
Study on the Volatile Organic Compounds and Its Correlation with Water Dynamics of Bigeye Tuna (Thunnus obesus) during Cold Storage


Abstract
1. Introduction
2. Results and Discussion
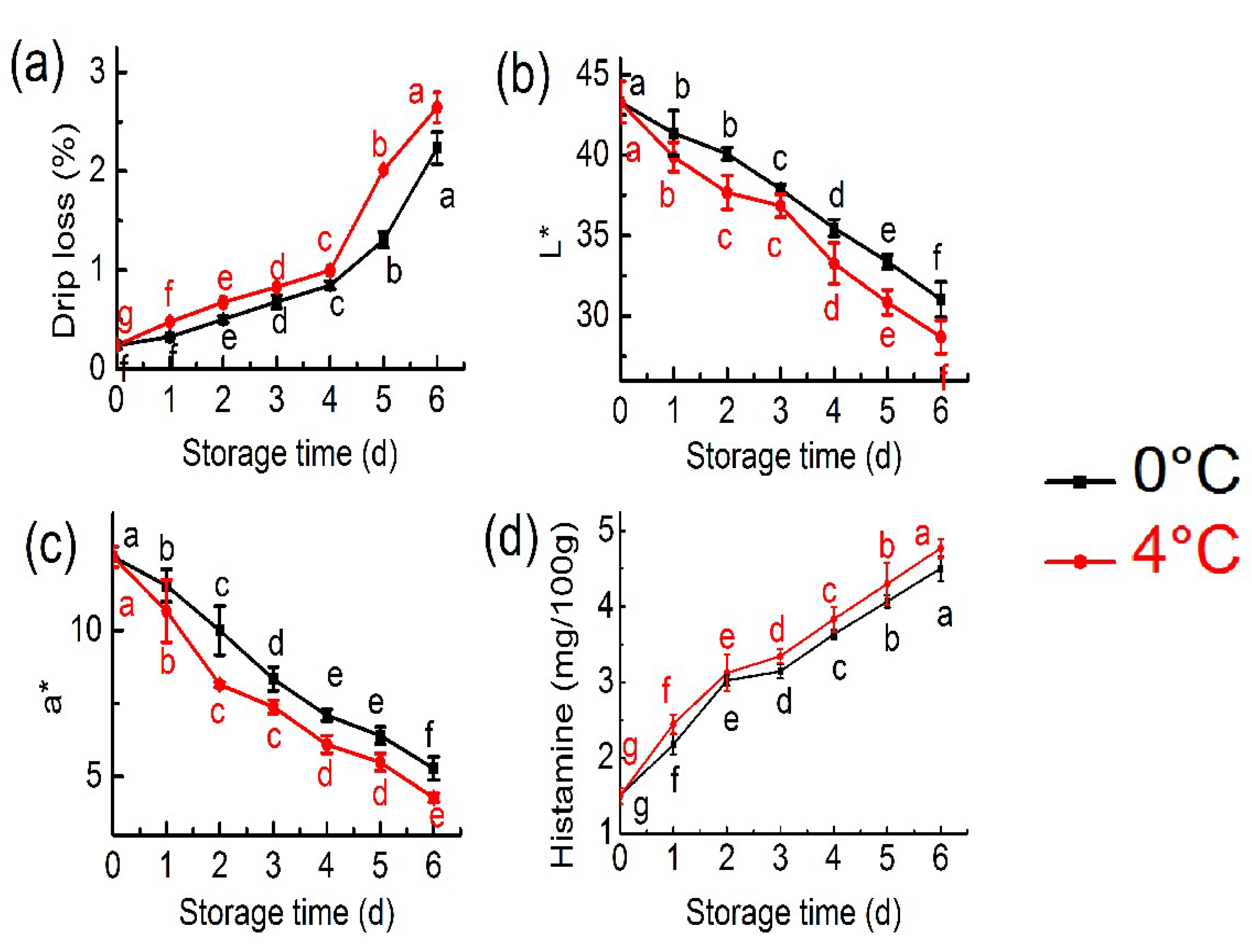
2.1. Physical and Chemical Properties Analysis
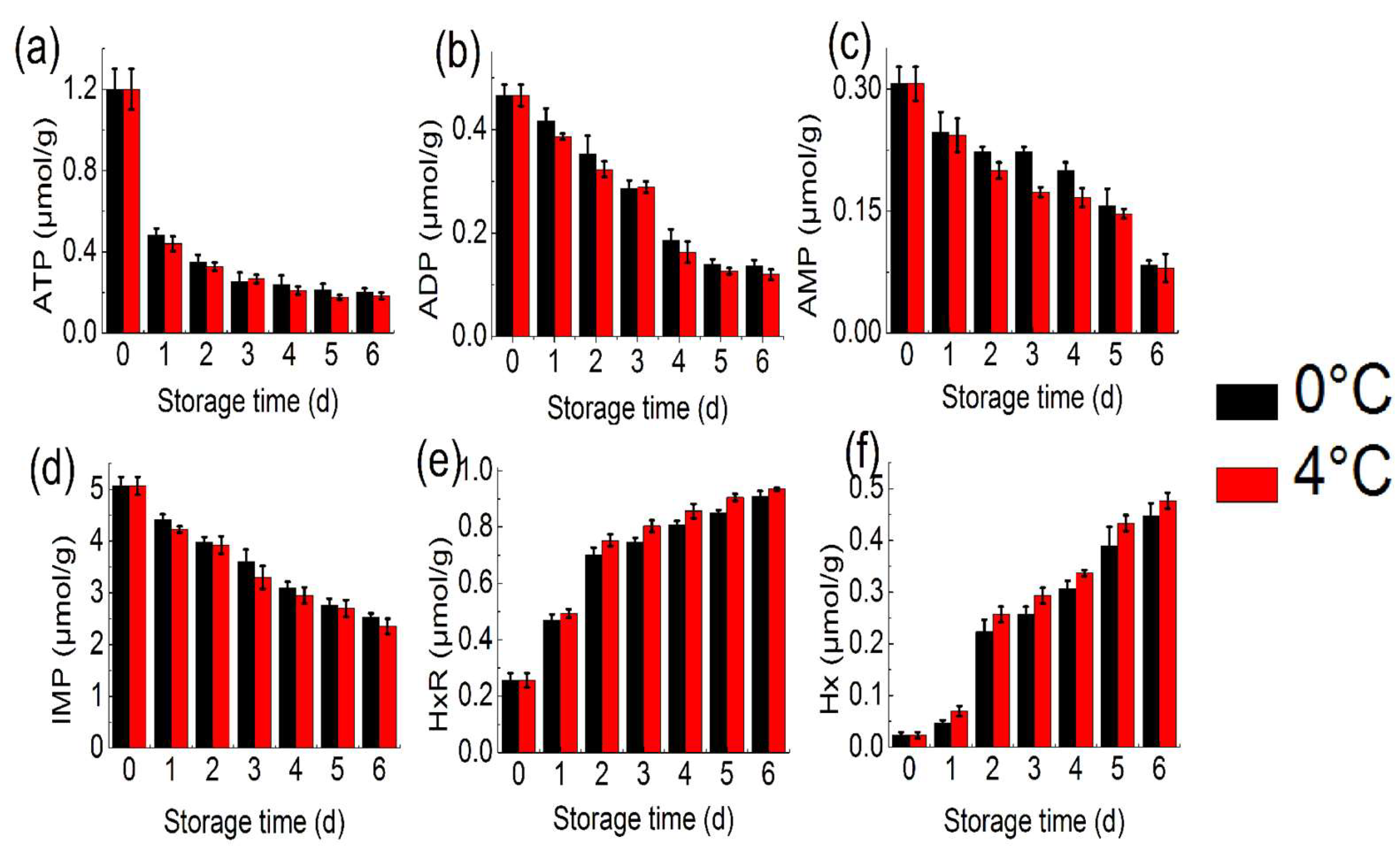
2.2. ATP-Related Compound Analysis
2.3. Headspace SPME-GC/MS Analysis of VOCs
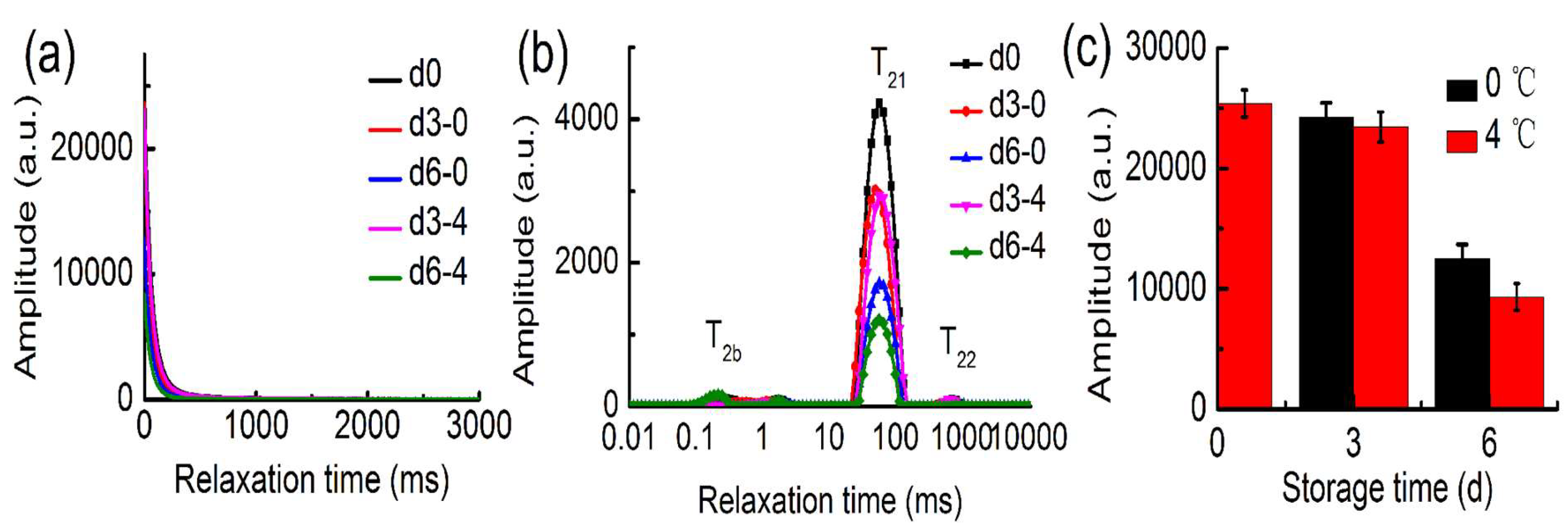
2.4. LF-NMR Analysis
2.5. Histological Analysis in Bigeye Tuna Muscle Tissue
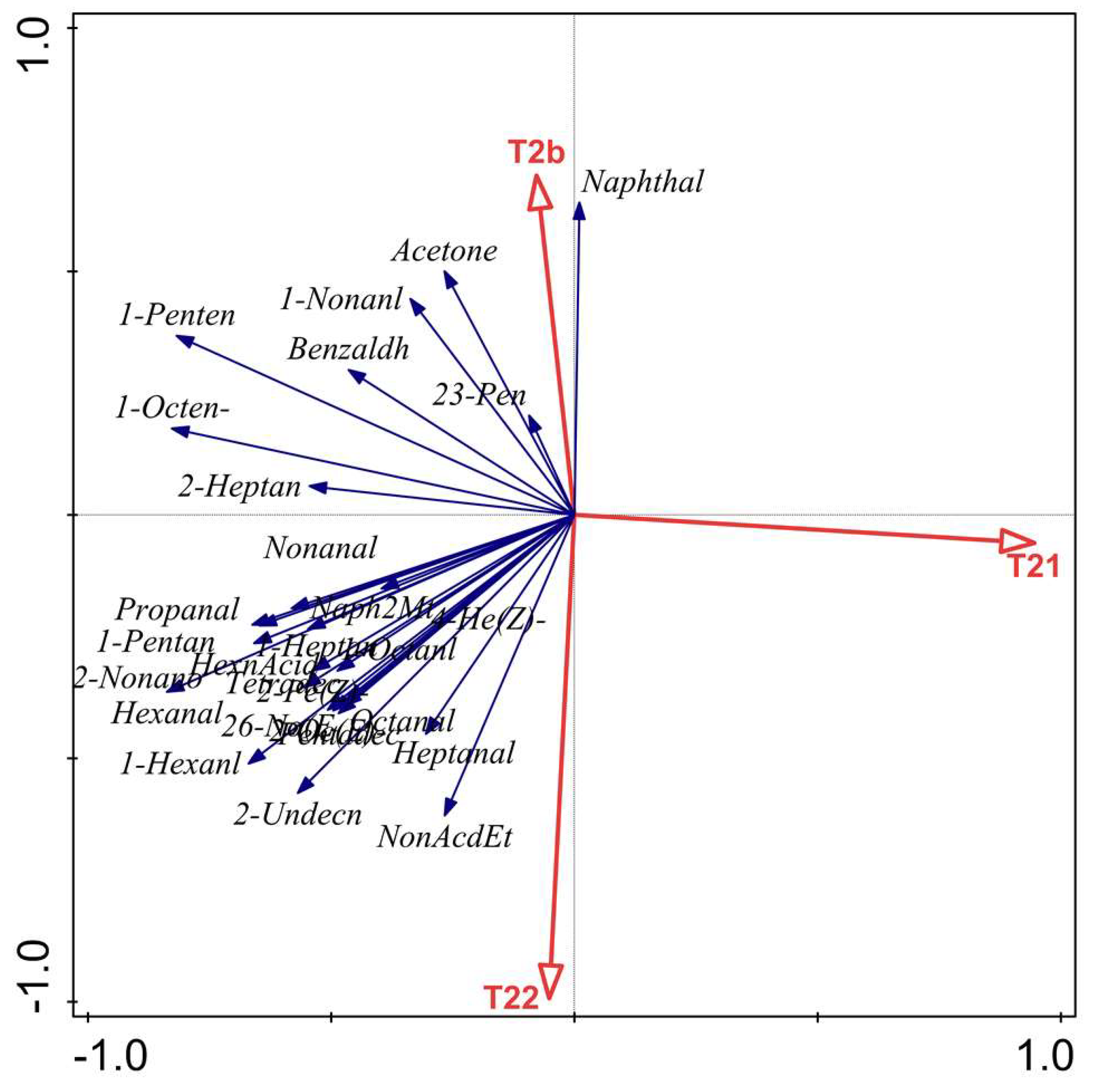
2.6. Relationship between LF-NMR Parameters (T2b, T21 and T22) and VOCs
3. Materials and Methods
3.1. Sample Preparation
3.2. Physical and Chemical Property Determination
3.2.1. Drip Loss and Colour Measurement Analysis
3.2.2. Histamine Analysis
3.3. ATP-Related Compounds Analysis
3.4. VOCs Determination by SPME-GC/MS Analysis
3.5. LF-NMR
3.6. Haematoxylin and Eosin (HE) Staining of Bigeye Tuna Muscle
3.7. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Ruiz-Capillas, C.; Moral, A. Sensory and biochemical aspects of quality of whole bigeye tuna (thunnus obesus) during bulk storage in controlled atmospheres. Food Chem. 2005, 89, 347–354. [Google Scholar] [CrossRef]
- Roiha, I.S.; Jónsson, Á.; Backi, C.J.; Lunestad, B.T.; Karlsdóttir, M.G. A comparative study of quality and safety of atlantic cod (gadus morhua) fillets during cold storage, as affected by different thawing methods of pre-rigor frozen headed and gutted fish. J. Sci. Food Agric. 2018, 98, 400–409. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Forsyth, D.; Belisle, S.; Beraldin, F.; Sparling, M.; Trudelle, R.-A.; Lapointe, P.; Bellon-Gagnon, P. Comparison of two analytical methods for the analysis of methylmercury in fish. Food Anal. Methods 2012, 6, 157–163. [Google Scholar] [CrossRef]
