Comparison of the Inhibitory Activities of 5,6-Dihydroergosterol Glycoside α- and β-Anomers on Skin Inflammation

Department of Applied Chemistry and Global Center for Pharmaceutical Ingredient Materials, Kyung Hee University, Yongin 17410, Korea
*
Authors to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Molecules 2019, 24(2), 371; https://doi.org/10.3390/molecules24020371
Submission received: 5 December 2018
/
Revised: 12 January 2019
/
Accepted: 19 January 2019
/
Published: 21 January 2019
(This article belongs to the Special Issue Application of Organic Synthesis to Bioactive Compounds)
Abstract
:Chronic skin inflammatory diseases, such as atopic dermatitis, are associated with a dysfunctional skin barrier due to an increase in various inflammatory stimuli, for instance inflammatory cytokines and chemokines. In particular, CCL17 and CCL22 expression is increased in patients with chronic skin inflammation. In this study, we synthesized several α- and β-anomers of dihydroergosterol (DHE)-glycosides and assessed their effects on CCL17 and CCL22 expression. We confirmed that the β-anomers of DHE-glycosides were superior to α-anomers of DHE-glycosides in inhibiting CCL17 and CCL22 mRNA and protein expression. In addition, we determined that DHE-glycoside β-anomers showed strong inhibitory activity towards pro-inflammatory cytokine mRNA and protein expression, including that of TNF-α, IL-6, and IL-1β- in stimulated HaCaT cells. These results imply that DHE-glycoside α- and β-anomers should be separated during synthesis of drugs for chronic skin inflammation. Our results also suggest that β-anomers of DHE-glycosides may play an important role as new drugs for chronic skin inflammation because of their ability to inhibit the skin inflammatory biomarker proteins CCL17 and CCL22.
1. Introduction
Natural products from various plants with soap-like properties, such as saponins, have played an important role in medicine and daily life for thousands of years [1]. Saponins are amphipathic glycosides grouped structurally by having one or more hydrophilic glycoside moieties combined with a lipophilic triterpene or steroid derivative, and have well characterized biological properties including hemolytic activity, molluscicidal activity, and anti-inflammatory activity [2].
We previously confirmed that spinasterol-glucose (1) (spinasterol-Glc, Figure 1), extracted and identified from natural herbs, not only exhibited potent anti-inflammatory activity but also inhibited CCL17 mRNA and protein expression in TNF-α/INF-γ-stimulated HaCaT cells [3]. However, spinasterol-Glc is difficult to obtain from either natural plant extracts or using synthetic methods. The synthesis of spinasterol-Glc is very difficult because its steroidal skeleton, α-spinasterol, is not readily available. Recently, synthesis of α-spinasterol from stigmasterol using a double bond migration method has been reported; however, the synthesis was still difficult and inefficient [4,5]. Therefore, we previously designed and synthesized a new sterol, 5,6-dihydroergosterol (DHE or stellarsterol) that is structurally similar to spinasterol and more easily obtained. In previous studies, we demonstrated that DHE and its glycosides have comparable anti-inflammatory activity to spinasterol-Glc. Of the synthesized DHE-glycosides, the β-anomer of DHE-Glc (5,6-dihydroergosterol-glucose (2) showed the greatest anti-inflammatory activity [6]. In addition, we confirmed that the expression of CCL 17 and CCL22 mRNA and protein were strongly inhibited in a DNCB-induced atopic dermatitis (AD) animal model [7].
Chronic skin inflammatory diseases are associated with a dysfunctional skin barrier that is characterized by elevated expression of various inflammatory stimuli, specifically pro-inflammatory cytokines and chemokines [8,9]. In skin conditions like chronic inflammation, small signaling proteins, including chemokines, are secreted. The main function of these chemokines is to recruit leukocytes, monocytes, and neutrophils to inflammatory sites [10,11]. Specifically, in abnormal skin conditions like AD, chemokines have been reported to promote the secretion of pro-inflammatory cytokines and thymic stromal lymphopoietin (TSLP) proteins that cause itching [12]. Thymus and activation-regulated chemokine (TARC/CCL17) are a type of C-C chemokine that uses CC chemokine receptor 4 (CCR4) as a receptor, and specifically binds to and induces chemotaxis in Th2 cells and is associated with Th2 cell-mediated inflammatory diseases, such as AD [13]. Another CC-chemokine, macrophage-derived chemokine (MDC/CCL22), shares homology (37% amino acid identity) with CCL17, and both chemokines mediate a variety of biological activities through CCR4 [14]. Previous reports on the various pharmacological and medicinal functions of CCL17 and CCL22 in skin inflammation suggest that these chemokines can be used as biomarkers of chronic skin inflammation [15,16,17]. Furthermore, CCL17 has been shown to be present at high levels in the blood of AD patients and highly expressed in skin lesions [15]. In addition, CCL22 expression was increased in the epidermis and serum of patients with chronic inflammatory skin diseases [16]. These results indicate that CCL17 and CCL22 play crucial roles in the mobilization of Th2 cells and that increased expression of these chemokines further exacerbates chronic skin inflammation in diseases like AD [18].
Generally, glycosides consist of glycone (sugar) and aglycone (non-sugar) components, and can be divided into alpha (α) and beta (β) anomers depending on the linking stereochemistry of the glycone and aglycone portions (Figure 1). In our synthesis process, we have always obtained both the α- and β-anomers of DHE-glycosides, particularly during the glycosylation step. Since there have been few reports describing the relative bioactivity data of α- and β-anomers of DHE-glycosides, we were interested in determining their relative biological activities. To do this, we compared the inhibitory activities of α- and β-DHE-glycosides on TNF-α/IFN-γ-induced expression of pro-inflammatory cytokines and chemokines, including CCL17 and CCL22, in TNF-α/IFN-γ-stimulated human keratinocytes. Here, we report the synthesis and anti-inflammatory activity of α- and β-anomers of DHE-glycosides, and which anomer of DHE-glycosides more effectively inhibits the gene expression of pro-inflammatory cytokines IL1-b, IL-6, and TNF-α, and the chemokines CCL17 and CCL22, in HaCaT cells. Our results indicate that β-anomers of DHE-glycosides inhibit the mRNA and protein expression of pro-inflammatory cytokines and chemokines in TNF-α/IFN-γ-activated HaCaT cells to a greater extent than the α-anomers of DHE-glycosides, and that this is likely because of the instability in solution of the α-anomers of DHE-glycosides. These findings may contribute to the development of useful therapeutic agents for chronic skin inflammation diseases such as atopic dermatitis.
2. Results
2.1. Synthesis of α- and β-Anomers of 5,6-Dihydroergosterol-Glycosides (DHE-Glycosides)
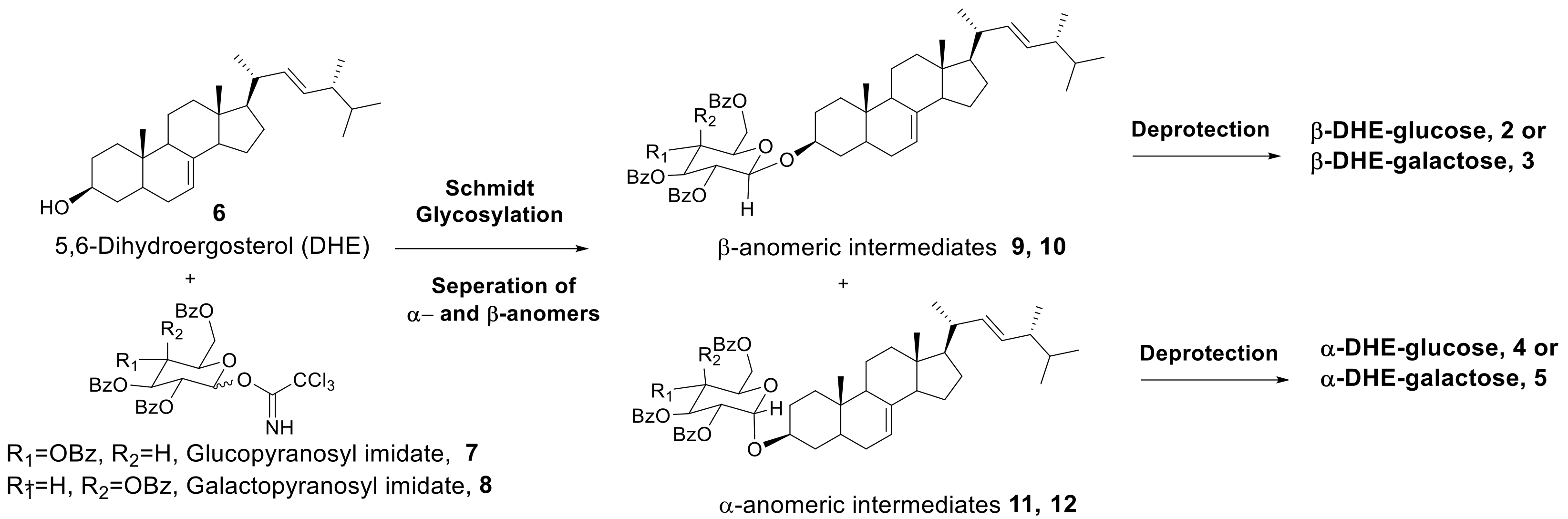
As shown in Scheme 1, β- and α-anomers (2–5) of DHE-glycosides can be prepared by acid-catalyzed Schmidt glycosylation of DHE with OH-protected sugars followed by deprotection.
Previously, we determined which of various glycosyl donors, including glycosyl halide, glycosyl sulfide, and glycosyl trichloroacetimidate, in the presence of several catalysts, resulted in the most efficient glycosylation of DHE. We found that when O-tetrabenzoyl-protected glycosyl trichloroacetimidates (7 and 8) were used as glycosyl donors and Cu(OTf)2 was used as a catalyst, an effective stereoselective glycosylation reaction occurred, especially for β-anomers [6]. Therefore, glycosyl trichloroacetimidates (7 or 8), glycosyl donors for Cu(OTf)2-catalyzed glycosylation, were used in this study. The two glycosyl donors, glucopyranosyl and galactopyranosyl trichloroacetimidate, and DHE were synthesized according to previous methods [6].
