A Novel Synthetic Steroid of 2β,3α,5α-Trihydroxy-androst-6-one Alleviates the Loss of Rat Retinal Ganglion Cells Caused by Acute Intraocular Hypertension via Inhibiting the Inflammatory Activation of Microglia


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
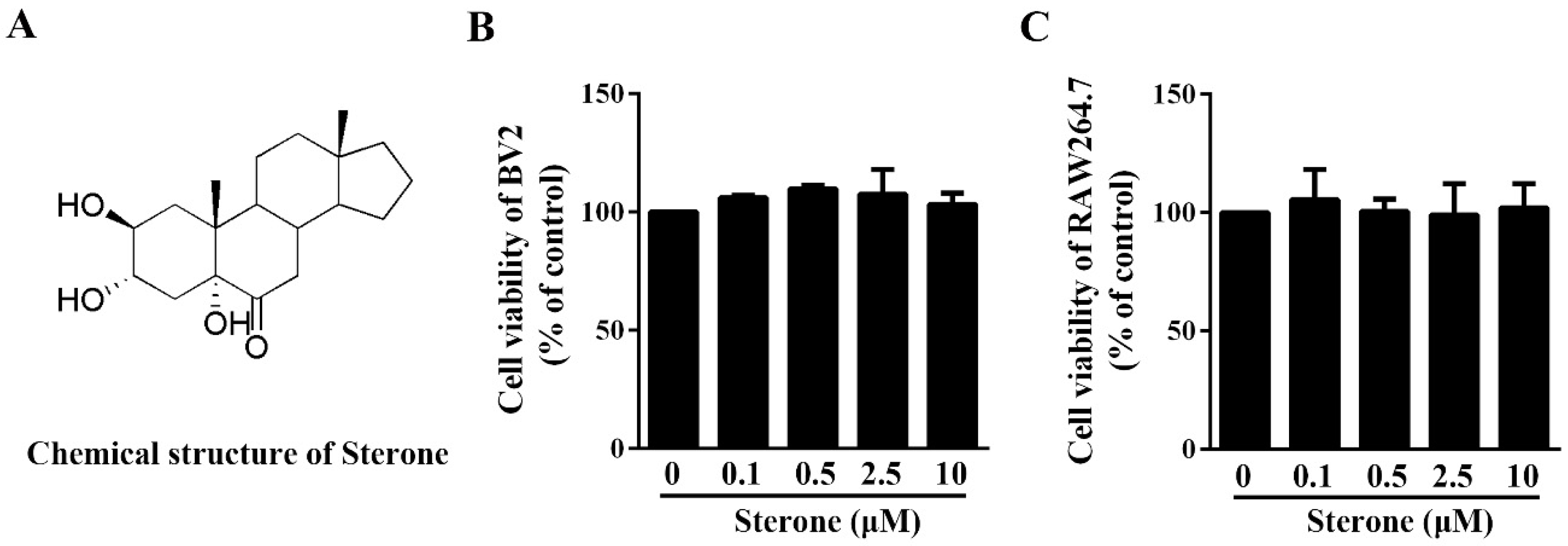
2.1. Sterone Has No Effect on the Viability of BV2 Cells and RAW264.7 Cells
2.2. Sterone Inhibits LPS-Induced Inflammatory Activation of BV2 and RAW264.7 Cells.
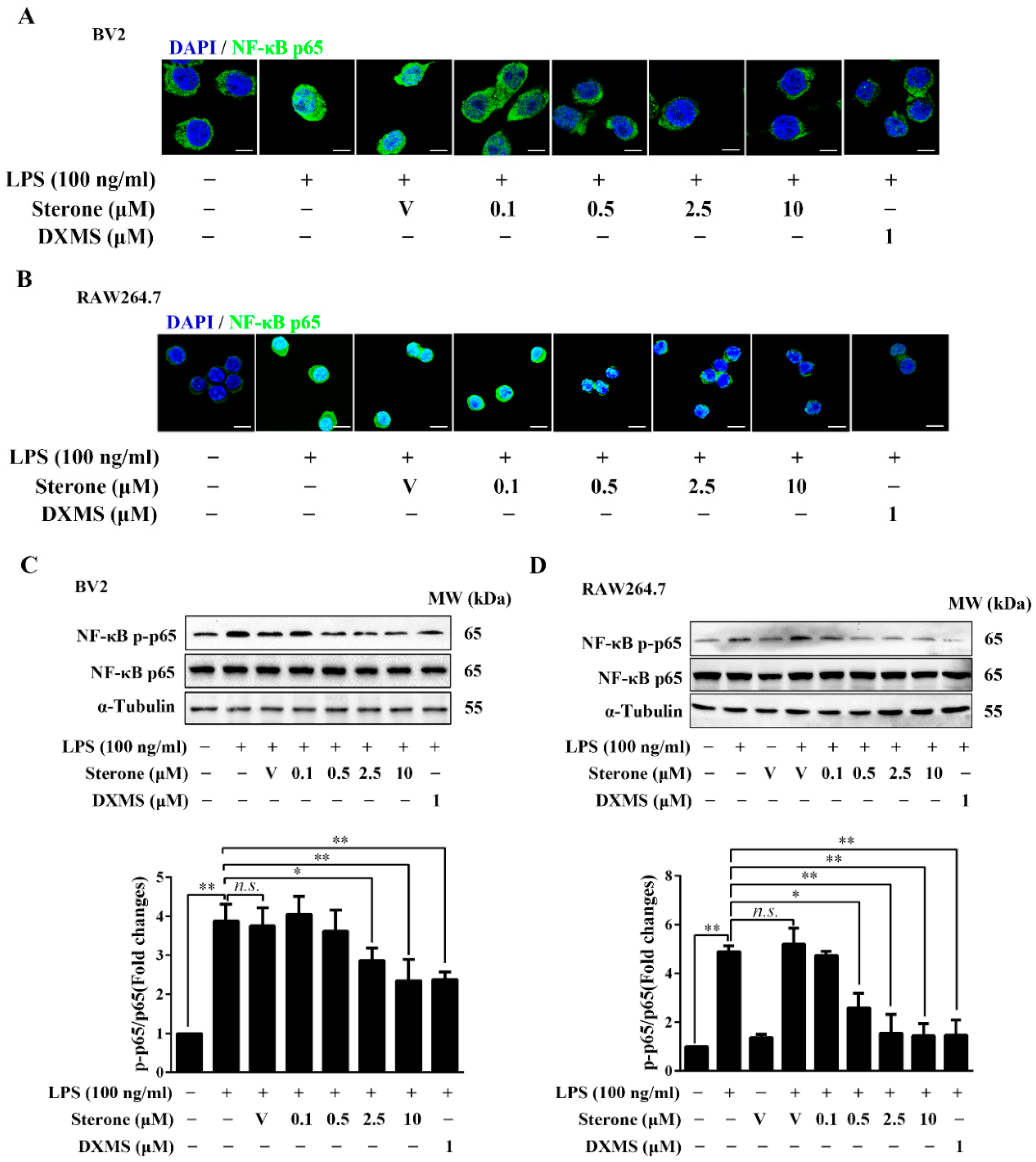
2.3. Sterone Inhibits LPS-Induced NF-κB Activation in BV2 and RAW264.7 Cells.
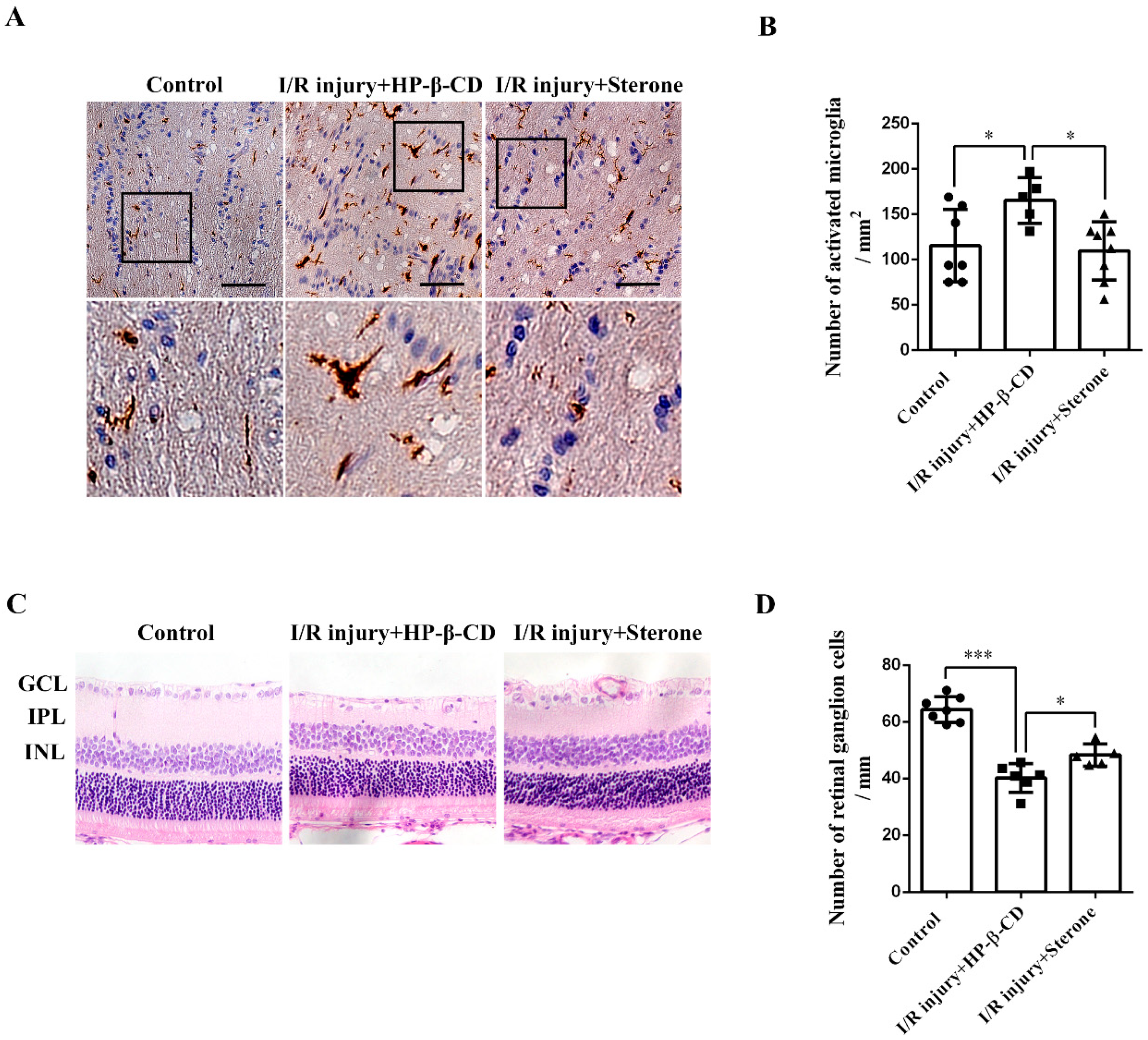
2.4. Sterone Inhibits Microglial Activation and Retinal RGCs Loss Induced via Ischemia/Reperfusion Injury in a Rat AIH Model
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Treatment
4.2. Cell Viability Analysis
4.3. Western Blotting Analysis
4.4. Quantitative Real-Time PCR (RT-qPCR) Analysis
4.5. Immunofluorescence Analysis
4.6. Retinal Ischemia/Reperfusion of AIH and Treatment
4.7. Immunohistochemistry Analysis and Hematoxylin Eosin (H&E) Staining
4.8. Statistical Analysis
5. Conclusions
6. Patents
Author Contributions
Funding
Conflicts of Interest
References
- Almasieh, M.; Wilson, A.M.; Morquette, B.; Cueva Vargas, J.L.; Di Polo, A. The molecular basis of retinal ganglion cell death in glaucoma. Progr. Retinal Eye Res. 2012, 31, 152–181. [Google Scholar] [CrossRef] [PubMed]
- Schehlein, E.M.; Novack, G.D.; Robin, A.L. New classes of glaucoma medications. Curr. Opin. Ophthalmol. 2017, 28, 161–168. [Google Scholar] [CrossRef]
- Heijl, A.; Leske, M.C.; Bengtsson, B.; Hyman, L.; Bengtsson, B.; Hussein, M. Reduction of intraocular pressure and glaucoma progression: Results from the Early Manifest Glaucoma Trial. Arch. Ophthalmol. 2002, 120, 1268–1279. [Google Scholar] [CrossRef] [PubMed]