- Pearce, K.L.; Rosenvold, K.; Andersen, H.J.; Hopkins, D.L. Water distribution and mobility in meat during the conversion of muscle to meat and ageing and the impacts on fresh meat quality attributes—A review. Meat Sci. 2011, 89, 111–124. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Peng, Z.; Lu, J.; Li, B.; Hou, H. Self-degradation of sea cucumber body wall under 4c storage condition. J. Food Process. Preserv. 2016, 40, 715–723. [Google Scholar] [CrossRef]
- Koizumi, M.; Naito, S.; Ishida, N.; Haishi, T.; Kano, H. A dedicated mri for food science and agriculture. Food Sci. Technol. Res. 2008, 14, 74–82. [Google Scholar] [CrossRef][Green Version]
- Wang, X.; Geng, L.; Xie, J.; Qian, Y.-F. Relationship between water migration and quality changes of yellowfin tuna (thunnus albacares) during storage at 0 °C and 4 °C by lf-nmr. J. Aquat. Food Prod. Technol. 2017, 1–13. [Google Scholar] [CrossRef]
- Yu, D.; Xu, Y.; Regenstein, J.M.; Xia, W.; Yang, F.; Jiang, Q.; Wang, B. The effects of edible chitosan-based coatings on flavor quality of raw grass carp (ctenopharyngodon idellus) fillets during refrigerated storage. Food Chem. 2018, 242, 412–420. [Google Scholar] [CrossRef] [PubMed]
- Kuda, T.; Fujita, M.; Goto, H.; Yano, T. Effects of retort conditions on atp-related compounds in pouched fish muscle. Lwt - Food Sci. Technol. 2008, 41, 469–473. [Google Scholar] [CrossRef]