The anomeric effect in sugar chemistry predicts that α-anomers will be the major product of glycosylation reactions. However, β-anomers have been reported to be produced when acyl groups capable of neighboring group participation (NGP) are present at adjacent carbon positions. Unlike β-anomers, which are the major products in our reaction with a benzoyl group in an adjacent position, α-anomers are not readily available in sufficient quantities to carry out biological activity studies. According to the literature, α-anomers are primarily produced when a benzyl (Bn)-protected sugar is used as a glycosyl donor [19]. We attempted to glycosylate with a Bn-protected sugar, but there was an issue during the debenzylation step. Therefore, we examined the various reaction conditions under which α-anomers are obtained with a benzoyl (Bz)-protected sugar. Experimental results for reactions of glucosyl imidate (7) with DHE (6) under various conditions are shown in Table 1. As expected, β-anomers were primarily synthesized in the four acid catalytic conditions (Entry 1–4), and α-anomers were obtained at a 10% yield under the SiO2-H2SO4 acid condition (Entry 2).
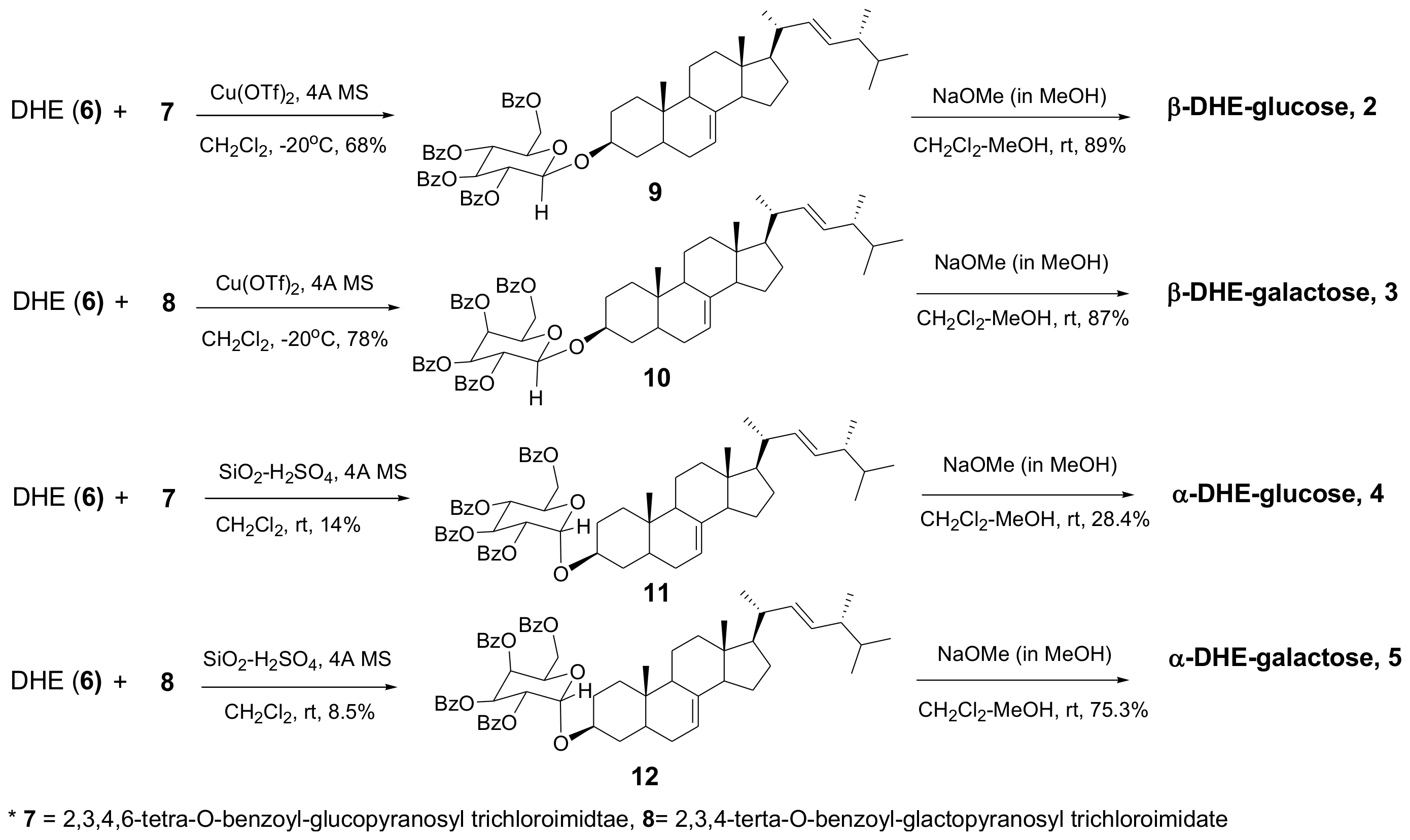
The reaction was monitored by thin layer chromatography (TLC) and we confirmed that both α- and β-anomers were produced during the early stages of the reaction (the major product was β-anomers), and that α-anomers disappeared as reaction time elapsed. Therefore, an SiO2-H2SO4-catalyzed glycosylation reaction was done to obtain the benzoyl group-protected α-anomers, α-DHE-Glc (4) and α-DHE-Gal (5). The yields of the two α-anomers were 14% and 8.5%, respectively. OH-protected glucosyl DHE (9 and 11) and OH-protected galactosyl DHE (10 and 12) were hydrolyzed using NaOMe/MeOH to yield α- and β-anomers of DHE-Glc (2 and 4) and DHE-Gal (3 and 5), respectively (Scheme 2).
2.2. Effect of α- and β-Anomers of 5,6-Dihydroergosterol(DHE)-Glycosides on Cell Viability
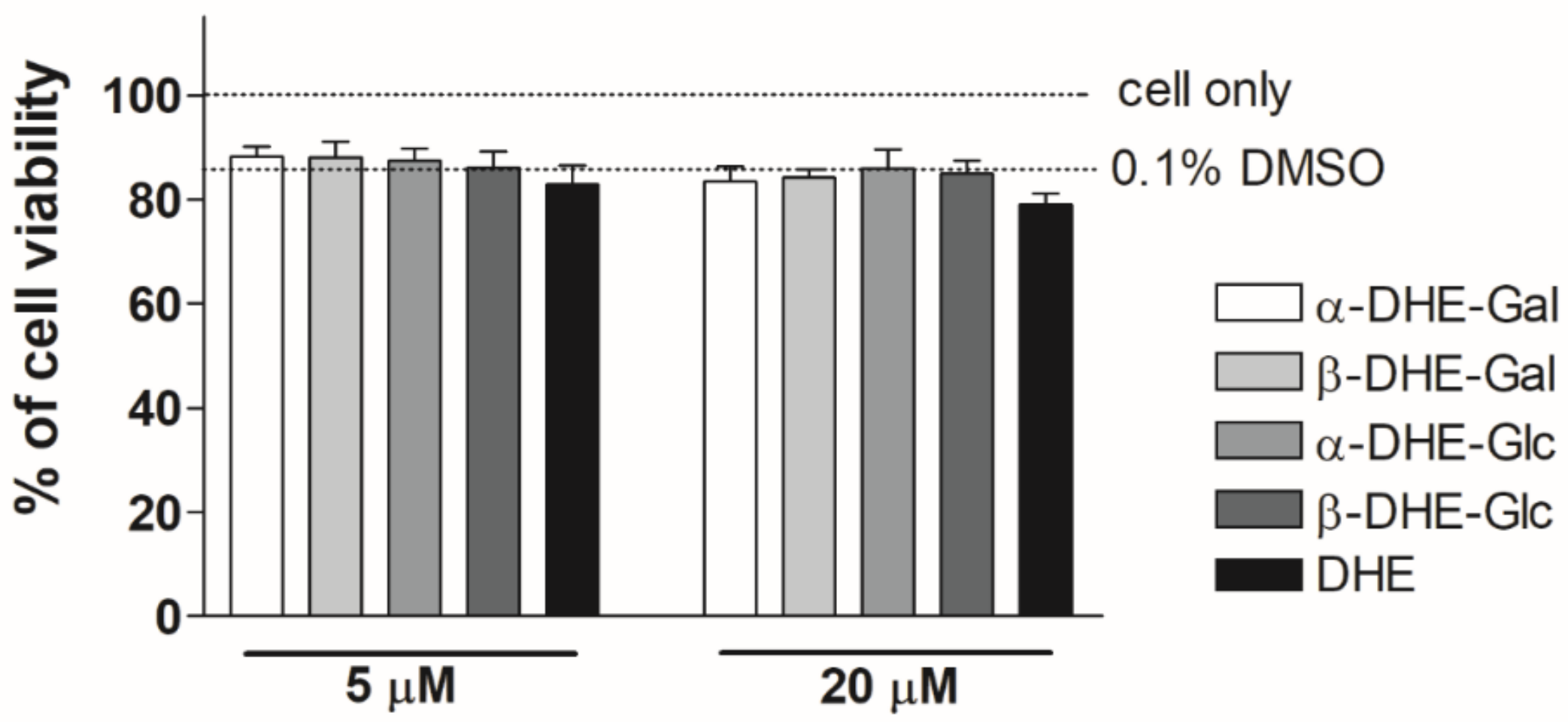
The viability of HaCaT cells treated with α- and β-anomers was determined with MTT assays to determine experimental DHE-Glc and DHE-Gal concentrations. We have previously confirmed the effects of β-anomer DHE-sugar derivatives on cell viability [15]. As shown in Figure 2, none of the tested α- and β-anomers of DHE-Glc and DHE-Gal had an effect on cell viability up to a concentration of 20 μM. In addition, we confirmed that cell survival remained at about 80% when the concentration of the compounds was 20 μM, and cell viability at these concentrations was similar to that obtained with a 0.1% DMSO working solution alone. Therefore, all experiments were performed using concentrations up to 20 μM, and results can be interpreted independently of cell survival.
