
High-Level Heterologous Expression of Endo-1,4-β-Xylanase from Penicillium citrinum in Pichia pastoris X-33 Directed through Codon Optimization and Optimized Expression

Abstract
1. Introduction
2. Results and Discussion
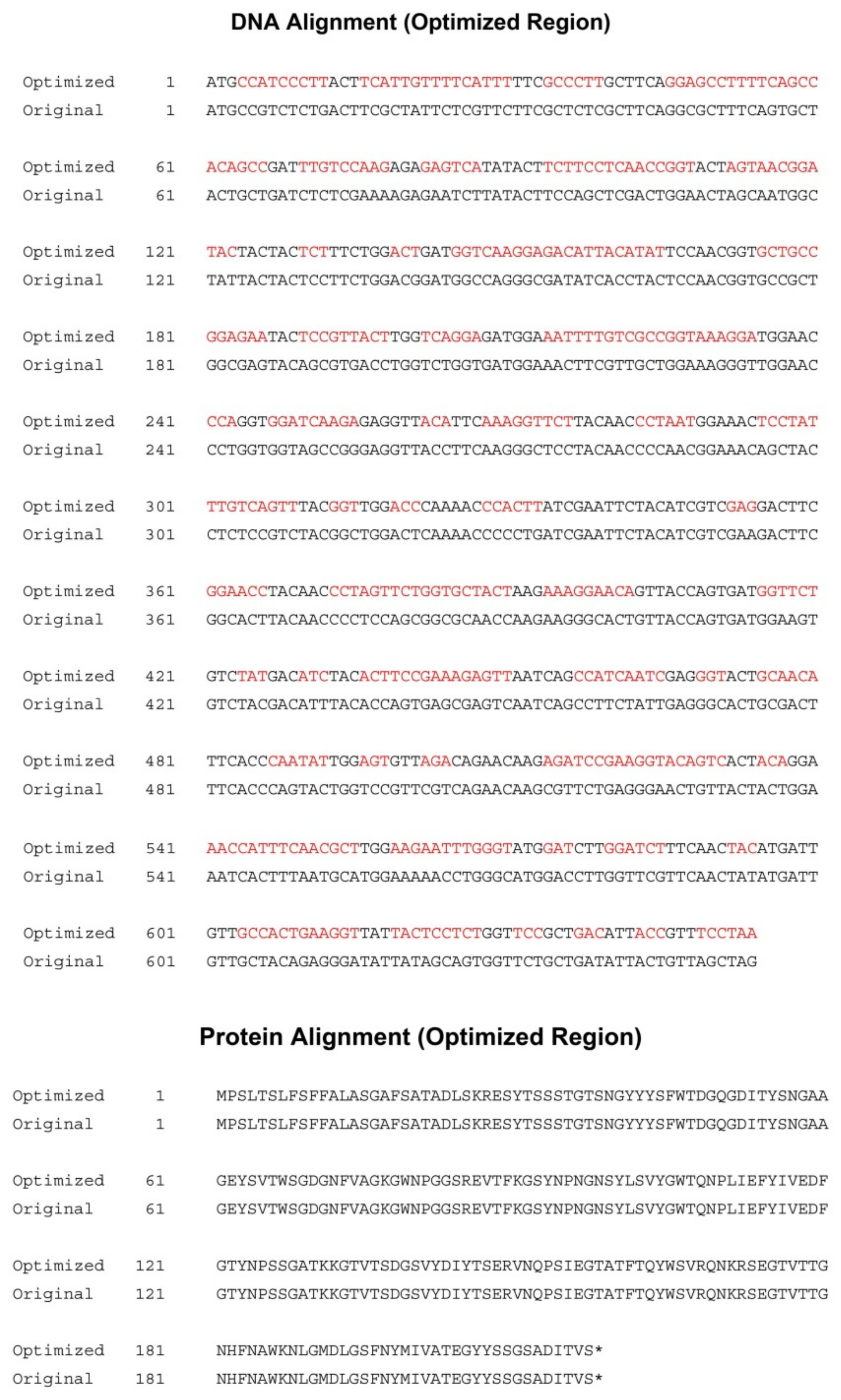
2.1. Construction of Expression Vectors, Transformation and Selection
2.2. Expression of xynA-PC in P. pastoris
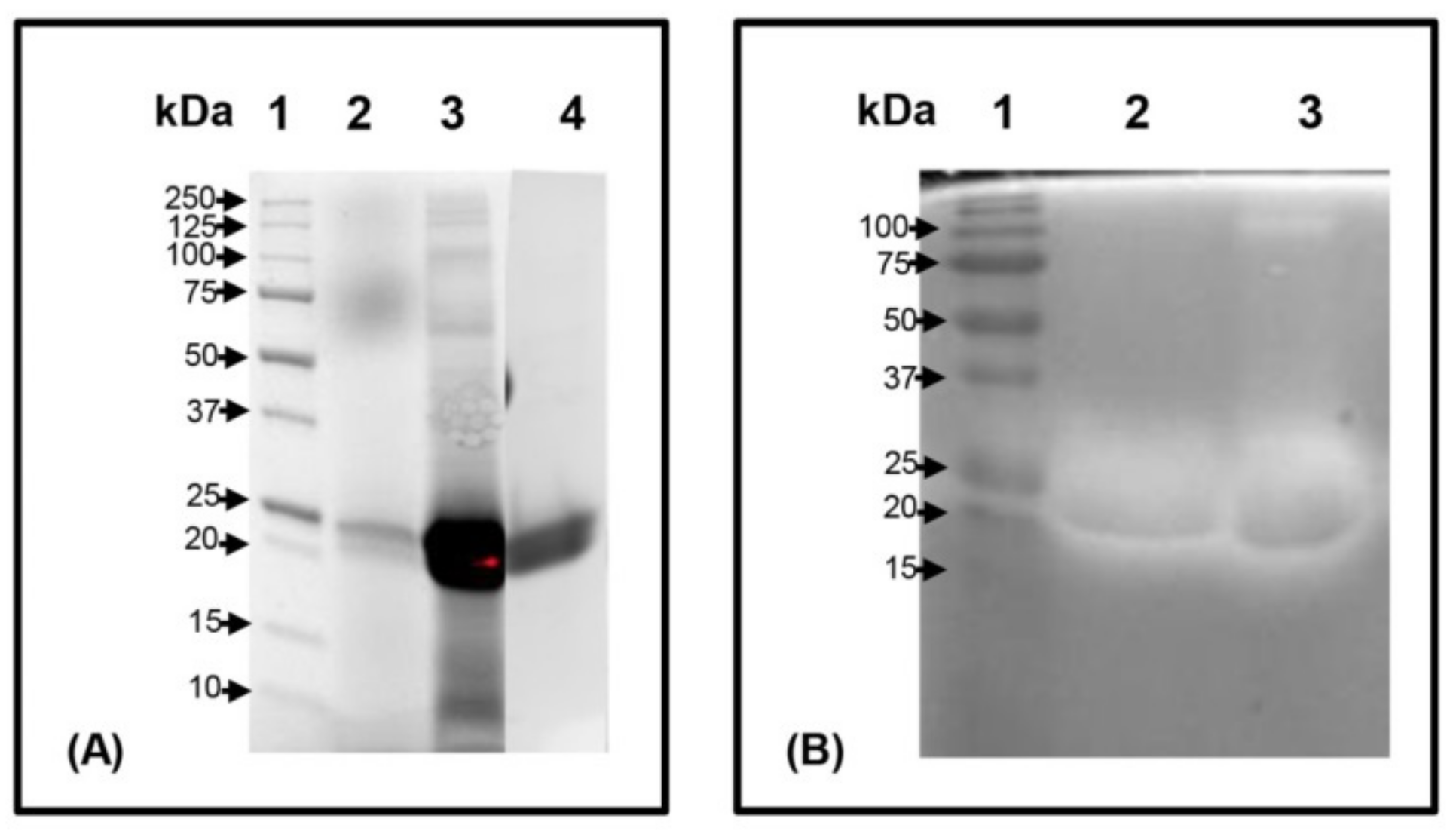
2.3. Purification, SDS-PAGE and Zymography Analysis
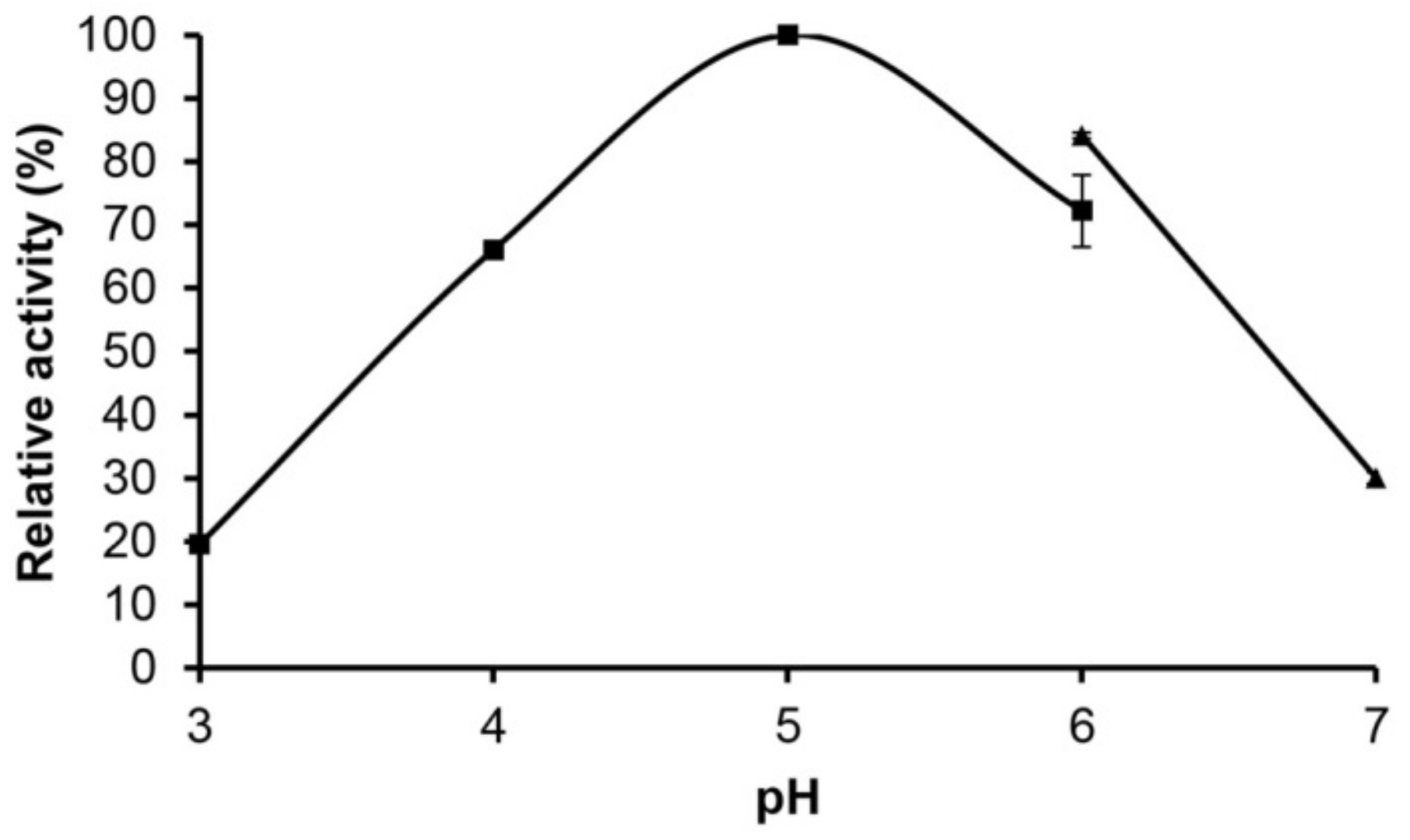
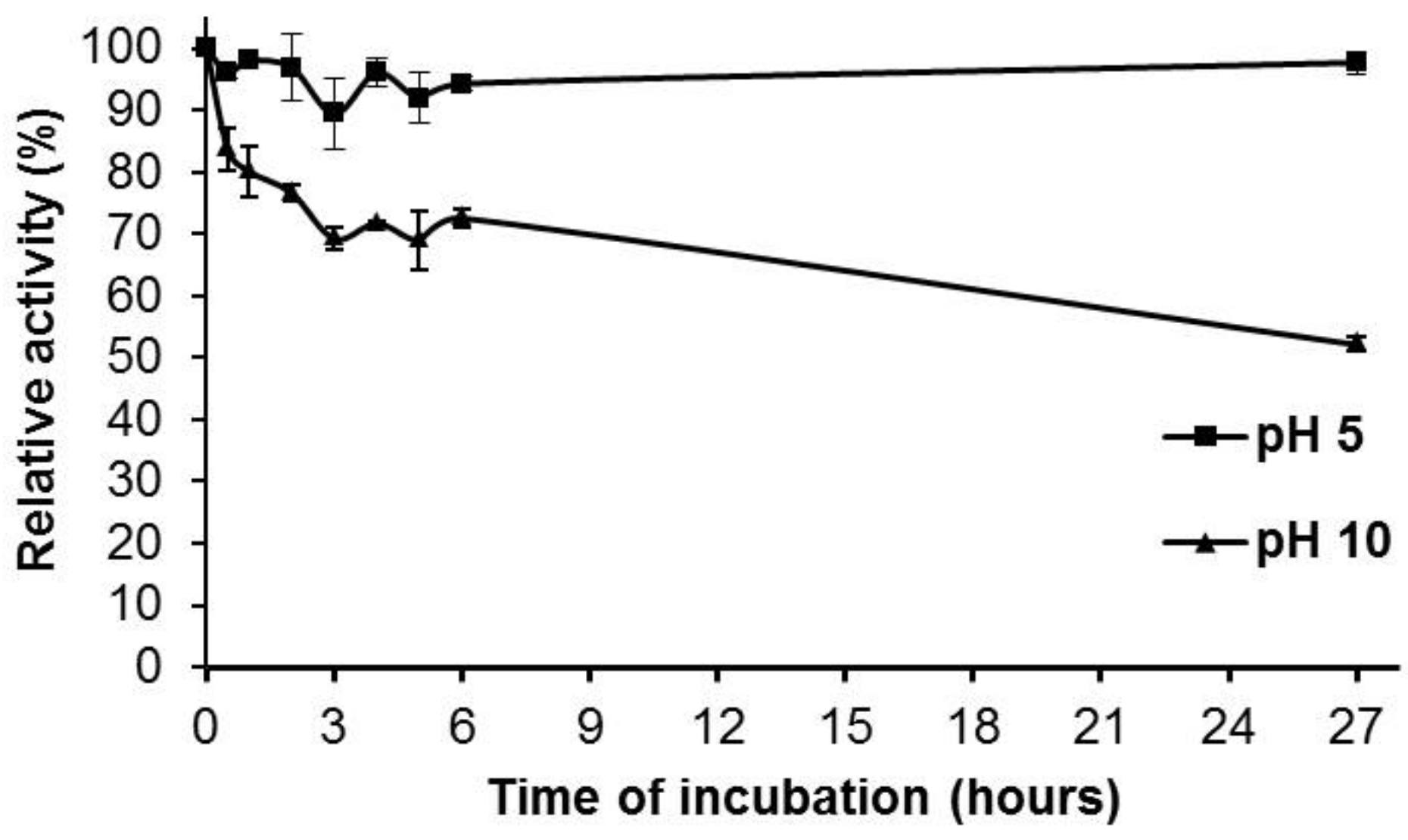
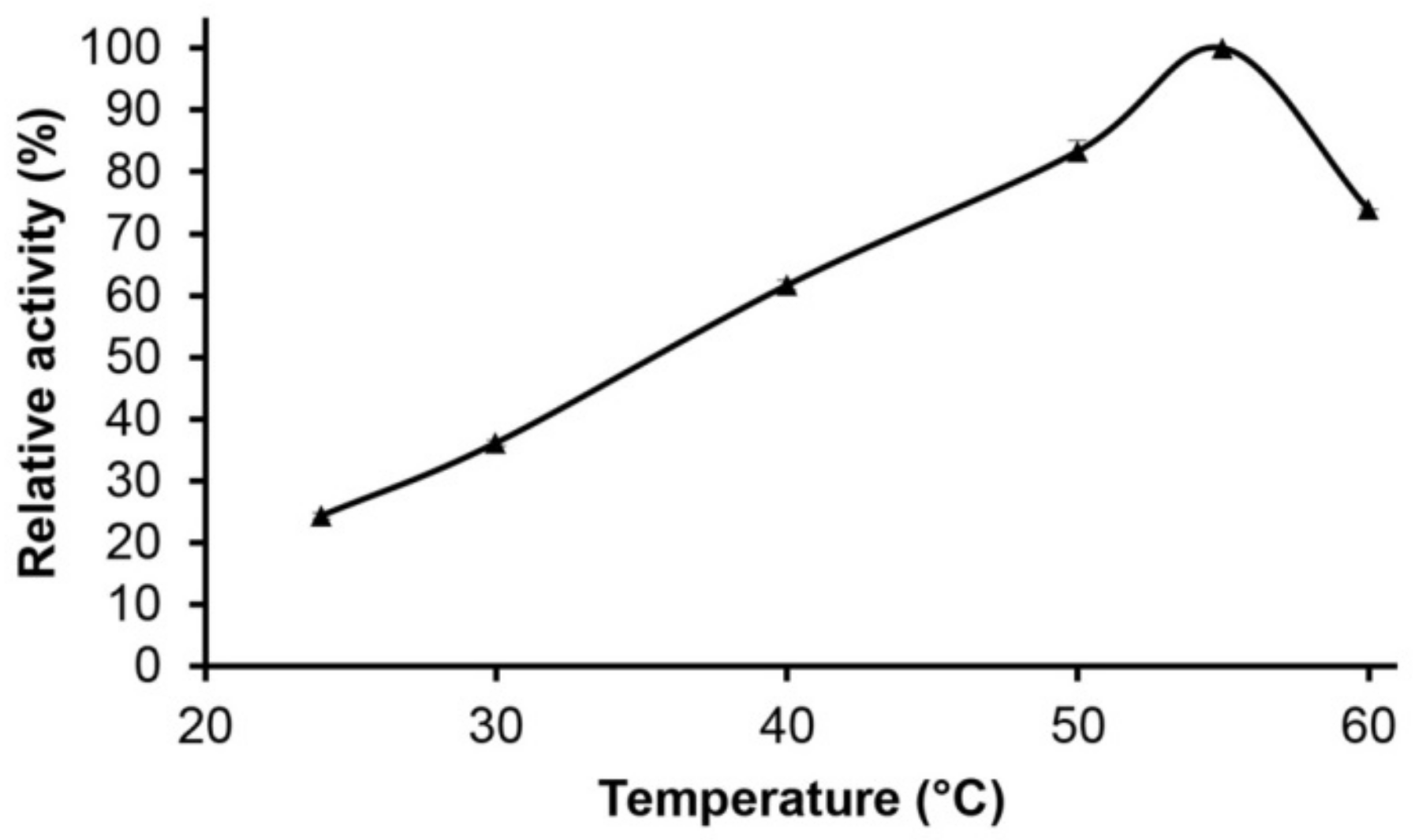
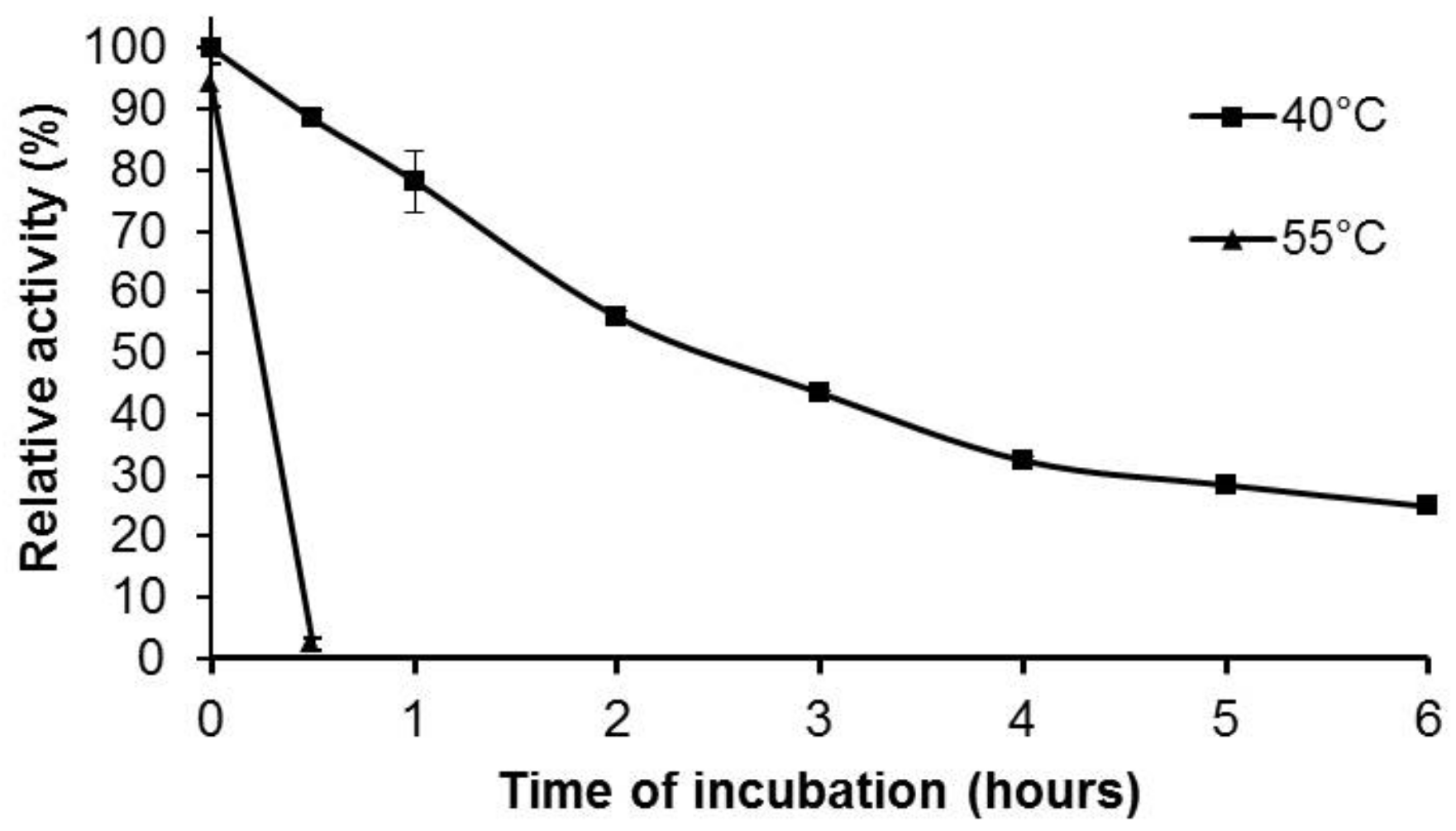
2.4. Effect of pH and Temperature
2.5. Kinetic Parameters
3. Materials and Methods
3.1. Gene, Plasmids, Strains, Culture Conditions and Materials
3.2. Construction of Expression Plasmids
3.3. Yeast Transformation and Selection
3.4. Expression of Recombinant xynA-PC in P. pastoris
3.5. Xylanase Activity Assay and Protein Determination
3.6. Purification of the Pc Xyn11A Enzyme
3.7. SDS-PAGE and Zymography Analysis
3.8. Effect of pH and Temperature on Xylanase Activity and Stability
3.9. Kinetic Parameters
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Loqué, D.; Scheller, H.V.; Pauly, M. Engineering of plant cell walls for enhanced biofuel production. Curr. Opin. Plant Biol. 2015, 25, 151–161. [Google Scholar] [CrossRef] [PubMed]
- Polizeli, M.L.T.M.; Rizzatti, A.C.S.; Monti, R.; Terenzi, H.F.; Jorge, J.A.; Amorim, D.S. Xylanases from fungi: Properties and industrial applications. Appl. Microbiol. Biotechnol. 2005, 67, 577–591. [Google Scholar] [CrossRef] [PubMed]
- Moreira, L.R.; Filho, E.X. Insights into the mechanism of enzymatic hydrolysis of xylan. Appl. Microbiol. Biotechnol. 2016, 100, 5205–5214. [Google Scholar] [CrossRef] [PubMed]
- Walia, A.; Guleria, S.; Mehta, P.; Chauhan, A.; Parkash, J. Microbial xylanases and their industrial application in pulp and paper biobleaching: A review. 3 Biotech 2017, 7, 11. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Han, J.; Li, H.; Zhang, Y.; Feng, J. The effect of enzymes on release of trace elements in feedstuffs based on in vitro digestion model for monogastric livestock. J. Anim. Sci. Biotechnol. 2018, 9, 73. [Google Scholar] [CrossRef] [PubMed]