- Pongsetkul, J.; Benjakul, S.; Vongkamjan, K.; Sumpavapol, P.; Osako, K.; Faithong, N. Changes in volatile compounds, atp-related compounds and antioxidative properties of kapi, produced from acetes vulgaris, during processing and fermentation. Food Biosci. 2017, 19, 49–56. [Google Scholar] [CrossRef]
- Odeyemi, O.A.; Burke, C.M.; Bolch, C.C.J.; Stanley, R. Seafood spoilage microbiota and associated volatile organic compounds at different storage temperatures and packaging conditions. Int. J. Food Microbiol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Phan, N.T.; Kim, K.H.; Jeon, E.C.; Kim, U.H.; Sohn, J.R.; Pandey, S.K. Analysis of volatile organic compounds released during food decaying processes. Env. Monit Assess 2012, 184, 1683–1692. [Google Scholar] [CrossRef] [PubMed]
- Mace, S.; Cardinal, M.; Jaffres, E.; Cornet, J.; Lalanne, V.; Chevalier, F.; Serot, T.; Pilet, M.F.; Dousset, X.; Joffraud, J.J. Evaluation of the spoilage potential of bacteria isolated from spoiled cooked whole tropical shrimp (penaeus vannamei) stored under modified atmosphere packaging. Food Microbiol. 2014, 40, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Moreira, N.; Valente, L.M.; Castro-Cunha, M.; Cunha, L.M.; Guedes de Pinho, P. Effect of storage time and heat processing on the volatile profile of senegalese sole (solea senegalensis kaup, 1858) muscle. Food Chem. 2013, 138, 2365–2373. [Google Scholar] [CrossRef] [PubMed]
- Edirisinghe, R.K.B.; Graffham, A.J.; Taylor, S.J. Characterisation of the volatiles of yellowfin tuna (thunnus albacares) during storage by solid phase microextraction and GC–MS and their relationship to fish quality parameters. Int. J. Food Sci. Technol. 2007, 42, 1139–1147. [Google Scholar] [CrossRef]
