of bacterial isolates to biofilm production. Medicine (Baltimore) 2015, 94, e799. [Google Scholar] [CrossRef]
- Lau, P.C.; Dutcher, J.R.; Beveridge, T.J.; Lam, J.S. Absolute quantitation of bacterial biofilm adhesion and viscoelasticity by microbead force spectroscopy. Biophys. J. 2009, 96, 2935–2948. [Google Scholar] [CrossRef]
- Available online: https://www.cdc.gov/hi-disease/clinicians.html (accessed on 7 March 2019).
- Tristram, S.; Jacobs, M.R.; Appelbaum, P.C. Antimicrobial resistance in Haemophilus influenzae. Clin. Microbiol. Rev. 2007, 20, 368–389. [Google Scholar] [CrossRef]
- Abotsi, R.E.; Govinden, U.; Essack, S.Y. Mechanisms of antibiotic resistance in Haemophilus parainfluenzae. S. Afr. J. Infect. Dis. 2017, 32, 111–114. [Google Scholar] [CrossRef]
- Jain, V.S.; Vora, D.K.; Ramaa, C.S. Thiazolidine-2,4-diones: Progress towards multifarious applications. Bioorg. Med. Chem. 2013, 21, 1599–1620. [Google Scholar] [CrossRef] [PubMed]
- Asati, V.; Mahapatra, D.K.; Bharti, S.K. Thiazolidine2,4-diones as multi-targed scaffold in medicinal chemistry: Potential anticancer agents. Eur. J. Med. Chem. 2014, 87, 814–833. [Google Scholar] [CrossRef]
- Trotsko, N.; Kosikowska, U.; Paneth, A.; Wujec, M.; Malm, A. Synthesis and antibacterial activity of new (2,4-dioxothiazolidin-5-yl/ylidene)acetic acid derivatives with thiazolidine-2,4-dione, rhodanine and 2-thiohydantoin moieties. Saudi Pharm. J. 2018, 26, 568–577. [Google Scholar] [CrossRef] [PubMed]
- Trotsko, N.; Kosikowska, U.; Paneth, A.; Plech, T.; Malm, A.; Wujec, M. Synthesis and antibacterial activity of new thiazolidine-2,4-dione-based chlorophenylthiosemicarbazone hybrids. Molecules 2018, 23, 1023. [Google Scholar] [CrossRef]
- Shenderovich, J.; Feldman, M.; Kirmayer, D.; Al-Quntar, A.; Steinberg, D.; Lavy, E.; Friedman, M. Local sustained-release delivery systems of the antibiofilm agent thiazolidinedione-8 for prevention of catheter-associated urinary tract infections. Int. J. Pharm. 2015, 485, 164–170. [Google Scholar] [CrossRef]
- Feldman, M.; Al-Quntar, A.; Polacheck, I.; Friedman, M.; Steinberg, D. Therapeutic potential of thiazolidinedione-8 as an antibiofilm agent against Candida albicans. PLoS ONE 2014, 9, e93225. [Google Scholar] [CrossRef] [PubMed]
- Feldman, M.; Shenderovich, J.; Al-Quntar, A.; Friedman, M.; Steinberg, D. Sustained Release of a Novel Anti-Quorum-Sensing Agent against Oral Fungal Biofilms. Antimicrob. Agents Chemother. 2015, 59, 2265–2272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feldman, M.; Ginsburg, I.; Al-Quntar, A.; Steinberg, D. Thiazolidinedione-8 alters symbiotic relationship in C. albicans- S. mutans dual species biofilm. Front. Microbiol. 2016, 7, 140. [Google Scholar] [CrossRef] [PubMed]
- Feldman, M.; Shenderovich, J.; Lavy, E.; Friedman, M.; Steinberg, D. A sustained-release membrane of thiazolidinedione-8: Effect on formation of a Candida/Bacteria mixed biofilm on hydroxyapatite in a continuous flow model. BioMed Res. Int. 2017, 2017, 3510124. [Google Scholar] [CrossRef] [PubMed]