2.3. Effects of DHE-Glycoside α- and β-Anomers on mRNA Expression Levels of Pro-Inflammatory Cytokines and Chemokines in TNF-α/IFN-γ Induced HaCaT Cells
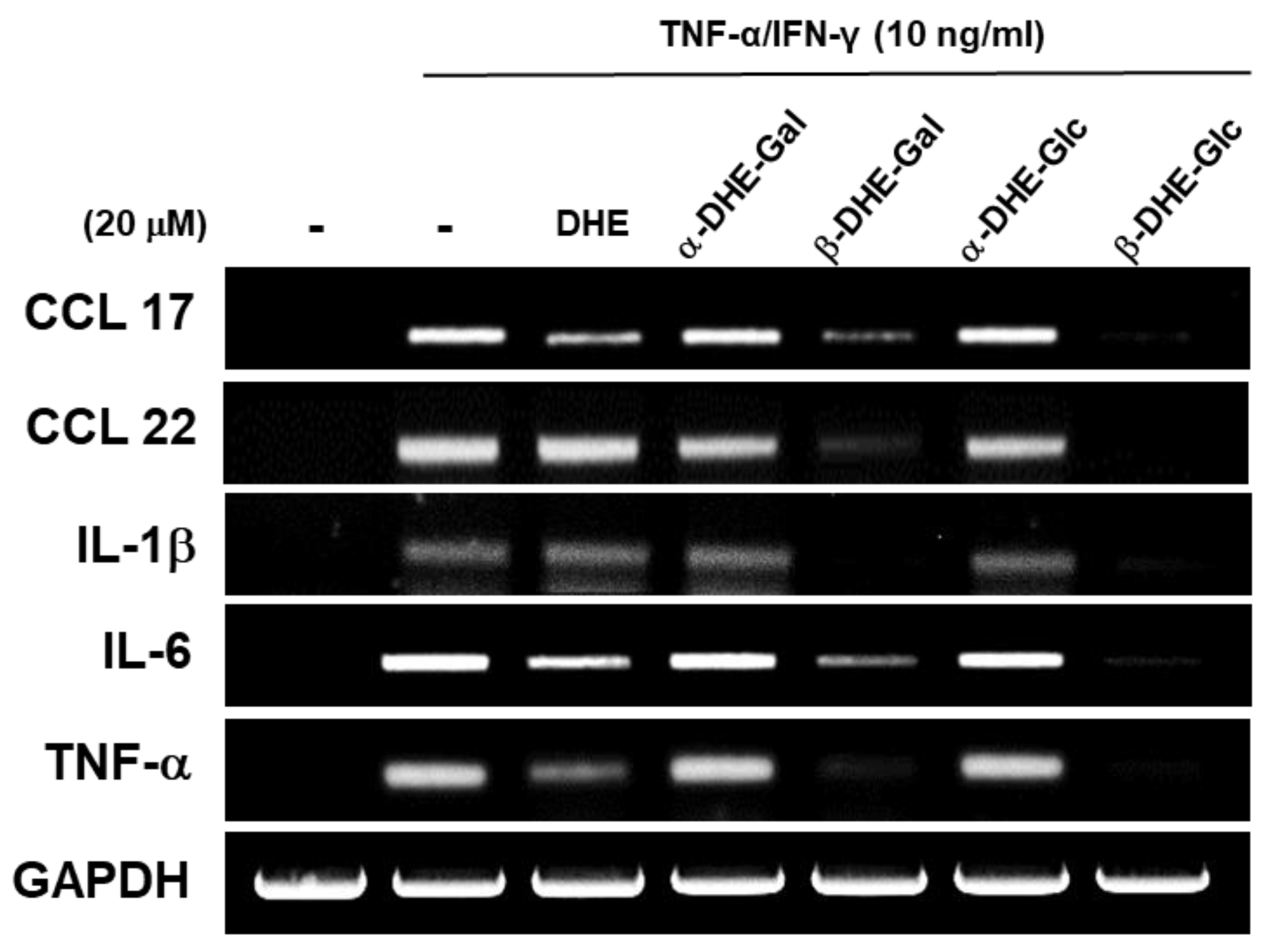
We examined the effects of the α- and β-anomers of DHE-Glc and DHE-Gal on pro-inflammatory cytokine and chemokine gene expression in HaCaT cells. We evaluated the mRNA expression levels of pro-inflammatory cytokines TNF-α, IL-1β, and IL-6 using an RT-PCR assay in TNF-α/IFN-γ-treated HaCaT cells. In addition, we also measured mRNA expression levels of chemokines CCL17 and CCL22, which are known to have a role in chronic skin inflammation. We confirmed that mRNA expression levels of the pro-inflammatory cytokines TNF-α, IL-6, and IL-1β were dramatically increased in TNF-α/IFN-γ-treated HaCaT cells. CCL17 and CCL22 mRNA expression levels had a similar expression pattern to TNF-α, IL-6, and IL-1β. Neither DHE (6), nor the DHE-glycoside α-anomers (3 and 5), had an effect on TNF-α, IL-6, or IL-1β mRNA levels in TNF-α/IFN-γ-treated HaCaT cells. In contrast, we found that DHE-glycoside β-anomers (2 and 3) reduced pro-inflammatory cytokine and chemokine mRNA expression in activated HaCaT cells (Figure 3). These results indicate that the β-anomers of DHE-glycosides were more effective than the α-anomers of DHE-glycosides at inhibiting gene expression of pro-inflammatory cytokines including IL1-β, IL-6, TNF-α and chemokines CCL17 and CCL22 in TNF-α/IFN-γ-treated HaCaT cells.
2.4. Effects of DHE-Glycoside α- and β-Anomers on Pro-Inflammatory Cytokine and Chemokine Protein Expression Levels in TNF-α/IFN-γ-Induced HaCaT Cells
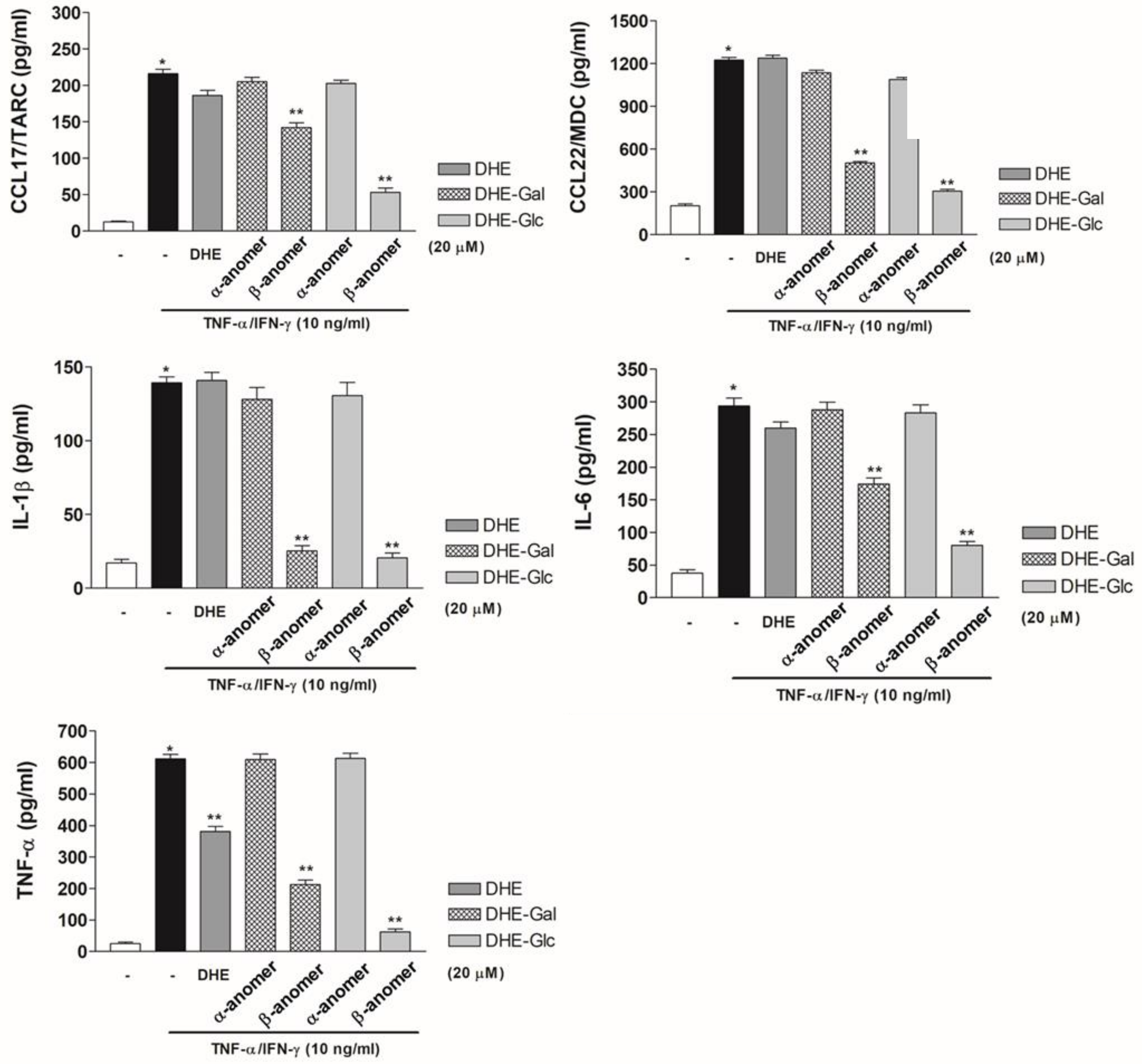
Next, we used ELISA assays to examine whether DHE-glycoside α- and β-anomers have an effect on pro-inflammatory cytokine and chemokine protein expression in TNF-α/IFN-γ–induced HaCaT cells. In TNF-α/IFN-γ–induced HaCaT cells, TNF-α, IL-6, IL-1β, CCL17, and CCL22 protein expression increased by approximately 4- to 9-fold compared to controls. DHE treatment had no effect on pro-inflammatory cytokine protein expression in HaCaT cells relative to control cells. Moreover, TNF-α protein expression was slightly reduced compared to the positive control. HaCaT cells treated with α-anomers of DHE-Glc and DHE-Gal (4 and 5) had similar levels of IL-1β, IL-6, TNF-α, CCL17, and CCL22 protein expression as DHE-treated cells. These results were consistent with the mRNA gene expression results. However, TNF-α/IFN-γ-induced HaCaT cells treated with β-anomers of DHE-Glc and DHE-Gal (2 and 3) showed reduced pro-inflammatory cytokine and chemokine protein expression. We confirmed that the β-anomer of DHE-Glc was more effective than the β-anomer of DHE-Gal at reducing cytokine and chemokine protein expression (Figure 4). Our results indicate that the β-anomer of DHE-Glc (2) inhibited TNF-α/IFN-γ-activated expression of IL-1β, IL-6, TNF-α, CCL17, and CCL22 proteins, which resulted in the reduction of cytokine and chemokine mRNA levels in HaCaT cells.