- He, H.J.; Wu, D.; Sun, D.W. Rapid and non-destructive determination of drip loss and ph distribution in farmed atlantic salmon (salmo salar) fillets using visible and near-infrared (vis–nir) hyperspectral imaging. Food Chem. 2014, 156, 394–401. [Google Scholar] [CrossRef] [PubMed]
- Harnkarnsujarit, N.; Kawai, K.; Suzuki, T. Effects of freezing temperature and water activity on microstructure, color, and protein conformation of freeze-dried bluefin tuna (thunnus orientalis). Food Bioprocess Technol. 2014, 8, 916–925. [Google Scholar] [CrossRef]
- Saito, K.; Ahhmed, A.M.; Kawahara, S.; Sugimoto, Y.; Aoki, T.; Muguruma, M.T. Effects of humidity-stabilizing sheets on the quality of bigeye tuna meat (thunnus obesus) during refrigerated storage. Food Sci. Technol. Res. 2009, 15, 283–292. [Google Scholar] [CrossRef]
- Guizani, N.; Albusaidy, M.A.; Albelushi, I.M.; Mothershaw, A.; Rahman, M.S. The effect of storage temperature on histamine production and the freshness of yellowfin tuna (thunnus albacares). Food Res. Int. 2005, 38, 215–222. [Google Scholar] [CrossRef]
- Silva, C.C.G.; Ponte, D.J.B.d.; Enes Dapkevicius, M.L.N. Storage temperature effect on histamine formation in big eye tuna and skipjack. J. Food Sci. 1998, 63, 644–647. [Google Scholar] [CrossRef]
- Alfonzo, A.; Gaglio, R.; Francesca, N.; Barbera, M.; Saiano, F.; Santulli, A.; Matraxia, M.; Rallo, F.; Moschetti, G. Influence of salt of different origin on the microbiological characteristics, histamine generation and volatile profile of salted anchovies ( engraulis encrasicolus l.). Food Control 2018, 92, 301–311. [Google Scholar] [CrossRef]