- Maas, R.M.; Verdegem, M.C.; Dersjant-Li, Y.; Schrama, J.W. The effect of phytase, xylanase and their combination on growth performance and nutrient utilization in Nile tilapia. Aquaculture 2018, 487, 7–14. [Google Scholar] [CrossRef]
- Adigüzel, A.O.; Tunçer, M. Production, characterization and application of a xylanase from Streptomyces sp. AOA40 in fruit juice and bakery industries. Food Biotechnol. 2016, 30, 189–218. [Google Scholar] [CrossRef]
- Rosmine, E.; Sainjan, N.C.; Silvester, R.; Alikkunju, A.; Varghese, S.A. Statistical optimisation of xylanase production by estuarine Streptomyces sp. and its application in clarification of fruit juice. J. Genet. Eng. Biotechnol. 2017, 15, 393–401. [Google Scholar] [CrossRef]
- Zhao, L.; Meng, K.; Bai, Y.; Shi, P.; Huang, H.; Luo, H.; Wang, Y.; Yang, P.; Song, W.; Yao, B. Two family 11 xylanases from Achaetomium sp. Xz-8 with high catalytic efficiency and application potentials in the brewing industry. J. Agric. Food Chem. 2013, 61, 6880–6889. [Google Scholar] [CrossRef]
- Sena, L.M.; Morais, C.G.; Lopes, M.R.; Santos, R.O.; Uetanabaro, A.P.; Morais, P.B.; Vital, M.J.; de Morais, M.A., Jr.; Lachance, M.A.; Rosa, C.A. D-Xylose fermentation, xylitol production and xylanase activities by seven new species of Sugiyamaella. Antonie Van Leeuwenhoek 2017, 110, 53–67. [Google Scholar] [CrossRef]
- Uday, U.S.P.; Choudhury, P.; Bandyopadhyay, T.K.; Bhunia, B. Classification, mode of action and production strategy of xylanase and its application for biofuel production from water hyacinth. Int. J. Biol. Macromol. 2016, 82, 1041–1054. [Google Scholar] [CrossRef] [PubMed]
- Chakdar, H.; Kumar, M.; Pandiyan, K.; Singh, A.; Nanjappan, K.; Kashyap, P.L.; Srivastava, A.K. Bacterial xylanases: Biology to biotechnology. 3 Biotech 2016, 6, 150. [Google Scholar] [CrossRef] [PubMed]
- Shi, W.; Ding, S.Y.; Yuan, J.S. Comparison of Insect Gut Cellulase and Xylanase Activity Across Different Insect Species with Distinct Food Sources. Bioenerg. Res. 2011, 4, 1–10. [Google Scholar] [CrossRef]