3. Discussion
The genesis of this study was evaluating the anti-inflammatory activity of extract of the leaves of Stewartia koreana (SKE), which is a natural herb native to Korea. We found that SKE had potent anti-inflammatory activity. SKE inhibited the LPS-induced NF-κB signaling mechanism in RAW264.7 mouse macrophages, and strongly inhibited the production of NO and PGE2 [20]. Next, we investigated the active ingredient of SKE, and identified spinasterol-Glc (1) as the main active ingredient of SKE extract. Spinasterol-Glc (1) inhibited the production of LPS-induced NO in RAW264.7 mouse macrophage cells, in addition to inhibiting the expression of the pro-inflammatory cytokines TNF-α, IL-6, and IL-1β. This inhibition proved to be the result of inhibition of the IκB-α/IKK phosphorylation process [21]. We also found that spinasterol-Glc (1) strongly inhibited the expression of TARC/CCL17 stimulated by TNF-α/IFN-γ in HaCaT cells. This phenomenon was caused by suppression of phosphorylation of c-Raf, p38 MAPK, and JAK2. Furthermore, spinasterol-Glc (1) inhibited NF-κB and STAT1 promoter activation, and we confirmed that spinasterol-Glc (1) was a potential treatment option for chronic skin inflammation due to its ability to suppress CCL17 expression [3]. However, spinasterol-Glc (1) is difficult to isolate from natural extracts or synthesize. Therefore, we designed and synthesized an analogue of spinasterol, 5,6-dihydroergosterol (DHE). DHE has the same steroidal backbone as spinasterol, but different side chains. We have examined the biological activity of four DHE-glycosides, including ergosterol, in previous studies. We showed that DHE-glycosides strongly inhibited the production of NO induced by LPS in RAW264.7 mouse macrophages [6]. Next, we investigated the inhibitory effect of DHE-Glc on chronic skin inflammation in DNCB-induced animal models. We found that DHE-Glc inhibited the infiltration of epidermal eosinophil and mast cells in our DNCB-induced skin inflammation animal model and that DHE-Glc reduced the concentration of IgE and histamine and mRNA expression and protein levels of CCL17/22 in the plasma of DNCB-treated animals. We also confirmed that inhibition of CCL17/22 expression was due to suppression of NF-κB and STAT1 signaling [7]. Therefore, we reasoned that DHE-Glc (2) would be useful as a therapeutic agent for chronic skin inflammation.
In our synthesis process, however, the α- and β-anomers of DHE-glycosides have always been obtained together, particularly during the glycosylation step. To determine the relative biological activity of α- and β-anomers, we prepared α- and β-anomers of DHE-glycosides and evaluated their anti-inflammatory activity. In more detail, we synthesized α- and β-anomers of DHE-Glc and DHE-Gal (2, 3, 4, and 5) via acid-catalyzed glycosylation of DHE (6) with OH-protected glucosyl trichloroimidate (7) or OH-protected galactosyl trichloroimidate (8), followed by deprotection (Scheme 2). Based on the results of glycosylation reactions in various acids (Table 1), β-anomers were synthesized by Cu(OTf)2-catalyzed glycosylation followed by deprotection, and α-anomers were obtained by SiO2-H2SO4-catalyzed glycosylation followed by deprotection. We determined the anti-chronic inflammatory activities of these compounds by measuring mRNA and protein expression levels of pro-inflammatory cytokines TNF-α, IL-6, and IL-1β and chemokines CCL17 and CCL22 in TNF-α/IFN-γ-stimulated HaCaT cells after treatment of these cells with these compounds. We found that the β-anomers of DHE glycosides had a greater inhibitory effect than the α-anomers of DHE-glycosides on the gene expression of the pro-inflammatory cytokines IL1-β, IL-6, TNF-α and the chemokines CCL17 and CCL22 in HaCaT cells. In addition, the β-anomers of DHE-glycosides decreased protein expression levels of these pro-inflammatory cytokines and chemokines. This inhibitory effect is likely due to inhibition of the phosphorylation of IKK/IκBα and activation of NF-κB and STAT1 by the β-anomers of DHE-glycosides [7]. In chronic skin inflammatory sites, keratinocytes and various immune cells are activated by inflammatory stimuli, such as TNF-α and IFN-γ [22,23]. The secretion of chemokines and pro-inflammatory cytokines from keratinocytes is responsible for chronic skin disorder diseases as well as abnormal immune responses [24,25,26]. TNF-α and IFN-γ induce strong inflammation in vivo [26,27]. According to previous reports, TNF-α and IFN-γ increased the expression and secretion of pro-inflammatory cytokines IL-1β and IL-6 and chemokines CCL17 and CCL22 in chronic skin inflammatory lesions [10,28,29,30]. Therefore, inhibiting the secretion of pro-inflammatory cytokines and chemokines is important for developing drug candidates that can effectively treat chronic skin inflammation.
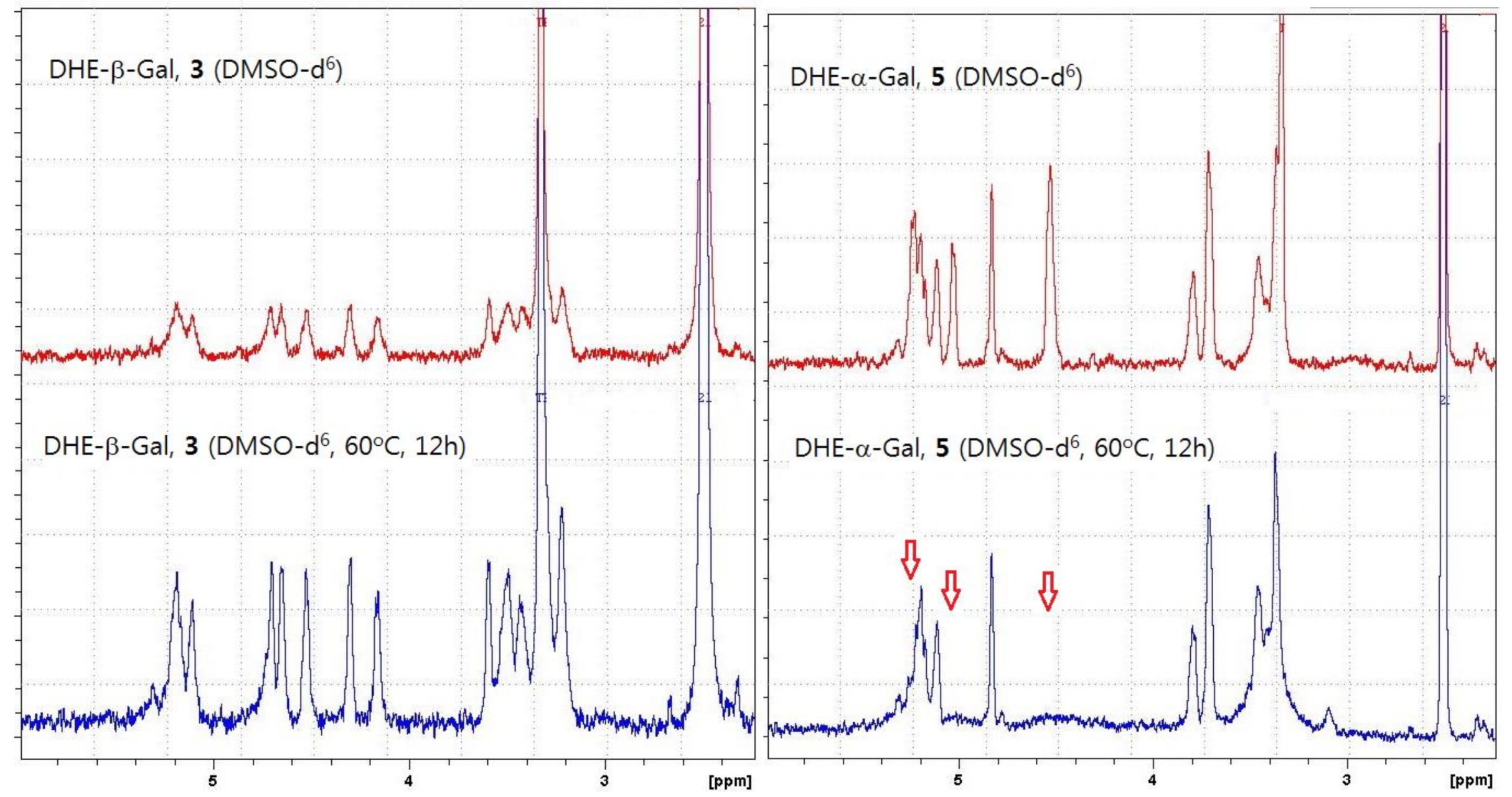
We found that the β-anomers of DHE-glycosides had a greater anti-inflammatory effect than the α-anomers of DHE-glycosides. We hypothesized that the higher physiological activity of β-anomers might be explained by their greater stability in solution than the α-anomers, because some studies have reported better stability of β-anomeric glycosides than α-anomeric glycosides [31,32]. Consistent with our hypothesis, we found that the purified α-anomers of DHE-glycosides were relatively unstable in solution or in weakly acidic conditions, like silica gel, based on both thin layer chromatography (TLC) and NMR experiments. TLC experiments showed that α-anomers were not completely stable in silica gel plates, and as a result, α-anomers decomposed during silica gel chromatography purification to give low yields. In the NMR experiment, α- and β-anomers of DHE-Gal (5 and 3) were dissolved in DMSO-d6 and kept for 12 h at 60 °C, after which NMR spectra were obtained. The spectrum of DHE-β-Gal (3) shows that it was stable, while the spectrum of DHE-α-Gal (5) revealed a change in the NMR peaks from 4.5 to 5.5 ppm (Figure 5). These results confirmed that DHE-α-Gal (5) was unstable and that its structure changed. Based on these results, we suggest that the anti-inflammatory activity of β-anomers of DHE-glycosides is higher than that of the α-anomers because of the poor stability of the α-anomers of DHE-glycosides.