- Leske, M.C.; Heijl, A.; Hyman, L.; Bengtsson, B.; Komaroff, E. Factors for progression and glaucoma treatment: The Early Manifest Glaucoma Trial. Curr. Opin. Ophthalmol. 2004, 15, 102–106. [Google Scholar] [CrossRef]
- Vohra, R.; Tsai, J.C.; Kolko, M. The role of inflammation in the pathogenesis of glaucoma. Survey Ophthalmol. 2013, 58, 311–320. [Google Scholar] [CrossRef] [PubMed]
- Williams, P.A.; Marsh-Armstrong, N.; Howell, G.R. Neuroinflammation in glaucoma: A new opportunity. Exp. Eye Res. 2017, 157, 20–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wan, P.; Su, W.; Zhang, Y.; Li, Z.; Deng, C.; Zhuo, Y. Trimetazidine protects retinal ganglion cells from acute glaucoma via the Nrf2/Ho-1 pathway. Clin. Sci. 2017, 131, 2363–2375. [Google Scholar] [CrossRef] [Green Version]
- Su, W.; Li, Z.; Jia, Y.; Zhuo, Y. Rapamycin is neuroprotective in a rat chronic hypertensive glaucoma model. PLoS ONE 2014, 9, e99719. [Google Scholar] [CrossRef]
- Soto, I.; Howell, G.R. The complex role of neuroinflammation in glaucoma. Cold Spring Harbor Perspect. Med. 2014, 4. [Google Scholar] [CrossRef]
- Xiong, X.Y.; Liu, L.; Yang, Q.W. Functions and Mechanisms of Microglia/Macrophages in Neuroinflammation and Neurogenesis during Stroke. Prog. Neurobiol. 2016, 142, 23–44. [Google Scholar] [CrossRef]
- Bi, W.; Zhu, L.; Jing, X.; Zeng, Z.; Liang, Y.; Xu, A.; Liu, J.; Xiao, S.; Yang, L.; Shi, Q.; et al. Rifampicin improves neuronal apoptosis in LPS-stimulated cocultured BV2 cells through inhibition of the TLR-4 pathway. Mol. Med. Rep. 2014, 10, 1793–1799. [Google Scholar] [CrossRef] [PubMed]
- Freedman, J.; Iserovich, P. Pro-inflammatory cytokines in glaucomatous aqueous and encysted Molteno implant blebs and their relationship to pressure. Investig. Ophthalmol. Visual Sci. 2013, 54, 4851–4855. [Google Scholar] [CrossRef] [PubMed]