- Lara, C.A.; Santos, R.O.; Cadete, R.M.; Ferreira, C.; Marques, S.; Gírio, F.; Oliveira, E.S.; Rosa, C.A.; Fonseca, C. Identification and characterisation of xylanolytic yeasts isolated from decaying wood and sugarcane bagasse in Brazil. Antonie Van Leeuwenhoek 2014, 105, 1107–1119. [Google Scholar] [CrossRef] [PubMed]
- Suda, C.N.K.; Buckeridge, M.S.; Giorgini, J.F. Cell wall hydrolases in the seeds of Euphorbia heterophylla L. during germination and early seedling development. Braz. J. Plant Physiol. 2003, 15, 135–143. [Google Scholar] [CrossRef]
- Lu, F.; Lu, M.; Lu, Z.; Bie, X.; Zhao, H.; Wang, Y. Purification and characterization of xylanase from Aspergillus ficuum AF-98. Bioresour. Technol. 2008, 99, 5938–5941. [Google Scholar] [CrossRef]
- Wu, J.; Qiu, C.; Ren, Y.; Yan, R.; Ye, X.; Wang, G. Novel Salt-Tolerant Xylanase from a Mangrove-Isolated Fungus Phoma sp. MF13 and Its Application in Chinese Steamed Bread. ACS Omega 2018, 3, 3708–3716. [Google Scholar] [CrossRef]
- Roy, S.; Dutta, T.; Sarkar, T.S.; Ghosh, S. Novel xylanases from Simplicillium obclavatum MTCC 9604: Comparative analysis of production, purification and characterization of enzyme from submerged and solid state fermentation. Springerplus 2013, 2, 382. [Google Scholar] [CrossRef]
- Guan, G.Q.; Zhao, P.X.; Zhao, J.; Wang, M.J.; Huo, S.H.; Cui, F.J.; Jiang, J.X. Production and Partial Characterization of an Alkaline Xylanase from a Novel Fungus Cladosporium oxysporum. Biomed. Res. Int. 2016, 2016, 4575024. [Google Scholar] [CrossRef]
- Raghukumar, C.; Muraleedharan, U.; Gaud, V.R.; Mishra, R. Xylanases of marine fungi of potential use for biobleaching of paper pulp. J. Ind. Microbiol. Biotechnol. 2004, 31, 433–441. [Google Scholar] [CrossRef]
- Tanaka, H.; Nakamura, T.; Hayashi, S.; Ohta, K. Purification and properties of an extracellular endo-1,4-beta-xylanase from Penicillium citrinum and characterization of the encoding gene. J. Biosci. Bioeng. 2005, 100, 623–630. [Google Scholar] [CrossRef] [PubMed]
- Bagewadi, Z.K.; Mulla, S.I.; Mulla, Y.; Ninnekar, H.Z. Xylanase production from Penicillium citrinum isolate HZN13 using response surface methodology and characterization of immobilized xylanase on glutaraldehyde-activated calcium-alginate beads. 3 Biotech 2016, 6, 164. [Google Scholar] [CrossRef] [PubMed]