In conclusion, we designed and synthesized α- and β-anomers (2, 3, 4, and 5) of 5,6-dihydroergosterol (DHE)-glycosides as anti-inflammatory agents and evaluated their relative anti-inflammatory activity. β-Anomers of DHE-Glc (2) and DHE-Gal (3) were prepared by Cu(OTf)2-catalyzed glycosylation followed by deprotection, while α-anomers of DHE-Glc (4) and DHE-Gal (5) were obtained by SiO2-H2SO4-catalyzed glycosylation followed by deprotection. We found that β-anomers of DHE-glycosides (2 and 3) reduced pro-inflammatory cytokine and chemokine mRNA expression in TNF-α/IFN-γ-activated HaCaT cells, and that the β-anomer of DHE-Glc (2) inhibited TNF-α/IFN-γ-activated expression of IL-1β, IL-6, TNF-α, CCL17, and CCL22 proteins, which resulted in the reduction of cytokine and chemokine mRNA levels in HaCaT cells. These results indicate that the β-anomer of DHE-Glc (2) had an inhibitory effect on inflammatory mediator expression in TNF-α/IFN-γ-stimulated HaCaT cells. Based on these results, we concluded that the β-anomers of DHE-Glc (2) and DHE-Gal (3) exhibited potent anti-inflammatory effects. Furthermore, the β-anomers of DHE-glycosides had a greater anti-inflammatory effect than the α-anomers of DHE-glycosides. We found that the purified α-anomers of DHE-glycosides were unstable in solution or in slightly acidic conditions based on both TLC and NMR experiments. This could explain why the anti-inflammatory activities of α-anomers of DHE-glycosides were lower than those of the β-anomers of DHE-glycosides. Further analyses are needed to determine the chemical stability of the α-anomers of DHE-glycosides. The structure-activity relationship (SAR) of various DHE-glycosides will be discussed in detail at a later date.
4. Materials and Methods
4.1. Chemicals and Instruments
1H NMR and 13C NMR spectra were recorded on a Jeol 300 MHz spectrometer and Jeol 75 MHz spectrometer (Peabody, MA, USA). Chemical shifts (δ) are reported in parts-per-million (ppm); coupling constants (J) are reported in Hertz (Hz). The following abbreviations were used to explain multiplicities: s = singlet, d = doublet, t = triplet, q = quartet, m = multiplet, and br s = broad singlet. Melting points were determined on a Barnstead Electrothermal 9100 instrument. HR-EI+-MS was recorded on a Jeol JMS-700 instrument. All chemicals and reagents were purchased from Sigma-Aldrich Co. Ltd. (St. Louis, MO, USA). All solvents used in reactions were purchased from Honeywell Burdick & Jackson® (Muskegon, MI, USA). Reaction progress was monitored by thin layer chromatography (TLC, Merck kiesegel 60F254, Kenilworth, NJ, USA), and column chromatography was performed using a Merck silica gel 60 (230–400 mesh). 5,6-Dihydroergosterol (DHE) (6), 2,3,4,6-tetra-O-benzoyl-glucopyranosyl trichloroacetimidate (OH-protected glucosyl imidate) (7), and 2,3,4,6-tetra-O-benzoyl-galactopyranosyl trichloroacetimidate (OH-protected galactosyl imidate) (8) were prepared as reported previously [6,33,34].
2,3,4,6-Tetrabenzoyl-β-glucopyranosyl dihydroergosterol (9)
2,3,4,6-Tetra-O-benzoyl-glucopyranosyl trichloroacetimidate (7) (1.21 g, 1.63 mmol), 5,6-dihydroergosterol (8) (0.50 g, 1.25 mmol), and a 4Å molecular sieve in methylene chloride (27.0 mL) were stirred at −20 °C for 0.5 h. After addition of Cu(OTf)2 (49.0 mg, 0.14 mmol), the mixture was stirred at −20 °C overnight. After neutralization by Et3N, the mixture was filtered through celite. The filtrate was concentrated in vacuo and purified by flash column chromatography (ethyl acetate/hexane) to produce a white solid.
Yield: 68%. m.p. 160–163 °C. 1H-NMR (300 MHz, CDCl3) δ 0.51–2.10 (m, 42H, peaks from steroidal structure), 3.51–3.66 (m, 1H), 4.14–4.20 (m, 1H), 4.52 (dd, 1H, J = 5.9, 12.1 Hz), 4.61 (dd, 1H, J = 3.3, 11.9 Hz), 4.95 (d, 1H, J = 7.9 Hz), 5.03–5.13 (m, 1H), 5.18 (t, 2H, J = 6.3 Hz), 5.50 (dd, 1H, J = 7.9, 9.7 Hz), 5.63 (t, 1H, J = 9.7 Hz), 5.90 (t, 1H, J = 9.7 Hz), 7.29–7.59 (m, 12H), 7.82–8.03 (m, 8H).
(3β,22E)-Ergosta-7,22-dien-3-yl-β-d-glucopyranoside (DHE-β-Glc) (2)
2,3,4,6-Tetrabenzoyl-β-glucopyranosyl dihydroergosterol (9) (0.70 g, 0.72 mmol) in a mixed solvent (CH2Cl2:MeOH = 7.0 mL:7.0 mL) was added to NaOMe (0.5 M in MeOH, 8.6 mL, 4.30 mmol). The mixture was stirred at room temperature overnight. After neutralization by Dowex Mac-3, the mixture was filtered. The filtrate was concentrated in vacuo and purified by flash column chromatography (CH2Cl2/MeOH) to produce a white solid.
Yield: 89%. m.p. 267–270 °C. 1H-NMR (300 MHz, Pyridine-d5) δ 0.59 (s, 3H), 0.74 (s, 3H), 0.89 (d, 6H, J = 6.8 Hz), 0.99 (d, 4H, J = 6.8 Hz), 1.08 (d, 3H, J = 6.4 Hz), 1.15–1.32 (m, 4H), 1.33–1.62 (m, 8H), 1.64–1.76 (m, 4H), 1.76–1.86 (m, 2H), 1.86–1.96 (m, 2H), 1.99–2.06 (m, 3H), 3.92–4.11 (m, 3H), 4.24–4.34 (m, 2H), 4.41–4.46 (m, 1H), 4.60 (d, 1H, J = 11.5 Hz), 4.87 (brs, −OH peaks), 5.04 (d, 1H, J = 7.7 Hz), 5.19 (brs, 1H), 5.26–5.29 (m, 1H). 13C-NMR (75 MHz, Pyridine-d5) δ 12.48, 13.26, 18.05, 20.04, 20.35, 21.58, 21.96, 23.48, 28.75, 30.24, 33.56, 34.77, 34.97, 37.56, 39.85, 40.41, 41.08, 43.31, 43.72, 49.84, 55.55, 56.34, 63.19, 72.05, 75.67, 77.36, 78.83, 78.96, 80.09, 102.60, 118.23, 132.46, 136.58, 139.95. HR-EI+-MS m/z: 560.4075 (Calcd for C34H56O6: 560.4077).
2,3,4,6-Tetrabenzoyl-β-galactopyranosyl dihydroergosterol (10)
2,3,4,6-Tetra-O-benzoyl-galactopyranosyl trichloroacetimidate (8) (2.23 g, 3.01 mmol), 5,6-dihydroergosterol (6) (1.0 g, 2.51 mmol), and a 4Å molecular sieve in methylene chloride (36.0 mL) were stirred at −20 °C for 0.5 h. After addition of Cu(OTf)2 (90 mg, 0.25 mmol), the mixture was stirred at −20 °C for 4 h. After neutralization by Et3N, the mixture was filtered through celite. The filtrate was concentrated in vacuo and purified by flash column chromatography (ethyl acetate/hexane) to produce a white solid.
Yield: 74%. m.p. 158–161 °C. 1H-NMR (300 MHz, CDCl3) δ 0.50–2.35 (42H, peaks from steroidal structure), 3.56–3.75 (m, 1H), 4.27–4.38 (m, 1H), 4.40–4.47 (m, 1H), 4.66–4.72 (m, 1H), 4.93 (d, 1H, J = 7.7 Hz), 5.10–5.25 (m, 2H), 5.60 (dd, 1H, J = 3.2, 10.4 Hz), 5.75–5.81 (m, 1H), 5.99 (d, 1H, J = 2.8 Hz), 7.21–7.26 (m, 2H), 7.35–7.63 (m, 10H), 7.80 (d, 2H, J = 7.5 Hz), 7.96 (d, 2H, J = 7.5 Hz), 8.03 (d, 2H, J = 8.1 Hz), 8.11 (d, 2H, J = 7.3 Hz).
(3β,22E)-Ergosta-7,22-dien-3-yl-β-d-galactopyranoside (DHE-β-Gal) (3)
2,3,4,6-Tetrabenzoyl-β-galactopyranosyl dihydroergosterol (10) (0.20 g, 0.20 mmol) in a mixed solvent (CH2Cl2:MeOH = 2.0 mL:2.0 mL) was added to NaOMe (0.5 M in MeOH, 2.5 mL, 1.23 mmol). The mixture was stirred at room temperature for 5 h. After neutralization by Dowex Mac-3, the mixture was filtered. The filtrate was concentrated in vacuo and purified by flash column chromatography (CH2Cl2-MeOH) to produce a white solid.