- Zaman, M.Z.; Bakar, F.A.; Selamat, J.; Bakar, J.; Ang, S.S.; Chong, C.Y. Degradation of histamine by the halotolerant staphylococcus carnosus fs19 isolate obtained from fish sauce. Food Control 2014, 40, 58–63. [Google Scholar] [CrossRef]
- Li, Q.; Zhang, L.; Lu, H.; Song, S.; Luo, Y. Comparison of postmortem changes in atp-related compounds, protein degradation and endogenous enzyme activity of white muscle and dark muscle from common carp (cyprinus carpio) stored at 4 °C. LWT 2017, 78, 317–324. [Google Scholar] [CrossRef]
- Li, Q.; Zhang, L.; Luo, Y. Changes in microbial communities and quality attributes of white muscle and dark muscle from common carp (cyprinus carpio) during chilled and freeze-chilled storage. Food Microbiol. 2018, 73, 237–244. [Google Scholar] [CrossRef] [PubMed]
- Nychas, G.J.; Skandamis, P.N.; Tassou, C.C.; Koutsoumanis, K.P. Meat spoilage during distribution. Meat Sci. 2008, 78, 77–89. [Google Scholar] [CrossRef]
- Casaburi, A.; Piombino, P.; Nychas, G.J.; Villani, F.; Ercolini, D. Bacterial populations and the volatilome associated to meat spoilage. Food Microbiol. 2015, 45, 83–102. [Google Scholar] [CrossRef]
- Sun, J.; Wang, Q.J.; Huang, J.; Hou, Y.D.; Chen, Y.F.; Su, X.R. Influence of heating temperature on the development of volatile compounds in bigeye tuna meat (thunnus obesus) as assessed by e-nose and spme-gc/ms. Int. Food Res. J. 2013, 20, 3077–3083. [Google Scholar]
- Liu, S.; Li, X.; Zhou, X.; Zhang, X.; Ding, Y. Comparative study of basic characteristics of ordinary and dark muscle in skipjack tuna (katsuwonus pelamis). Food Sci. Biotechnol. 2014, 23, 1397–1404. [Google Scholar] [CrossRef]
- Parlapani, F.F.; Mallouchos, A.; Haroutounian, S.A.; Boziaris, I.S. Microbiological spoilage and investigation of volatile profile during storage of sea bream fillets under various conditions. Int. J. Food Microbiol. 2014, 189, 153–163. [Google Scholar] [CrossRef]