- Luna, J.D.; Chan, C.C.; Derevjanik, N.L.; Mahlow, J.; Chiu, C.; Peng, B.; Tobe, T.; Campochiaro, P.A.; Vinores, S.A. Blood-retinal barrier (BRB) breakdown in experimental autoimmune uveoretinitis: Comparison with vascular endothelial growth factor, tumor necrosis factor alpha, and interleukin-1beta-mediated breakdown. J. Neurosci. Res. 1997, 49, 268–280. [Google Scholar] [CrossRef]
- Ramirez, A.I.; de Hoz, R.; Salobrar-Garcia, E.; Salazar, J.J.; Rojas, B.; Ajoy, D.; Lopez-Cuenca, I.; Rojas, P.; Trivino, A.; Ramirez, J.M. The Role of Microglia in Retinal Neurodegeneration: Alzheimer’s Disease, Parkinson, and Glaucoma. Front. Aging Neurosci. 2017, 9, 214. [Google Scholar] [CrossRef] [PubMed]
- Nakazawa, T.; Nakazawa, C.; Matsubara, A.; Noda, K.; Hisatomi, T.; She, H.; Michaud, N.; Hafezi-Moghadam, A.; Miller, J.W.; Benowitz, L.I. Tumor necrosis factor-alpha mediates oligodendrocyte death and delayed retinal ganglion cell loss in a mouse model of glaucoma. J. Neurosci. 2006, 26, 12633–12641. [Google Scholar] [CrossRef] [PubMed]
- Minchevatasheva, S.; Soler, R.M. NF-κB signaling pathways: Role in nervous system physiology and pathology. Neurosci. Rev. J. Bringing Neurobiol. Neurol. Psychiatry 2013, 19, 175–194. [Google Scholar]
- Shih, R.H.; Wang, C.Y.; Yang, C.M. NF-κB Signaling Pathways in Neurological Inflammation: A Mini Review. Front. Mol. Neurosci. 2015, 8, 77. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Tang, E.; Guan, K.; Wang, C.Y. IKK beta plays an essential role in the phosphorylation of RelA/p65 on serine 536 induced by lipopolysaccharide. J. Immunol. 2003, 170, 5630–5635. [Google Scholar] [CrossRef]
- Huang, C.; Wang, J.; Lu, X.; Hu, W.; Wu, F.; Jiang, B.; Ling, Y.; Yang, R.; Zhang, W. Z-guggulsterone negatively controls microglia-mediated neuroinflammation via blocking IκB-α-NF-κB signals. Neurosci. Lett. 2016, 619, 34–42. [Google Scholar] [CrossRef]