- Kang, Z.; Huang, H.; Zhang, Y.; Du, G.; Chen, J. Recent advances of molecular toolbox construction expand Pichia pastoris in synthetic biology applications. World J. Microbiol. Biotechnol. 2017, 33, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Schwarzhans, J.P.; Luttermann, T.; Geier, M.; Kalinowski, J.; Friehs, K. Towards systems metabolic engineering in Pichia pastoris. Biotechnol. Adv. 2017, 35, 681–710. [Google Scholar] [CrossRef] [PubMed]
- Weinacker, D.; Rabert, C.; Zepeda, A.B.; Figueroa, C.A.; Pessoa, A.; Farías, J.G. Applications of recombinant Pichia pastoris in the healthcare industry. Braz. J. Microbiol. 2013, 44, 1043–1048. [Google Scholar] [CrossRef] [PubMed]
- Spohner, S.C.; Müller, H.; Quitmann, H.; Czermak, P. Expression of enzymes for the usage in food and feed industry with Pichia pastoris. J. Biotechnol. 2015, 202, 118–134. [Google Scholar] [CrossRef]
- Elena, C.; Ravasi, P.; Castelli, M.E.; Peirú, S.; Menzella, H.G. Expression of codon optimized genes in microbial systems: Current industrial applications and perspectives. Front. Microbiol. 2014, 5, 21. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Kong, Q.; Zhang, D.; Yan, L. Codon optimization significantly enhanced the expression of human 37-kDa iLRP in Escherichia coli. 3 Biotech 2018, 8, 210. [Google Scholar] [CrossRef]
- Waterham, H.R.; Digan, M.E.; Koutz, P.J.; Lair, S.V.; Cregg, J.M. Isolation of the Pichia pastoris glyceraldehyde-3-phosphate dehydrogenase gene and regulation and use of its promoter. Gene 1997, 186, 37–44. [Google Scholar] [CrossRef]
- Döring, F.; Klapper, M.; Theis, S.; Daniel, H. Use of the glyceraldehyde-3-phosphate dehydrogenase promoter for production of functional mammalian membrane transport proteins in the yeast Pichia pastoris. Biochem. Biophys. Res. Commun. 1998, 250, 531–535. [Google Scholar] [CrossRef]
- Delroisse, J.M.; Dannau, M.; Gilsoul, J.J.; El Mejdoub, T.; Destain, J.; Portetelle, D.; Thonart, P.; Haubruge, E.; Vandenbol, M. Expression of a synthetic gene encoding a Tribolium castaneum carboxylesterase in Pichia pastoris. Protein Expr. Purif. 2005, 42, 286–294. [Google Scholar] [CrossRef]