- Duflos, G.; Coin, V.M.; Cornu, M.; Antinelli, J.-F.; Malle, P. Determination of volatile compounds to characterize fish spoilage using headspace/mass spectrometry and solid-phase microextraction/gas chromatography/mass spectrometry. J. Sci. Food Agric. 2006, 86, 600–611. [Google Scholar] [CrossRef]
- Iglesias, J.; Medina, I. Solid-phase microextraction method for the determination of volatile compounds associated to oxidation of fish muscle. J. Chromatogr A 2008, 1192, 9–16. [Google Scholar] [CrossRef]
- Quirós, R.B.D.; López-Hernández, J.; González-Castro, M.J.; Cruz-García, C.D.L.; Simal-Lozano, J.J.E.F.R. Comparison of volatile components in raw and cooked green beans by gc-ms using dynamic headspace sampling and microwave desorption. Eur. Food Res. Technol. 2000, 210, 226–230. [Google Scholar]
- Carneiro, C.d.S.; Mársico, E.T.; Ribeiro, R.d.O.R.; Conte-Júnior, C.A.; Mano, S.B.; Augusto, C.J.C.; Oliveira de Jesus, E.F. Low-field nuclear magnetic resonance (lf nmr 1h) to assess the mobility of water during storage of salted fish (sardinella brasiliensis). J. Food Eng. 2016, 169, 321–325. [Google Scholar] [CrossRef]
- Alhabsi, N.A.; Alhadhrami, S.; Alkasbi, H.; Rahman, M.S. Molecular mobility of fish flesh measured by low-field nuclear magnetic resonance (lf-nmr) relaxation: Effects of freeze–thaw cycles. Food Sci. Technol. 2017, 83. [Google Scholar]
- Tan, M.; Lin, Z.; Zu, Y.; Zhu, B.; Cheng, S. Effect of multiple freeze-thaw cycles on the quality of instant sea cucumber: Emphatically on water status of by lf-nmr and mri. Food Res. Int. 2018, 109, 65. [Google Scholar] [CrossRef] [PubMed]