- Su, L.; Zhang, R.; Chen, Y.; Zhu, Z.; Ma, C. Raf Kinase Inhibitor Protein Attenuates Ischemic-Induced Microglia Cell Apoptosis and Activation Through NF-κB Pathway. Cell. Physiol. Biochem. 2017, 41, 1125–1134. [Google Scholar] [CrossRef] [Green Version]
- Shi, S.; Liang, D.; Chen, Y.; Xie, Y.; Wang, Y.; Wang, L.; Wang, Z.; Qiao, Z. Gx-50 reduces β-amyloid-induced TNF-α, IL-1β, NO, and PGE2 expression and inhibits NF-κB signaling in a mouse model of Alzheimer’s disease. Eur. J. Immunol. 2016, 46, 665–676. [Google Scholar] [CrossRef] [PubMed]
- Sappington, R.M.; Calkins, D.J. Contribution of TRPV1 to microglia-derived IL-6 and NFkappaB translocation with elevated hydrostatic pressure. Investig. Ophthalmol. Visual Sci. 2008, 49, 3004–3017. [Google Scholar] [CrossRef] [PubMed]
- Reinehr, S.; Reinhard, J.; Gandej, M.; Gottschalk, I.; Stute, G.; Faissner, A. S100B immunization triggers NFkappaB and complement activation in an autoimmune glaucoma model. Sci. Rep. 2018, 8, 9821. [Google Scholar] [CrossRef] [PubMed]
- Bansal, R.; Singh, R. Exploring the potential of natural and synthetic neuroprotective steroids against neurodegenerative disorders: A literature review. Med. Res. Rev. 2018, 38, 1126–1158. [Google Scholar] [CrossRef] [PubMed]
- Nebbioso, M.; Buomprisco, G.; Pascarella, A.; Pescosolido, N. Modulatory effects of 1,25-dihydroxyvitamin D3 on eye disorders: A critical review. Crit. Rev. Food Sci. Nutr. 2017, 57, 559–565. [Google Scholar] [CrossRef] [PubMed]
- Kaarniranta, K.; Xu, H.; Kauppinen, A. Mechanistical retinal drug targets and challenges. Adv. Drug Delivery Rev. 2018, 126, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Luo, C.X.; Chu, W.H.; Shan, Y.A.; Qian, Z.M.; Zhu, G.; Yu, Y.B.; Feng, H. 20-Hydroxyecdysone protects against oxidative stress-induced neuronal injury by scavenging free radicals and modulating NF-κB and JNK pathways. PLoS ONE 2012, 7, e50764. [Google Scholar] [CrossRef]