- Menendez, J.; Hernandez, L.; Banguela, A.; Pais, J. Functional production and secretion of the Gluconoacetobacter diazatrophicus fructose-releasing exo-levanase (LsdB) in Pichia pastoris. Enz. Microb. Technol. 2004, 34, 446–452. [Google Scholar] [CrossRef]
- Mulder, K.C.; de Lima, L.A.; Aguiar, P.S.; Carneiro, F.C.; Franco, O.L.; Dias, S.C.; Parachin, N.S. Production of a modified peptide clavanin in Pichia pastoris: Cloning, expression, purification and in vitro activities. AMB Express 2015, 5, 129. [Google Scholar] [CrossRef]
- Kim, S.J.; Lee, J.A.; Kim, Y.H.; Song, B.K. Optimization of the functional expression of Coprinus cinereus peroxidase in Pichia pastoris by varying the host and promoter. J. Microbiol. Biotechnol. 2009, 19, 966–971. [Google Scholar] [CrossRef] [PubMed]
- Boer, H.; Teeri, T.T.; Koivula, A. Characterization of Trichoderma reesei cellobiohydrolase Cel7A secreted from Pichia pastoris using two different promoters. Biotechnol. Bioeng. 2000, 69, 486–494. [Google Scholar] [CrossRef]
- Ohta, K.; Tanaka, H.; Yamakawa, D.; Hamasuna, H.; Fujimoto, H. Signal peptide of Aureobasidium pullulans xylanase: Use for extracellular production of a fungal xylanase by Escherichia coli. J. Ind. Microbiol. Biotechnol. 2011, 38, 967–973. [Google Scholar] [CrossRef] [PubMed]
- Ouephanit, C.; Boonvitthya, N.; Theerachat, M.; Bozonnet, S.; Chulalaksananukul, W. Efficient expression and secretion of endo-1,4-β-xylanase from Penicillium citrinum in non-conventional yeast Yarrowia lipolytica directed by the native and the preproLIP2 signal peptides. Protein. Expr. Purif. 2019, 160, 1–6. [Google Scholar] [CrossRef]
- Uchima, C.A.; Arioka, M. Expression and one-step purification of recombinant proteins using an alternative episomal vector for the expression of N-tagged heterologous proteins in Pichia pastoris. Biosci. Biotechnol. Biochem. 2012, 76, 368–371. [Google Scholar] [CrossRef][Green Version]
- Zahrl, R.J.; Peña, D.A.; Mattanovich, D.; Gasser, B. Systems biotechnology for protein production in Pichia pastoris. FEMS Yeast Res. 2017, 17, fox068. [Google Scholar] [CrossRef]