- Kaale, L.D.; Eikevik, T.M. A study of the ice crystal sizes of red muscle of pre-rigor atlantic salmon (salmo salar) fillets during superchilled storage. J. Food Eng. 2013, 119, 544–551. [Google Scholar] [CrossRef]
- Shi, L.; Yang, T.; Xiong, G.; Li, X.; Wang, X.; Ding, A.; Qiao, Y.; Wu, W.; Liao, L.; Wang, L. Influence of frozen storage temperature on the microstructures and physicochemical properties of pre-frozen perch (micropterus salmoides). LWT 2018, 92, 471–476. [Google Scholar] [CrossRef]
- Li, D.; Zhang, J.; Song, S.; Feng, L.; Luo, Y. Influence of heat processing on the volatile organic compounds and microbial diversity of salted and vacuum-packaged silver carp (hypophthalmichthys molitrix) fillets during storage. Food Microbiol. 2018, 72, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Qian, Y.-F.; Xie, J.; Yang, S.-P.; Wu, W.-H. Study of the quality changes and myofibrillar proteins of white shrimp (litopenaeus vannamei) under modified atmosphere packaging with varying co2 levels. Eur. Food Res. Technol. 2013, 236, 629–635. [Google Scholar] [CrossRef]
- Köse, S.; Hall, G. Modification of a colorimetric method for histamine analysis in fish meal. Food Res. Int. 2000, 33, 839–845. [Google Scholar] [CrossRef]
- Chen, H.-C.; Huang, Y.-R.; Hsu, H.-H.; Lin, C.-S.; Chen, W.-C.; Lin, C.-M.; Tsai, Y.-H. Determination of histamine and biogenic amines in fish cubes (tetrapturus angustirostris) implicated in a food-borne poisoning. Food Control 2010, 21, 13–18. [Google Scholar] [CrossRef]