- Xie, M.; Li, X.; Li, Y.; Chen, Y.; Zhang, X.; Yin, W.; Lu, B.; Zhu, Z.; Shi, H.; Zhang, J. The synthesis of polyhydroxy sterone 2β,3α,5-trihydroxy-5α-androst-6-one and its neuroprotection. Ther. Targets Neurol. Dis. 2017, 4, e1559. [Google Scholar] [CrossRef]
- Huo, Y.; Rangarajan, P.; Ling, E.A.; Dheen, S.T. Dexamethasone inhibits the Nox-dependent ROS production via suppression of MKP-1-dependent MAPK pathways in activated microglia. BMC Neurosci. 2011, 12, 49. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, Z.; Artelt, M.; Burnet, M.; Schluesener, H.J. Dexamethasone attenuates early expression of three molecules associated with microglia/macrophages activation following rat traumatic brain injury. Acta Neuropathol. 2007, 113, 675–682. [Google Scholar] [CrossRef]
- Paranjape, A.; Chernushevich, O.; Qayum, A.A.; Spence, A.J.; Taruselli, M.T.; Abebayehu, D.; Barnstein, B.O.; McLeod, J.J.; Baker, B.; Bajaj, G.S.; et al. Dexamethasone rapidly suppresses IL-33-stimulated mast cell function by blocking transcription factor activity. J. Leukocyte Biol. 2016, 100, 1395–1404. [Google Scholar] [CrossRef] [PubMed]
- Ellis-Behnke, R.G.; Jonas, R.A.; Jonas, J.B. The microglial system in the eye and brain in response to stimuli in vivo. J. Glaucoma 2013, 22 (Suppl. 5), S32–S35. [Google Scholar] [CrossRef] [PubMed]
- Bosco, A.; Crish, S.D.; Steele, M.R.; Romero, C.O.; Inman, D.M.; Horner, P.J.; Calkins, D.J.; Vetter, M.L. Early reduction of microglia activation by irradiation in a model of chronic glaucoma. PLoS ONE 2012, 7, e43602. [Google Scholar] [CrossRef] [PubMed]