- Kagan, S.; Jabbour, A.; Sionov, E.; Alquntar, A.A.; Steinberg, D.; Srebnik, M.; Nir-Paz, R.; Weiss, A.; Polacheck, I. Anti-Candida albicans biofilm effect of novel heterocyclic compounds. J. Antimicrob. Chemother. 2014, 69, 416–427. [Google Scholar] [CrossRef]
- Korem, M.; Kagan, S.; Polacheck, I. The effect of novel heterocyclic compounds on Cryptococcal biofilm. J. Fungi 2017, 3, 42. [Google Scholar] [CrossRef]
- Brackman, G.; Forier, K.; Al Quntar, A.A.A.; De Canck, E.; Enk, C.D.; Srebnik, M.; Braeckmans, K.; Coenye, T. Thiazolidinedione derivatives as novel agents against Propionibacterium acnes biofilms. J. Appl. Microbiol. 2013, 116, 492–501. [Google Scholar] [CrossRef]
- Coenye, T.; Nelis, H. In vitro and in vivo model systems to study microbial biofilm formation. J. Microbiol. Methods 2010, 83, 89–105. [Google Scholar] [CrossRef]
- Jallat, C.; Darfeuille-Michaud, A.; Joly, B.; Cluzel, R. Inhibition of Haemophilus influenzae adherence to buccal epithelial cells by cefuroxime. Chemotherapy 1992, 38, 428–432. [Google Scholar] [CrossRef] [PubMed]
- Starner, T.D.; Shrout, J.D.; Parsek, M.R.; Appelbaum, P.C.; Kim, G. Subinhibitory concentrations of azithromycin decrease nontypeable Haemophilus influenzae biofilm formation and Diminish established biofilms. Antimicrob. Agents Chemother. 2008, 52, 137–145. [Google Scholar] [CrossRef]
- Wojnicz, D.; Jankowski, S. Effects of subinhibitory concentrations of amikacin and ciprofloxacin on the hydrophobicity and adherence to epithelial cells of uropathogenic Escherichia coli strains. Int. J. Antimicrob. Agents 2007, 29, 700–704. [Google Scholar] [CrossRef]
- Fonseca, A.P.; Extremina, C.; Fonseca, A.F.; Sousa, J.C. Effect of subinhibitory concentration of piperacillin/tazobactam on Pseudomonas aeruginosa. J. Med. Microbiol. 2004, 53, 903–910. [Google Scholar] [CrossRef]
- Vidya, K.C.; Mallya, P.S.; Rao, P.S. Inhibition of bacterial adhesion by subinhibitory concentrations of antibiotics. Indian J. Med. Microbiol. 2005, 23, 102–105. [Google Scholar]
- Høiby, N.; Bjarnsholt, T.; Moser, C.; Bassi, G.L.; Coenye, T.; Donelli, G.; Hall-Stoodley, L.; Hola, V.; Imbert, C.; Kirketerp-Møller, K.; et al. ESCMID guidline for the diagnosis and biofilm infections 2014. Clin. Microbiol. Infect. 2015, 21 (Suppl. 1), S1–S25. [Google Scholar] [CrossRef]
- European Committee for Antimicrobial Susceptibility Testing (EUCAST) of the European Society for Clinical Microbiology and Infectious Diseases (ESCMID). EUCAST Discussion Document E. Dis. 5.1: Determination of minimum inhibitory concentrations (MICs) of antibacterial agents by broth dilution. Clin. Microbiol. Infect. 2003, 9, 1–7. [Google Scholar]
- Kosikowska, U.; Andrzejczuk, S.; Plech, T.; Malm, A. Inhibitory effect of 1,2,4-triazole-ciprofloxacin hybrids on Haemophilus parainfluenzae and Haemophilus influenzae biofilm formation in vitro under stationary conditions. Res. Microbiol. 2016, 167, 647–654. [Google Scholar] [CrossRef]
- Kosikowska, U.; Malm, A.; Pitucha, M.; Rajtar, B.; Polz-Dacewicz, M. Inhibitory effect of N-ethyl-3-amino-5-oxo-4-phenyl-2,5-dihydro-1H-pyrazole-1-carbothioamide on Haemophilus spp. planktonic or biofilm-forming cells. Med. Chem. Res. 2014, 23, 1057–1066. [Google Scholar] [CrossRef]