- Collins, T.; Gerday, C.; Feller, G. Xylanases, xylanase families and extremophilic xylanases. FEMS Microbiol. Rev. 2005, 29, 3–23. [Google Scholar] [CrossRef]
- Kulkarni, N.; Shendye, A.; Rao, M. Molecular and biotechnological aspects of xylanases. FEMS Microbiol. Rev. 1999, 23, 411–456. [Google Scholar] [CrossRef] [PubMed]
- Do, T.T.; Quyen, D.T.; Dam, T.H. Purification and characterization of an acidstable and organic solvent-tolerant xylanase from Aspergillus awamori VTCCF312. Sci. Asia 2012, 38, 157–165. [Google Scholar]
- Barel, M.; Harduin-Lepers, A.; Portier, L.; Slomianny, M.; Charbit, A. Host glycosylation pathways and the unfolded protein response contribute to the infection by Francisella. Cell Microbiol. 2016, 18, 1763–1781. [Google Scholar] [CrossRef] [PubMed]
- Spiro, R.G. Protein glycosylation: Nature, distribution, enzymatic formation, and disease implications of glycopeptide bonds. Glycobiology 2002, 12, 43R–56R. [Google Scholar] [CrossRef] [PubMed]
- Jeya, M.; Thiagarajan, S.; Lee, J.K.; Gunasekaran, P. Cloning and expression of GH11 xylanase gene from Aspergillus fumigatus MKU1 in Pichia pastoris. J. Biosci. Bioeng. 2009, 108, 24–29. [Google Scholar] [CrossRef] [PubMed]
- Qu, W.; Shao, W. Cloning, expression and characterization of glycoside hydrolase family 11 endoxylanase from Bacillus pumilus ARA. Biotechnol. Lett. 2011, 33, 1407–1416. [Google Scholar] [CrossRef]
- Chantorn, S.; Piyapittayanun, C.; Khumphai, P.; Pannanusorn, S.; Phannachet, K.; Apiraksakorn, J. Suitable conditions for xylanases activities from Bacillus sp. GA2(1) and Bacillus sp. GA1(6) and their properties for agricultural residues hydrolysis. Songklanakarin J. Sci. Technol. 2018, 38, 177–182. [Google Scholar]
- Liao, H.; Sun, S.; Wang, P.; Bi, W.; Tan, S.; Wei, Z.; Mei, X.; Liu, D.; Raza, W.; Shen, Q.; et al. A new acidophilic endo-β-1,4-xylanase from Penicillium oxalicum: Cloning, purification, and insights into the influence of metal ions on xylanase activity. J. Ind. Microbiol. Biotechnol. 2014, 41, 1071–1083. [Google Scholar] [CrossRef]