- Luo, C.; Yang, X.; Kain, A.D.; Powell, D.W.; Kuehn, M.H.; Tezel, G. Glaucomatous tissue stress and the regulation of immune response through glial Toll-like receptor signaling. Investig. Ophthalmol. Visual Sci. 2010, 51, 5697–5707. [Google Scholar] [CrossRef] [PubMed]
- Bond, W.S.; Hines-Beard, J.; GoldenMerry, Y.L.; Davis, M.; Farooque, A.; Sappington, R.M.; Calkins, D.J.; Rex, T.S. Virus-mediated EpoR76E Therapy Slows Optic Nerve Axonopathy in Experimental Glaucoma. Mol. Ther. 2016, 24, 230–239. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Luo, C.; Cai, J.; Powell, D.W.; Yu, D.; Kuehn, M.H.; Tezel, G. Neurodegenerative and inflammatory pathway components linked to TNF-alpha/TNFR1 signaling in the glaucomatous human retina. Investig. Ophthalmol. Visual Sci. 2011, 52, 8442–8454. [Google Scholar] [CrossRef] [PubMed]
- Yan, M.; Leng, T.; Tang, L.; Zheng, X.; Lu, B.; Li, Y.; Sheng, L.; Lin, S.; Shi, H.; Yan, G. Neuroprotectant androst-3β,5α,6β-triol suppresses TNF-α-induced endothelial adhesion molecules expression and neutrophil adhesion to endothelial cells by attenuation of CYLD-NF-κB pathway. Biochem. Biophys. Res. Commun. 2017, 483, 892–896. [Google Scholar] [CrossRef]
- Hu, H.; Zhou, Y.; Leng, T.; Liu, A.; Wang, Y.; You, X.; Chen, J.; Tang, L.; Chen, W.; Qiu, P.; et al. The major cholesterol metabolite cholestane-3β,5α,6β-triol functions as an endogenous neuroprotectant. J. Neurosci. 2014, 34, 11426–11438. [Google Scholar] [CrossRef] [PubMed]
- De Sa Lima, L.; Kawamoto, E.M.; Munhoz, C.D.; Kinoshita, P.F.; Orellana, A.M.; Curi, R.; Rossoni, L.V.; Avellar, M.C.; Scavone, C. Ouabain activates NFκB through an NMDA signaling pathway in cultured cerebellar cells. Neuropharmacology 2013, 73, 327–336. [Google Scholar] [CrossRef]
- Wu, Q.; Zhao, Y.; Chen, X.; Zhu, M.; Miao, C. Propofol attenuates BV2 microglia inflammation via NMDA receptor inhibition. Can. J. Physiol. Pharmacol. 2018, 96, 241–248. [Google Scholar] [CrossRef] [Green Version]