- Černohorská, L.; Votava, M. Antibiotic synergy against biofilm-forming Pseudomonas aeruginosa. Folia Microbiol. 2008, 53, 57–60. [Google Scholar] [CrossRef]
- Sekiya, R.; Karakida, K.; Arai, T.; Sato, Y.; Sakamoto, Y.; Kanai, N.; Maruyama, R.; Kaneko, A. In vitro inhibitory effects of various antimicrobial and antifungal agents on biofilm formation of microorganisms isolated from odontogenic infections. Oral Ther. Pharmacol. 2010, 29, 69–76. [Google Scholar]
Sample Availability: Samples of the compounds 34A, 39A, 41A, 47A, 51A, 52A, 54A, 56A–60A, 64A and 19B–24B, 26B, 29B, 33B, 34B are available from the authors. |


{kind=link}

| Compound 1 | Z | X | R |  | Compound Name |
|---|---|---|---|---|---|
| 34A | O | S | Br | single bond | 2-Bromo-4-[(2,4-dioxo-1,3-thiazolidin-5-ylidene)methyl]phenyl (2,4-dioxo-1,3-thiazolidin-5-yl)acetate |
| 39A | S | S | OC2H5 | single bond | 2-Ethoxy-4-[(4-oxo-2-thioxo-1,3-thiazolidin-5-ylidene)methyl]phenyl (2,4-dioxo-1,3-thiazolidin-5-yl)acetate |
| 41A | S | S | Br | single bond | 2-Bromo-4-[(4-oxo-2-thioxo-1,3-thiazolidin-5-ylidene)methyl]phenyl (2,4-dioxo-1,3-thiazolidin-5-yl)acetate |
| 47A | S | NH | Br | single bond | 2-Bromo-4-[(5-oxo-2-thioxoimidazolidin-4-ylidene)methyl]phenyl (2,4-dioxo-1,3-thiazolidin-5-yl)acetate |
| 51A | O | S | OCH3 | double bond | 4-[(2,4-Dioxo-1,3-thiazolidin-5-ylidene)methyl]-2-methoxyphenyl (2,4-dioxo-1,3-thiazolidin-5-ylidene)acetate |
| 52A | O | S | OC2H5 | double bond | 4-[(2,4-Dioxo-1,3-thiazolidin-5-ylidene)methyl]-2-ethoxyphenyl (2,4-dioxo-1,3-thiazolidin-5-ylidene)acetate |
| 54A | O | S | Br | double bond | 2-Bromo-4-[(2,4-dioxo-1,3-thiazolidin-5-ylidene)methyl]phenyl (2,4-dioxo-1,3-thiazolidin-5-ylidene)acetate |
| 56A | - | - | - | - | 3-[(4-Oxo-2-thioxo-1,3-thiazolidin-5-ylidene)methyl]phenyl (2,4-dioxo-1,3-thiazolidin-5-ylidene)acetate |
| 57A | S | S | H | double bond | 4-[(4-Oxo-2-thioxo-1,3-thiazolidin-5-ylidene)methyl]phenyl (2,4-dioxo-1,3-thiazolidin-5-ylidene)acetate |
| 58A | S | S | OCH3 | double bond | 2-Methoxy-4-[(4-oxo-2-thioxo-1,3-thiazolidin-5-ylidene)methyl]phenyl (2,4-dioxo-1,3-thiazolidin-5-ylidene)acetate |
| 59A | S | S | OC2H5 | double bond | 2-Ethoxy-4-[(4-oxo-2-thioxo-1,3-thiazolidin-5-ylidene)methyl]phenyl (2,4-dioxo-1,3-thiazolidin-5-ylidene)acetate |
| 60A | S | S | Cl | double bond | 2-Chloro-4-[(4-oxo-2-thioxo-1,3-thiazolidin-5-ylidene)methyl]phenyl (2,4-dioxo-1,3-thiazolidin-5-ylidene)acetate |
| 64A | S | NH | OC2H5 | double bond | 2-Ethoxy-4-[(5-oxo-2-thioxoimidazolidin-4-ylidene)methyl]phenyl (2,4-dioxo-1,3-thiazolidin-5-ylidene)acetate |

| Compound 1 | R | R1 | | Compound Name |
|---|---|---|---|---|
| 19B | - | - | - | 3-[{2-[(2,6-Dichlorophenyl)carbamothioyl]hydrazinylidene}methyl]phenyl (2,4-dioxo-1,3-thiazolidin-5-yl)acetate |
| 20B | H | 2-Cl | single bond | 4-[{2-[(2-Chlorophenyl)carbamothioyl]hydrazinylidene}methyl]phenyl (2,4-dioxo-1,3-thiazolidin-5-yl)acetate |