- Ahmad, Z.; Butt, M.S.; Ahmed, A.; Riaz, M.; Sabir, S.M.; Farooq, U.; Rehman, F.U. Effect of Aspergillus niger xylanase on dough characteristics and bread quality attributes. J. Food Sci. Technol. 2014, 51, 2445–2453. [Google Scholar] [CrossRef]
- Fan, G.; Yang, S.; Yan, Q.; Guo, Y.; Li, Y.; Jiang, Z. Characterization of a highly thermostable glycoside hydrolase family 10 xylanase from Malbranchea cinnamomea. Int. J. Biol. Macromol. 2014, 70, 482–489. [Google Scholar] [CrossRef]
- Sun, J.; Shi, S.; Wu, J.; Xie, R.; Geng, A.; Zhu, D.C. Characterization of a salt-tolerant and cold-adapted xylanase from Bacillus cellulosilyticus. BioResources 2016, 11, 8875–8889. [Google Scholar] [CrossRef]
- Berrin, J.G.; Williamson, G.; Puigserver, A.; Chaix, J.C.; McLauchlan, W.R.; Juge, N. High-level production of recombinant fungal endo-beta-1,4-xylanase in the methylotrophic yeast Pichia pastoris. Protein. Expr. Purif. 2000, 19, 179–187. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Siika-Aho, M.; Puranen, T.; Tang, M.; Tenkanen, M.; Viikari, L. Thermostable recombinant xylanases from Nonomuraea flexuosa and Thermoascus aurantiacus show distinct properties in the hydrolysis of xylans and pretreated wheat straw. Biotechnol. Biofuels 2011, 4, 12. [Google Scholar] [CrossRef] [PubMed]
- Goettig, P. Effects of Glycosylation on the Enzymatic Activity and Mechanisms of Proteases. Int. J. Mol. Sci. 2016, 17, 1969. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Letchworth, G.J. High efficiency transformation by electroporation of Pichia pastoris pretreated with lithium acetate and dithiothreitol. BioTechniques 2004, 36, 152–154. [Google Scholar] [CrossRef] [PubMed]
- Miller, G.L. Use of dinitrosalicylic acid reagent for determinating of reducing sugars. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are not available. |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microorganism | Activity (µmol/mL/min) | Comparison with The Native Strain (fold) | References |
|---|---|---|---|
| P. citrinum FERM P-15944 the native stain | 3.5 | - | [21] |
| P. pastoris GS115 + pPIC3.5 | 17 | 5 | [21] |
| E. coli BL21(DE3) + pEXP401 | 16.8 | 5 | [36] |
| Y. lipolytica zeta + JMP62UraTEF_natXynAPc | 38 | 11 | [37] |
