
New Functional Ingredients Based on Microencapsulation of Aqueous Anthocyanin-Rich Extracts Derived from Black Rice (Oryza sativa L.)

 ,
, 
 ,
,  ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
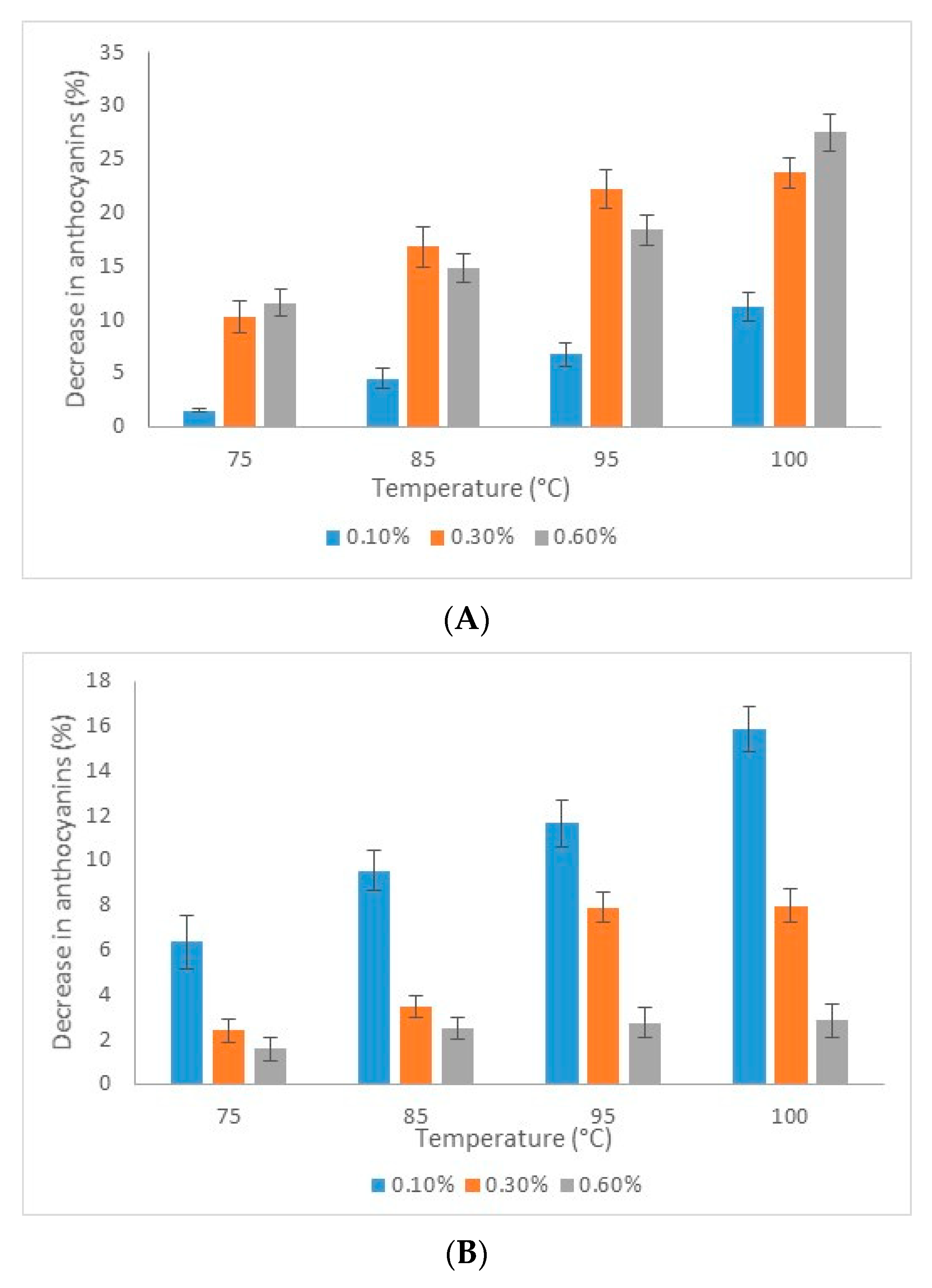
Abstract
Share and Cite
Aprodu, I.; Milea, Ș.A.; Anghel, R.-M.; Enachi, E.; Barbu, V.; Crăciunescu, O.; Râpeanu, G.; Bahrim, G.E.; Oancea, A.; Stănciuc, N. New Functional Ingredients Based on Microencapsulation of Aqueous Anthocyanin-Rich Extracts Derived from Black Rice (Oryza sativa L.). Molecules 2019, 24, 3389. https://doi.org/10.3390/molecules24183389
Aprodu I, Milea ȘA, Anghel R-M, Enachi E, Barbu V, Crăciunescu O, Râpeanu G, Bahrim GE, Oancea A, Stănciuc N. New Functional Ingredients Based on Microencapsulation of Aqueous Anthocyanin-Rich Extracts Derived from Black Rice (Oryza sativa L.). Molecules. 2019; 24(18):3389. https://doi.org/10.3390/molecules24183389
Chicago/Turabian StyleAprodu, Iuliana, Ștefania Adelina Milea, Roxana-Mădălina Anghel, Elena Enachi, Vasilica Barbu, Oana Crăciunescu, Gabriela Râpeanu, Gabriela Elena Bahrim, Anca Oancea, and Nicoleta Stănciuc. 2019. "New Functional Ingredients Based on Microencapsulation of Aqueous Anthocyanin-Rich Extracts Derived from Black Rice (Oryza sativa L.)" Molecules 24, no. 18: 3389. https://doi.org/10.3390/molecules24183389
APA StyleAprodu, I., Milea, Ș. A., Anghel, R.-M., Enachi, E., Barbu, V., Crăciunescu, O., Râpeanu, G., Bahrim, G. E., Oancea, A., & Stănciuc, N. (2019). New Functional Ingredients Based on Microencapsulation of Aqueous Anthocyanin-Rich Extracts Derived from Black Rice (Oryza sativa L.). Molecules, 24(18), 3389. https://doi.org/10.3390/molecules24183389