| 21B | H | 3-Cl | single bond | 4-[{2-[(3-Chlorophenyl)carbamothioyl]hydrazinylidene}methyl]phenyl (2,4-dioxo-1,3-thiazolidin-5-yl)acetate |
| 22B | H | 4-Cl | single bond | 4-[{2-[(4-Chlorophenyl)carbamothioyl]hydrazinylidene}methyl]phenyl (2,4-dioxo-1,3-thiazolidin-5-yl)acetate |
| 23B | H | 2,4-diCl | single bond | 4-[{2-[(2,4-Dichlorophenyl)carbamothioyl]hydrazinylidene}methyl]phenyl (2,4-dioxo-1,3-thiazolidin-5-yl)acetate |
| 24B | H | 2,6-diCl | single bond | 4-[{2-[(2,6-Dichlorophenyl)carbamothioyl]hydrazinylidene}methyl]phenyl (2,4-dioxo-1,3-thiazolidin-5-yl)acetate |
| 26B | OCH3 | 3-Cl | single bond | 4-[{2-[(3-Chlorophenyl)carbamothioyl]hydrazinylidene}methyl]-2-methoxyphenyl (2,4-dioxo-1,3-thiazolidin-5-yl)acetate |
| 29B | OCH3 | 2,6-diCl | single bond | 4-[{2-[(2,6-Dichlorophenyl)carbamothioyl]hydrazinylidene}methyl]-2-methoxyphenyl (2,4-dioxo-1,3-thiazolidin-5-yl)acetate |
| 33B | H | 2-Cl | double bond | 4-[{2-[(2-Chlorophenyl)carbamothioyl]hydrazinylidene}methyl]phenyl (2,4-dioxo-1,3-thiazolidin-5-ylidene)acetate |
| 34B | H | 3-Cl | double bond | 4-[{2-[(3-Chlorophenyl)carbamothioyl]hydrazinylidene}methyl]phenyl (2,4-dioxo-1,3-thiazolidin-5-ylidene)acetate |
| Reference Species | Clinical Isolates | ||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Compound | Haemophilus parainfluenzae ATCC 33392 | Haemophilus parainfluenzae ATCC 51505 | Haemophilus influenzae ATCC 10211 | Haemophilus parainfluenzae 128 | Haemophilus parainfluenzae 134 | Haemophilus parainfluenzae 201 | Haemophilus parainfluenzae 206 | ||||||||||||||
| Antibacterial Activity (mg/L) | |||||||||||||||||||||
| MIC | MBIC | MBIC/ MIC | MIC | MBIC | MBIC/ MIC | MIC | MBIC | MBIC/ MIC | MIC | MBIC | MBIC/ MIC | MIC | MBIC | MBIC/ MIC | MIC | MBIC | MBIC/ MIC | MIC | MBIC | MBIC/ MIC | |
| 34A | 125 | 250 | 2 | 125 | 250 | 2 | 125 | 250 | 2 | >1000 | >1000 | - | >1000 | >1000 | - | 1000 | 250 | 0.25 | >1000 | 1000 | - |
| 39A | 500 | >1000 | - | 500 | 1000 | 2 | 500 | >1000 | - | >1000 | >1000 | - | >1000 | 1000 | - | 250 | 125 | 0.5 | >1000 | 1000 | - |
| 41A | 125 | 250 | 2 | 125 | 250 | 2 | 125 | 250 | 2 | >1000 | 500 | - | >1000 | >1000 | - | 500 | 250 | 0.5 | >1000 | >1000 | - |
| 47A | 500 | 1000 | 2 | 500 | 1000 | 2 | 500 | 1000 | 2 | >1000 | 500 | - | >1000 | 1000 | - | 1000 | 62.5 | 0.0625 | >1000 | 1000 | - |
| 51A | 500 | 500 | 1 | 500 | 500 | 1 | 500 | 500 | 1 | >1000 | 500 | - | >1000 | 1000 | - | 1000 | >1000 | - | >1000 | 500 | - |
| 52A | 500 | 1000 | 2 | 500 | 1000 | 2 | 500 | 1000 | 2 | >1000 | 1000 | - | >1000 | 500 | - | 250 | 125 | 0.5 | >1000 | 500 | - |
| 54A | 125 | 250 | 2 | 125 | 250 | 2 | 62.5 | 250 | 4 | >1000 | >1000 | - | >1000 | >1000 | - | 500 | 125 | 0.25 | >1000 | 1000 | - |
| 56A | 125 | 250 | 2 | 125 | 250 | 2 | 125 | 250 | 2 | >1000 | >1000 | - | >1000 | 250 | - | 1000 | 125 | 0.125 | >1000 | 250 | - |
| 57A | 125 | 1000 | 8 | 62.5 | 1000 | 16 | 500 | >1000 | - | >1000 | >1000 | - | >1000 | >1000 | - | 1000 | 125 | 0.125 | >1000 | 250 | - |
| 58A | 250 | 1000 | 4 | 250 | 1000 | 4 | 250 | 1000 | 4 | >1000 | >1000 | - | >1000 | 1000 | - | 500 | >1000 | - | >1000 | 1000 | - |