Yield: 87%. m.p. 267–269 °C. 1H-NMR (300 MHz, Pyridine-d5) δ 0.59 (s, 3H), 0.73 (s, 3H), 0.80–0.85 (m, 1 H), 0.89 (dd, 6H, J = 1.9, 6.7 Hz), 0.99 (d, 4H, J = 6.8 Hz), 1.08 (d, 3H, J = 6.4 Hz), 1.18–1.62 (m, 13H), 1.68–1.72 (m, 4H), 1.77–1.86 (m, 1H), 1.88–1.94 (m, 1H), 1.98–2.17 (m, 3H), 3.92–4.07 (m, 1 H), 4.14 (t, 1H, J = 5.9 Hz), 4.21–4.29 (m, 1 H), 4.42–4.56 (m, 3H), 4.59–4.66 (m, 1 H), 4.97 (d, 2H, J = 7.5 Hz), 5.19 (brs, 1H), 5.27 (t, 1H, J = 5.8 Hz). 13C-NMR (75 MHz, Pyridine-d5) δ 12.46, 13.24, 18.05, 20.03, 20.35, 21.58, 21.94, 23.48, 28.74, 30.24, 30.27, 33.55, 34.73, 34.74, 34.97, 37.57, 39.85, 40.41, 41.07, 43.30, 43.70, 49.84, 55.54, 56.34, 62.85, 70.63, 72.99, 75.72, 77.24, 103.18, 118.22, 132.45, 136.57, 139.93. HR-EI+-MS m/z: 560.4075 (Calcd for C34H56O6: 560.4077).
2,3,4,6-Tetrabenzoyl-β-glucopyranosyl dihydroergosterol (9)
2,3,4,6-Tetra-O-benzoyl-glucopyranosyl trichloroacetimidate (7) (1.86 g, 2.51 mmol), DHE (6) (0.50 g, 1.25 mmol), and a 4Å molecular sieve in methylene chloride (31.0 mL) were stirred at room temperature for 0.5 h. After the addition of SiO2-H2SO4 (4.67 mmol/g, 27 mg, 0.125 mmol), the mixture was stirred at room temperature for 2.5 h. After neutralization by Et3N, the mixture was filtered. The filtrate was concentrated in vacuo and purified by flash column chromatography (ethyl acetate/hexane) to produce a white solid.
Yield: 14% (α-anomer). 1H-NMR (300 MHz, CDCl3) δ 0.50–2.06 (m, 42H, peaks from steroidal backbone), 3.26–3.40 (m, 1H), 4.05–4.13 (m, 1H), 4.35 (dd, 1H, J = 4.7, 12.1 Hz), 4.50 (dd, 1H, J = 2.8, 12.1 Hz), 4.74–4.84 (m, 1H), 5.09 (brs, 1 H), 5.17 (t, 2H, J = 6.1 Hz), 5.49 (d, 1H, J = 8.8 Hz), 5.77 (d, 1H, J = 1.8 Hz), 6.04 (d, J = 5.1 Hz, 1H), 7.19–7.29 (m, 2H), 7.36–7.52 (m, 8H), 7.55–7.66 (m, 2H), 7.75–7.83 (m, 2H), 7.88–8.00 (m, 4H), 8.05–8.13 (m, 2H).13C-NMR (75 MHz, CDCl3) δ 12.05, 12.88, 17.63, 19.66, 19.98, 21.12, 21.44, 22.73, 22.92, 29.74, 33.12, 34.04, 35.62, 37.16, 39.45, 40.28, 40.52, 42.87, 43.29, 49.37, 55.10, 56.00, 64.08, 67.50, 68.61, 69.25, 72.12, 73.60, 97.65, 117.52, 121.67, 126.62, 128.35, 128.40, 128.64, 128.76, 129.25, 129.35, 129.72, 129.83, 129.87, 130.11, 130.25, 132.04, 133.17, 133.70, 133.85, 135.88, 136.44, 139.52, 164.83, 165.44, 166.24 HR-FAB+-MS m/z: 977.5209 (M + 1) (Calcd for C62H72O10: 976.51).
(3β,22E)-Ergosta-7,22-dien-3-yl-α-d-glucopyranoside (DHE-α-Glc) (4)
2,3,4,6-Tetrabenzoyl-α-glucopyranosyl dihydroergosterol (11) (0.10 g, 0.10 mmol) in a mixed solvent (CH2Cl2:MeOH = 1.0 mL:1.0 mL) was added to NaOMe (0.5 M in MeOH, 1.0 mL, 0.51 mmol). The mixture was stirred at room temperature overnight. After neutralization by Dowex Mac-3, the mixture was filtered. The filtrate was concentrated in vacuo and purified by flash column chromatography (CH2Cl2/MeOH) to produce a white solid.
Yield = 28.4% (unstable). 1H-NMR (300 MHz, Pyridine-d5) 0.51–2.10 (42H, peaks from steroidal backbone), 4.30–4.53 (m, 4H), 4.59–4.71 (m, 1H), 5.00–5.20 (m, 3H), 5.22–5.31 (m, 2H), 6.49–6.60 (m, 1H). 13C NMR (75 MHz, Pyridine-d5) δ 12.30, 13.06, 17.90, 19.89, 20.21, 21.26, 21.42, 21.73, 23.32, 30.01, 33.42, 34.27, 37.38, 38.52, 40.46, 41.51, 43.16, 43.55, 49.61, 49.78, 55.37, 56.15, 62.88, 70.36, 73.18, 74.95, 76.03, 79.81, 99.49, 117.95, 121.06, 132.20, 139.75. HR-FAB−-MS m/z: 559.3994 (M − 1) (Calcd for C34H56O6: 560.4077).
2,3,4,6-Tetrabenzoyl-α-galactopyranosyl dihydroergosterol (12)
2,3,4,6-Tetra-O-benzoyl-galactopyranosyl trichloroacetimidate (8) (2.23 g, 3.01 mmol), DHE (6) (1.0 g, 2.51 mmol), and a 4Å molecular sieve in methylene chloride (63.0 mL) were stirred at room temperature for 0.5 h. After the addition of SiO2-H2SO4 (4.67 mmol/g, 54 mg, 0.25 mmol), the mixture was stirred at room temperature overnight. After neutralization by Et3N, the mixture was filtered through celite. The filtrate was concentrated in vacuo and purified by flash column chromatography (ethyl acetate/hexane) to produce a white solid.
Yield: 8.5% (α-anomer). 1H-NMR (300 MHz, CDCl3) δ 0.54–2.06 (42H, peaks from steroidal backbone), 3.54–3.69 (m, 1H), 4.62–4.69 (m, 1H), 4.71–4.80 (m, 2H), 5.09–5.26 (m, 3H), 5.45 (d, 2H, J = 12.1 Hz), 5.61 (d, 1H, J = 5.3 Hz), 5.97–6.10 (m, 1H), 7.25–7.33 (m, 4H), 7.34–7.47 (m, 4H), 7.48–7.61 (m, 4H), 7.92 (d, 2H, J = 8.3 Hz), 7.99 (d, 2H, J = 8.3 Hz), 8.07 (t, 4H, J = 8.2 Hz). 13C-NMR (75 MHz, CDCl3) δ 12.09, 12.99, 17.62, 19.66, 19.99, 21.14, 21.48, 22.94, 28.16, 29.40, 29.70, 33.13, 33.97, 34.35, 37.05, 39.48, 40.14, 40.57, 42.85, 43.34, 49.38, 55.15, 56.00, 63.44, 70.27, 81.97, 82.76, 104.02, 117.59, 128.55, 128.59, 129.22, 129.33, 129.68, 129.77, 129.93, 130.02, 130.16, 132.05, 133.30, 133.42, 133.52, 133.63, 135.88, 139.70, 165.77, 165.97, 166.33. HR-FAB+-MS m/z: 977.5209 (M + 1) (Calcd for C62H72O10: 976.51).
(3β,22E)-Ergosta-7,22-dien-3-yl-α-d-galactopyranoside (DHE-α-Gal) (5)
2,3,4,6-Tetrabenzoyl-α-galactopyranosyl dihydroergosterol (12) (0.17 g, 0.17 mmol) in a mixed solvent (CH2Cl2:MeOH = 1.8 mL:1.8 mL) was added to NaOMe (0.5 M in MeOH, 1.05 mL, 0.51 mmol). The mixture was stirred at room temperature overnight. After neutralization by Dowex Mac-3, the mixture was filtered. The filtrate was concentrated in vacuo and purified by flash column chromatography (CH2Cl2/MeOH) to produce a white solid.
Yield= 75.3% (unstable). 1H-NMR (300 MHz, Pyridine-d5) δ 0.58–2.17 (42H, peaks from steroidal backbone), 2.70–3.00 (m, 4H, −OH peaks), 3.71–3.87 (m, 1H), 4.39–4.47 (m, 2H), 4.55–4.65 (m, 1H), 4.88–4.97 (m, 2H), 5.10 (dd, 1H, J = 4.7, 6.9 Hz,), 5.21 (brs, 1H), 5.28 (t, 2H, J = 6.0 Hz), 5.70–5.75 (m, 1H). 13C-NMR (75 MHz, Pyridine-d5) δ 12.47, 13.26, 18.04, 20.04, 20.35, 21.58, 21.95, 23.49, 28.75, 30.24, 33.56, 34.75, 35.01, 37.54, 39.85, 40.55, 41.08, 43.31, 43.72, 46.13, 49.85, 55.55, 56.34, 65.12, 73.16, 76.66, 79.08, 84.21, 107.94, 118.21, 132.46, 136.59, 139.99. HR-FAB+-MS m/z: 561.4161 (M + 1) (Calcd for C34H56O6: 560.4077).
4.2. Cell Culture
Human keratinocytes (HaCaT) were cultured in Dulbecco’s modified Eagles medium (Welgene, Seoul, Korea), containing 10% fetal bovine serum (FBS), 100 units/mL penicillin, and 100 μg/mL streptomycin (Gibco BRL, Rockville, MD, USA). Cell culture conditions were as follows: 95% humidity and 5% (v/v) mixture of air and CO2, in a cell culture incubator at 37 °C.
4.3. Cell Viability Assay
Cultured cells were removed from cell culture plates, and HaCaT cells at a concentration of 5 × 104 cells/well were re-cultured in 96-well plates for 18 h. After the growth medium was discarded, HaCaT cells were treated with the indicated concentration of each compound in serum-free medium for 24 h. After removing the culture medium, cells were treated with 100 μg/mL of 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-thetazolium bromide (MTT) for 1 h for formazan formation in live cells. Crystallized formazan was dissolved in 200 μL DMSO and the wavelength of each sample was analyzed by spectrophotometer using an absorbance measurement at 560 nm. Analyses were repeated three times, and results are expressed as means of three independent experiments.