Sample Availability: Not available. |




© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, H.-J.-Q.; Xue, D.-D.; Lu, B.-Z.; Li, Y.; Sheng, L.-X.; Zhu, Z.; Zhou, Y.-W.; Zhang, J.-X.; Lin, G.-J.; Lin, S.-Z.; et al. A Novel Synthetic Steroid of 2β,3α,5α-Trihydroxy-androst-6-one Alleviates the Loss of Rat Retinal Ganglion Cells Caused by Acute Intraocular Hypertension via Inhibiting the Inflammatory Activation of Microglia. Molecules 2019, 24, 252. https://doi.org/10.3390/molecules24020252
Sun H-J-Q, Xue D-D, Lu B-Z, Li Y, Sheng L-X, Zhu Z, Zhou Y-W, Zhang J-X, Lin G-J, Lin S-Z, et al. A Novel Synthetic Steroid of 2β,3α,5α-Trihydroxy-androst-6-one Alleviates the Loss of Rat Retinal Ganglion Cells Caused by Acute Intraocular Hypertension via Inhibiting the Inflammatory Activation of Microglia. Molecules. 2019; 24(2):252. https://doi.org/10.3390/molecules24020252
Chicago/Turabian StyleSun, Hong-Jia-Qi, Dong-Dong Xue, Bing-Zheng Lu, Yuan Li, Long-Xiang Sheng, Zhu Zhu, Yu-Wei Zhou, Jing-Xia Zhang, Gan-Jian Lin, Sui-Zhen Lin, and et al. 2019. "A Novel Synthetic Steroid of 2β,3α,5α-Trihydroxy-androst-6-one Alleviates the Loss of Rat Retinal Ganglion Cells Caused by Acute Intraocular Hypertension via Inhibiting the Inflammatory Activation of Microglia" Molecules 24, no. 2: 252. https://doi.org/10.3390/molecules24020252
APA StyleSun, H.-J.-Q., Xue, D.-D., Lu, B.-Z., Li, Y., Sheng, L.-X., Zhu, Z., Zhou, Y.-W., Zhang, J.-X., Lin, G.-J., Lin, S.-Z., Yan, G.-M., Chen, Y.-P., & Yin, W. (2019). A Novel Synthetic Steroid of 2β,3α,5α-Trihydroxy-androst-6-one Alleviates the Loss of Rat Retinal Ganglion Cells Caused by Acute Intraocular Hypertension via Inhibiting the Inflammatory Activation of Microglia. Molecules, 24(2), 252. https://doi.org/10.3390/molecules24020252