| 59A | 500 | 500 | 1 | 500 | 500 | 1 | 500 | 500 | 1 | >1000 | >1000 | - | >1000 | >1000 | - | 1000 | 125 | 0.125 | 1000 | >1000 | - |
| 60A | 125 | 250 | 2 | 125 | 250 | 2 | 125 | 250 | 2 | >1000 | >1000 | - | >1000 | >1000 | - | 1000 | 62.5 | 0.0625 | >1000 | 1000 | - |
| 64A | 250 | >1000 | - | 1000 | >1000 | - | 1000 | >1000 | - | >1000 | 1000 | - | >1000 | >1000 | - | 1000 | 125 | 0.125 | 250 | >1000 | - |
| 19B | >1000 | 500 | - | >1000 | >1000 | - | >1000 | 500 | - | >1000 | >1000 | - | >1000 | 500 | - | 125 | 125 | 1 | >1000 | 1000 | - |
| 20B | >1000 | >1000 | - | >1000 | >1000 | - | >1000 | >1000 | - | >1000 | >1000 | - | >1000 | 1000 | - | 500 | 1000 | 2 | >1000 | 1000 | - |
| 21B | >1000 | >1000 | - | >1000 | >1000 | - | >1000 | >1000 | - | >1000 | >1000 | - | >1000 | 1000 | - | 125 | 1000 | 8 | >1000 | 1000 | - |
| 22B | >1000 | >1000 | - | >1000 | >1000 | - | >1000 | >1000 | - | >1000 | >1000 | - | >1000 | 500 | - | 31.25 | 1000 | 32 | >1000 | 1000 | - |
| 23B | 1000 | >1000 | - | >1000 | >1000 | - | >1000 | >1000 | - | 1000 | 1000 | 1 | >1000 | 1000 | - | 125 | 1000 | 8 | >1000 | 1000 | - |
| 24B | >1000 | >1000 | - | >1000 | >1000 | - | >1000 | >1000 | - | >1000 | >1000 | - | >1000 | >1000 | - | 125 | 1000 | 8 | >1000 | 1000 | - |
| 26B | >1000 | >1000 | - | >1000 | >1000 | - | >1000 | >1000 | - | >1000 | >1000 | - | >1000 | 1000 | - | 100 | 1000 | 10 | >1000 | 1000 | - |
| 29B | >1000 | >1000 | - | >1000 | >1000 | - | >1000 | >1000 | - | >1000 | >1000 | - | >1000 | 1000 | - | 100 | 500 | 5 | >1000 | 1000 | - |
| 33B | >1000 | >1000 | - | >1000 | >1000 | - | >1000 | >1000 | - | >1000 | >1000 | - | >1000 | >1000 | - | 1000 | 1000 | 1 | >1000 | 1000 | - |
| 34B | 250 | >1000 | - | 500 | >1000 | - | 500 | >1000 | - | >1000 | >1000 | - | >1000 | >1000 | - | 500 | 250 | 0.5 | >1000 | 1000 | - |
| Ge | 250 | 250 | 1 | 1000 | 250 | 0.25 | 500 | 250 | 0.5 | 1000 | 1000 | 1 | 1000 | 500 | 0.5 | 500 | 250 | 0.5 | 1000 | 500 | 0.5 |
| Cef | 125 | 125 | 1 | 500 | 125 | 0.25 | 125 | 62.5 | 0.5 | 500 | 125 | 0.25 | 500 | 62.5 | 0.13 | 250 | 62.5 | 0.25 | 250 | 125 | 0.5 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trotsko, N.; Kosikowska, U.; Andrzejczuk, S.; Paneth, A.; Wujec, M. Influence of Thiazolidine-2,4-Dione Derivatives with Azolidine or Thiosemicarbazone Moieties on Haemophilus spp. Planktonic or Biofilm-Forming Cells. Molecules 2019, 24, 1051. https://doi.org/10.3390/molecules24061051
Trotsko N, Kosikowska U, Andrzejczuk S, Paneth A, Wujec M. Influence of Thiazolidine-2,4-Dione Derivatives with Azolidine or Thiosemicarbazone Moieties on Haemophilus spp. Planktonic or Biofilm-Forming Cells. Molecules. 2019; 24(6):1051. https://doi.org/10.3390/molecules24061051
Chicago/Turabian StyleTrotsko, Nazar, Urszula Kosikowska, Sylwia Andrzejczuk, Agata Paneth, and Monika Wujec. 2019. "Influence of Thiazolidine-2,4-Dione Derivatives with Azolidine or Thiosemicarbazone Moieties on Haemophilus spp. Planktonic or Biofilm-Forming Cells" Molecules 24, no. 6: 1051. https://doi.org/10.3390/molecules24061051