4.4. Reverse Transcriptase-Polymerase Chain Reaction (RT-PCR) Analysis
The RNA-Bee isolation kit (Tel-Test, Friendswood, TX, USA) was used according to the manufacturer’s recommendations for isolating total RNA from HaCaT cells. cDNA was prepared using reverse-transcriptase M-MuLV (Fermentas Life Science, Pittsburgh, PA, USA). PCR amplification was performed using specific primers (Bioneer, Daejeon, Korea).
4.5. Enzyme-Linked Immunosorbent Assay
HaCaT cells were cultured in 6-well plates and then the growth medium was removed. Cells were treated with TNF-α/IFN-γ (10 ng/mL) in the presence or absence of each compound for 18 h. Supernatants of cultured cells were collected and analyzed using an enzyme-linked immunosorbent assay (ELISA) kit (R&D Systems, Minneapolis, MN, USA) according to the manufacturer’s instructions.
4.6. Statistical Analysis
Unless otherwise stated, all experiments were performed with triplicate samples and repeated at least three times. Data are presented as means ± S.D. and statistical comparisons between groups were performed using 1-way ANOVA followed by a Student’s t-test.
Author Contributions
H.K. and T.H.L. designed the study; T.K.K., Y.K.C., and H.P. performed the chemical syntheses; T.H.L. and H.K. analyzed the data and wrote the manuscript.
Funding
This research was funded by the GRRC Program of Gyeonggi province [GRRC-KyungHee2017(B01)], Republic of Korea.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Fiore, C.; Eisenhut, M.; Ragazzi, E.; Zanchin, G.; Armanini, D. A history of the therapeutic use of liquorice in Europe. J. Ethnopharmacol. 2005, 99, 317–324. [Google Scholar] [CrossRef] [PubMed]
- Sparg, S.G.; Light, M.E.; van Staden, J. Biological activities and distribution of plant saponins. J. Ethnopharmacol. 2004, 94, 219–243. [Google Scholar] [CrossRef] [PubMed]
- Jung, M.R.; Lee, T.H.; Bang, M.H.; Kim, H.; Son, Y.; Chung, D.K.; Kim, J. Suppression of thymus- and activation-regulated chemokine (TARC/CCL17) production by 3-O-β-d-glucopyanosylspinasterol via blocking NF-κB and STAT1 signaling pathways in TNF-α and IFN-γ-induced HaCaT keratinocytes. Biochem. Biophys. Res. Commun. 2012, 427, 236–241. [Google Scholar] [CrossRef] [PubMed]
- Giroux, S.; Corey, E.J. Stereocontrolled synthesis of dafachronic acid A, the ligand for the DAF-12 nuclear receptor of Caenorhabditis elegans. J. Am. Chem. Soc. 2007, 129, 9866–9867. [Google Scholar] [CrossRef] [PubMed]
- Badreddine, A.; Mostafa, K.E.; Zarrouk, A.; Nury, T.; Kharassi, Y.E.; Nasser, B.; Malki, M.C.; Lizard, G.; Samadi, M. An expeditious synthesis of spinasterol and schottenol, two phytosterols present in argan oil and in cactus pear seed oil, and evaluation of their biological activities on cells of the central nervous system. Steroids 2015, 99, 119–124. [Google Scholar] [CrossRef] [PubMed]
- Park, H.G.; Lee, T.H.; Chang, F.; Kwon, H.J.; Kim, J.Y.; Kim, H.W. Synthesis of ergosterol and 5,6-dihydroergosterol glycosides and their inhibitory activities on lipopolysaccharide-induced nitric oxide production. Bull. Korean Chem. Soc. 2013, 34, 1339–1344. [Google Scholar] [CrossRef]
- Jung, M.; Lee, T.H.; Oh, H.J.; Kim, H.; Son, Y.; Lee, E.H.; Kim, J. Inhibitory effect of 5,6-dihydroergosteol-glucoside on atopic dermatitis-like skin lesions via suppression of NF-κB and STAT activation. J. Dermatol. Sci. 2015, 79, 252–261. [Google Scholar] [CrossRef]
- Kasraie, S.; Werfel, T. Role of macrophages in the pathogenesis of atopic dermatitis. Mediators Inflamm. 2013, 2013. [Google Scholar] [CrossRef]
- Yamanaka, K.; Mizutani, H. The role of cytokines/chemokines in the pathogenesis of atopic dermatitis. Curr. Probl. Dermatol. 2011, 41, 80–92. [Google Scholar]
- Caproni, M.; Torchia, D.; Cardinali, C.; Volpi, W.; Del Bianco, E.; D’Agata, A.; Fabbri, P. Infiltrating cells, related cytokines and chemokine receptors in lesional skin of patients with dermatomyositis. Br. J. Dermatol. 2004, 151, 784–791. [Google Scholar] [CrossRef]
- Mu, Z.; Zhao, Y.; Chang, C.; Zhang, J. Molecular biology of atopic dermatitis. Clin. Rev. Allergy Immunol. 2014, 47, 193–218. [Google Scholar] [CrossRef] [PubMed]
- Soumelis, V.; Reche, P.A.; Kanzler, H.; Yuan, W.; Edward, G.; Homey, B.; Gilliet, M.; Ho, S.; Antonenko, S.; Lauerma, A.; et al. Human epithelial cells trigger dendritic cell mediated allergic inflammation by producing TSLP. Nat. Immunol. 2002, 3, 673–680. [Google Scholar] [CrossRef] [PubMed]
- Albanesi, C.; Scarponi, C.; Sebastiani, S.; Cavani, A.; Federici, M.; Sozzani, S.; Girolomoni, G. A cytokine-to-chemokine axis between T lymphocytes and keratinocytes can favor Th1 cell accumulation in chronic inflammatory skin diseases. J. Leukoc. Biol. 2001, 70, 617–623. [Google Scholar] [PubMed]
- Scheu, S.; Ali, S.; Ruland, C.; Arolt, V.; Alferink, J. The C-C Chemokines CCL17 and CCL22 and Their Receptor CCR4 in CNS Autoimmunity. Int. J. Mol. Sci. 2017, 18, 2306. [Google Scholar] [CrossRef] [PubMed]
- Osabe, M.; Tajika, T.; Tohkin, M. Allopurinol suppresses expression of the regulatory T-cell migration factors TARC/CCL17 and MDC/CCL22 in HaCaT keratinocytes via restriction of nuclear factor-κB activation. J. Appl. Toxicol. 2018, 38, 274–283. [Google Scholar] [CrossRef] [PubMed]
- Miyano, K.; Matsushita, S.; Tsuchida, T.; Nakamura, K. Inhibitory effect of a histamine 4 receptor antagonist on CCL17 and CCL22 production by monocyte-derived Langerhans cells in patients with atopic dermatitis. J. Dermatol. 2016, 43, 1024–1029. [Google Scholar] [CrossRef] [PubMed]
- Kwon, Y.S.; Oh, S.H.; Wu, W.H.; Bae, B.G.; Lee, H.J.; Lee, M.G.; Lee, K.H. CC chemokines as potential immunologic markers correlated with clinical improvement of atopic dermatitis patients by immunotherapy. Exp. Dermatol. 2010, 19, 246–251. [Google Scholar] [CrossRef] [PubMed]
- Narbutt, J.; Lesiak, A.; Sysa-Jedrzeiowska, A.; Zakrzewski, M.; Bogaczewicz, J.; Stelmach, I.; Kuna, P. The imbalance in serum concentration of Th-1- and Th-2-derived chemokines as one of the factors involved in pathogenesis of atopic dermatitis. Mediators Inflamm. 2009, 2009. [Google Scholar] [CrossRef] [PubMed]
- Wuts, P.G.M.; Green, T.W. Protective Group in Organic Synthesis, 4th ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2006; pp. 102–120. [Google Scholar]
- Lee, T.H.; Kwak, H.B.; Kim, H.H.; Lee, Z.H.; Chung, D.K.; Baek, N.I.; Kim, J. Methanol extracts of Stewartia koreana inhibit cyclooxygenase-2 (COX-2) and inducible nitric oxide synthase (iNOS) gene expression by blocking NF-kappaB transactivation in LPS-activated RAW264.7 cells. Mol. Cells 2007, 23, 398–404. [Google Scholar]
- Lee, T.H.; Jung, M.; Bang, M.H.; Chung, D.K.; Kim, J. Inhibitory effects of a spinasterol glycoside on lipopolysaccharide-induced production of nitric oxide and proinflammatory cytokines via down-regulating MAP kinase pathways and NF-kB activation in RAW264.7 macrophage cells. Int. Immunopharmacol. 2012, 13, 264–270. [Google Scholar] [CrossRef]
- Lee, K.S.; Chun, S.Y.; Lee, M.G.; Kim, S.; Jang, T.J.; Nam, K.S. The prevention of TNF-α/IFN-γ mixture-induced inflammation in human keratinocyte and atopic dermatitis-like skin lesions in Nc/Nga mice by mineral-balanced deep sea water. Biomed. Pharmacother. 2018, 97, 1331–1340. [Google Scholar] [CrossRef] [PubMed]
- Mazanet, M.M.; Neote, K.; Hughes, C.C. Expression of IFN-inducible T cell alpha chemoattractant by human endothelial cells is cyclosporin A-resistant and promotes T cell adhesion: Implications for cyclosporin A-resistant immune inflammation. J. Immunol. 2000, 164, 5383–5388. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, N.; Sugaya, M.; Suga, H.; Oka, T.; Kawaguchi, M.; Miyagaki, T.; Fujita, H.; Sato, S. Thymic Stromal Chemokine TSLP Acts through Th2 Cytokine Production to Induce Cutaneous T-cell Lymphoma. Cancer Res. 2016, 76, 6241–6252. [Google Scholar] [CrossRef] [PubMed]