| Y.lipolytica zeta + JMP62UraTEF_fusXynAPc | 180.3 | 52 | [37] |
| P. pastoris X-33 + pGAPZαA-xynA | 119.5 | 34 | this study |
| P. pastoris X-33 + pPICZαA-xynA | 676 | 193 | this study |
| Step | Total Activity (U) | Total Protein (mg) | Specific Activity (U/mg) | Purification (fold) | Yield (%) |
|---|---|---|---|---|---|
| Culture supernatant | 81118 | 2175.57 | 37.29 | 1.00 | 100 |
| Ultrafiltration (10 kDa MWCO) | 77305 | 533.25 | 144.97 | 3.89 | 95.3 |
| Q Sepharose anion exchange | 21425 | 17.02 | 1258.96 | 33.77 | 26.4 |
| Enzyme | Km(app) (mg/mL) | kcat (s−1) | kcat/Km(app) (mL/mg/s) | References |
|---|---|---|---|---|
| Native XylAb | 6.8 | 85 | 12.5 | [52] |
| reXylAc | 12.6 | 150 | 11.9 | [52] |
| Nf Xyn11Ad | 6 | 136.9 | 22.8 | [53] |
| re-XynAe | 5.2 | 245 | 47.1 | [37] |
| Pc Xyn11A | 2.8 | 243 | 86.8 | this study |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ouephanit, C.; Boonvitthya, N.; Bozonnet, S.; Chulalaksananukul, W. High-Level Heterologous Expression of Endo-1,4-β-Xylanase from Penicillium citrinum in Pichia pastoris X-33 Directed through Codon Optimization and Optimized Expression. Molecules 2019, 24, 3515. https://doi.org/10.3390/molecules24193515
Ouephanit C, Boonvitthya N, Bozonnet S, Chulalaksananukul W. High-Level Heterologous Expression of Endo-1,4-β-Xylanase from Penicillium citrinum in Pichia pastoris X-33 Directed through Codon Optimization and Optimized Expression. Molecules. 2019; 24(19):3515. https://doi.org/10.3390/molecules24193515
Chicago/Turabian StyleOuephanit, Chanika, Nassapat Boonvitthya, Sophie Bozonnet, and Warawut Chulalaksananukul. 2019. "High-Level Heterologous Expression of Endo-1,4-β-Xylanase from Penicillium citrinum in Pichia pastoris X-33 Directed through Codon Optimization and Optimized Expression" Molecules 24, no. 19: 3515. https://doi.org/10.3390/molecules24193515
APA StyleOuephanit, C., Boonvitthya, N., Bozonnet, S., & Chulalaksananukul, W. (2019). High-Level Heterologous Expression of Endo-1,4-β-Xylanase from Penicillium citrinum in Pichia pastoris X-33 Directed through Codon Optimization and Optimized Expression. Molecules, 24(19), 3515. https://doi.org/10.3390/molecules24193515