- Mollanazar, N.K.; Smith, P.K.; Yosipovitch, G. Mediators of Chronic Pruritus in Atopic Dermatitis: Getting the Itch Out? Clin. Rev. Allergy Immunol. 2016, 51, 263–292. [Google Scholar] [CrossRef] [PubMed]
- Rebane, A.; Zimmermann, M.; Aab, A.; Baurecht, H.; Koreck, A.; Karelson, M.; Abram, K.; Metsalu, T.; Pihlap, M.; Meyer, N.; et al. Mechanisms of IFN-γ-induced apoptosis of human skin keratinocytes in patients with atopic dermatitis. J. Allergy Clin. Immunol. 2012, 129, 1297–1306. [Google Scholar] [CrossRef] [PubMed]
- Byrne, A.M.; Goleva, E.; Chouiali, F.; Kaplan, M.H.; Hamid, Q.A.; Leung, D.Y. Induction of GITRL expression in human keratinocytes by Th2 cytokines and TNF-α: Implications for atopic dermatitis. Clin. Exp. Allergy 2012, 42, 550–559. [Google Scholar] [CrossRef] [PubMed]
- Giustizieri, M.L.; Mascia, F.; Frezzolini, A.; De Pità, O.; Chinni, L.M.; Giannetti, A.; Girolomoni, G.; Pastore, S. Keratinocytes from patients with atopic dermatitis and psoriasis show a distinct chemokine production profile in response to T cell-derived cytokines. J. Allergy Clin. Immunol. 2001, 107, 871–877. [Google Scholar] [CrossRef] [PubMed]
- Park, J.H.; Kim, M.S.; Jeong, G.S.; Yoon, J. Xanthii fructus extract inhibits TNF-α/IFN-γ-induced Th2-chemokines production via blockade of NF-κB, STAT1 and p38-MAPK activation in human epidermal keratinocytes. J. Ethnopharmacol. 2015, 171, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Classen, A.; Kalali, B.N.; Schnopp, C.; Andres, C.; Aguilar-Pimentel, J.A.; Ring, J.; Ollert, M.; Mempel, M. TNF receptor I on human keratinocytes is a binding partner for staphylococcal protein A resulting in the activation of NF kappa B, AP-1, and downstream gene transcription. Exp. Dermatol. 2011, 20, 48–52. [Google Scholar] [CrossRef]
- Usabiaga, I.; González, J.; León, I.; Arnaiz, P.F.; Cocinero, E.J.; Ferńandez, J.A. Influence of the anomeric conformation in the intermolecular interactions of glucose. J. Phys. Chem. Lett. 2017, 8, 1147–1151. [Google Scholar] [CrossRef]
- Davis, A.P.; Wareham, R.S. Carbohydrate Recognition through Noncovalent Interactions: A Challenge for Biomimetic and Supramolecular Chemistry. Angew. Chem. Int. Ed. 1999, 38, 2978–2996. [Google Scholar] [CrossRef]
- Mbadugha, B.N.S.; Menger, F.M. Sugar/Steroid/Sugar conjugates: Sensitivity of lipid binding to sugar structure. Org. Lett. 2003, 5, 4041–4044. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.Y.; Yao, Q.; Liu, Y.; Si, C.L.; Chen, J.; Jiu, N.; Lian, H.Y.; Ding, L.N.; Zhang, Y.M. Highly efficient synthesis and antitumor activity of monosaccharide saponins mimicking components of Chinese folk medicine Cordyceps sinensis. J. Asian Nat. Pro. Res. 2012, 14, 429–435. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the α-and β-anomers of DHE-glycosides are available from the authors upon request. |
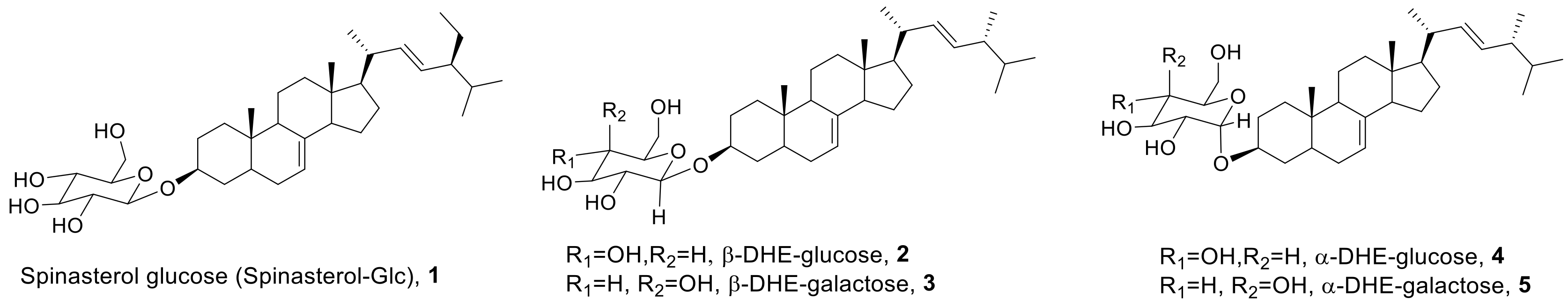
Figure 1.
Structures of spinasterol-glucose (spinasterol-Glc) and α- and β-anomers of 5,6-dihydroergosterol (DHE)-glycosides.
Figure 1.
Structures of spinasterol-glucose (spinasterol-Glc) and α- and β-anomers of 5,6-dihydroergosterol (DHE)-glycosides.

Scheme 1.
Synthesis of α- and β-anomers of DHE-glycosides by Schmidt glycosylation.

Scheme 2.
Synthesis of α- and β-anomers of DHE-Glc and DHE-Gal.

Figure 2.
Effects of α- and β-anomers of DHE-Glc (2 and 4) and DHE-Gal (3 and 5) on HaCaT cell viability. HaCaT cells were treated with synthetic α- and β-anomers of DHE-Glc and DHE-Gal, and cell viability was evaluated using an MTT assay. Cells were stabilized and then incubated for 24 h at the indicated concentrations. Results are shown as a percentage of the negative control treated with the medium alone. Each value represents the mean ± S.D.
Figure 2.
Effects of α- and β-anomers of DHE-Glc (2 and 4) and DHE-Gal (3 and 5) on HaCaT cell viability. HaCaT cells were treated with synthetic α- and β-anomers of DHE-Glc and DHE-Gal, and cell viability was evaluated using an MTT assay. Cells were stabilized and then incubated for 24 h at the indicated concentrations. Results are shown as a percentage of the negative control treated with the medium alone. Each value represents the mean ± S.D.

Figure 3.
Effects of α- and β-anomers of DHE-Glc and DHE-Gal (2, 3, 4, and 5) on pro-inflammatory cytokine and chemokine gene expression. To determine the effects of synthetic α- and β-anomers of DHE-Glc and DHE-Gal on pro-inflammatory cytokine IL-1b, IL-6, and TNF-α expression, mRNA levels were determined using an RT-PCR assay. mRNA expression levels of the chemokines CCL17 and CCL22 were compared under similar conditions. RT-PCR reactions were performed in triplicate and repeated at least three times.
Figure 3.
Effects of α- and β-anomers of DHE-Glc and DHE-Gal (2, 3, 4, and 5) on pro-inflammatory cytokine and chemokine gene expression. To determine the effects of synthetic α- and β-anomers of DHE-Glc and DHE-Gal on pro-inflammatory cytokine IL-1b, IL-6, and TNF-α expression, mRNA levels were determined using an RT-PCR assay. mRNA expression levels of the chemokines CCL17 and CCL22 were compared under similar conditions. RT-PCR reactions were performed in triplicate and repeated at least three times.

Figure 4.
Effects of DHE-glycoside α- and β-anomers on pro-inflammatory cytokine and chemokine protein expression. To determine the effects of synthetic DHE-glycoside α- and β-anomers on IL-1β, IL-6, and TNF-α protein expression, protein expression was determined using ELISA assays. CCL17 and CCL22 protein expression was determined under same conditions. The values for protein expression intensity are the mean ± S.D. of three independent experiments (n = 3). * p < 0.01 compared to mock medium alone, ** p < 0.01 compared to treatment with TNF-α/IFN-γ.
Figure 4.
Effects of DHE-glycoside α- and β-anomers on pro-inflammatory cytokine and chemokine protein expression. To determine the effects of synthetic DHE-glycoside α- and β-anomers on IL-1β, IL-6, and TNF-α protein expression, protein expression was determined using ELISA assays. CCL17 and CCL22 protein expression was determined under same conditions. The values for protein expression intensity are the mean ± S.D. of three independent experiments (n = 3). * p < 0.01 compared to mock medium alone, ** p < 0.01 compared to treatment with TNF-α/IFN-γ.

Figure 5.
NMR spectra of α- and β-anomers of DHE-Gal (3 and 5) in DMSO-d6 at room temperature and 60 °C.
Figure 5.
NMR spectra of α- and β-anomers of DHE-Gal (3 and 5) in DMSO-d6 at room temperature and 60 °C.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Glycosylation of glucosyl imidate (7) with DHE (6) under various conditions a.

| Entry | Acid Catalyst a | Temp. | Time (h) | Yield (%) b | |
|---|---|---|---|---|---|
| α-anomer, 11 | β-anomer, 9 | ||||
| 1 | Cellulose-HClO4 | R.T. | 48 | Trace | 58% |
| 2 | SiO2-H2SO4 | R.T. | 24 | 10% | 55% |
| 3 | TMSOTf | −20 °C | 24 | Trace | 77% |
| 4 | Cu(OTf)2 | −20 °C | 24 | Trace | 68% |
a DHE (1.0 equiv), imidate (1.3 equiv), and 4Å MS in CH2Cl2 were stirred for 0.5 h. After addition of an acid catalyst (10 mol%), the mixture was stirred at room temperature or −20 °C. b Isolated yields.
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kim, T.K.; Cho, Y.K.; Park, H.; Lee, T.H.; Kim, H. Comparison of the Inhibitory Activities of 5,6-Dihydroergosterol Glycoside α- and β-Anomers on Skin Inflammation. Molecules 2019, 24, 371. https://doi.org/10.3390/molecules24020371
AMA Style
Kim TK, Cho YK, Park H, Lee TH, Kim H. Comparison of the Inhibitory Activities of 5,6-Dihydroergosterol Glycoside α- and β-Anomers on Skin Inflammation. Molecules. 2019; 24(2):371. https://doi.org/10.3390/molecules24020371
Chicago/Turabian StyleKim, Tae Kyun, Young Kyoung Cho, HoonGyu Park, Tae Hoon Lee, and Hakwon Kim. 2019. "Comparison of the Inhibitory Activities of 5,6-Dihydroergosterol Glycoside α- and β-Anomers on Skin Inflammation" Molecules 24, no. 2: 371. https://doi.org/10.3390/molecules24020371