- Liu, X.; Chen, N.; Gao, X.; Zhang, Y.; Li, X.; Zhang, Y.; Bing, X.; Huang, H.; Zhang, X. The infection of red seabream iridovirus in mandarin fish (siniperca chuatsi) and the host immune related gene expression profiles. Fish Shellfish Immunol. 2018, 74, 474–484. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Not available. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| VOCs | Relative Concentration (Area 10−6) | ||||
|---|---|---|---|---|---|
| Day 0 | Day 3–0 | Day 6–0 | Day 3–4 | Day 6–4 | |
| Aldehydes | |||||
| Hexanal | 8.61 ± 0.12 d | 10.63 ± 0.64 d | 31.39 ± 0.69 b | 23.72 ± 0.02 c | 35.56 ± 2.5 a |
| Heptanal | 5.74 ± 0.12 b | 1.12 ± 0.05 c | 0.51 ± 0.14 c,d | 0.31 ± 0.00 c,d | 28.87 ± 0.38 a |
| Octanal | 1.35 ± 0.01 b,c | 3.83 ± 0.01 b | 2.33 ± 0.06 b,c | 0.02 ± 0.00 c,d | 18.04 ± 0.27 a |
| Nonanal | 1.21 ± 0.04 c | 9.98 ± 0.10 b | 1.08 ± 0.03 c | 0.02 ± 0.00 d | 17.62 ± 0.31 a |
| 2,6-Nonadienal,(E,Z)- | - | 0.01 ± 0.00 b | - | 0.02 ± 0.00 b | 6.54 ± 0.04 a |
| Tetradecanal | - | 0.01 ± 0.00 b | 0.01 ± 0.00 b | 0.02 ± 0.00 b | 2.95 ± 0.05 a |
| Propanal | 0.79 ± 0.01 e | 10.05 ± 0.12 b | 4.96 ± 0.14 c | 2.83 ± 0.02 d | 20.90 ± 0.98 a |
| 4-Heptenal, (Z)- | - | 0.29 ± 0.01 b | 0.28 ± 0.01 b | 0.02 ± 0.00 c | 5.67 ± 0.05 a |
| Pentadecanal- | - | 0.01 ± 0.00 c | 0.50 ± 0.02 b | 0.02 ± 0.00 c | 16.18 ± 0.20 a |
| ketones | |||||
| Acetone | 0.04 ± 0.01 c | 0.29 ± 0.01 b | 0.58 ± 0.01 b | 5.23 ± 0.36 a | 0.13 ± 0.00 b |
| 2-Heptanone | - | 0.05 ± 0.00 b | - | 0.08 ± 0.06 b | 0.14 ± 0.06 a |
| 2-Nonanone | - | 0.04 ± 0.01 b | 0.06 ± 0.01 b | 0.09 ± 0.03 b | 0.76 ± 0.04 a |
| 2-Undecanone | - | - | 0.93 ± 0.26 a | - | 0.56 ± 0.06 b |
| 2,3-Pentanedione | 0.05 ± 0.00 a | 0.04 ± 0.00 a | 0.08 ± 0.01 a | 0.09 ± 0.00 a | 0.02 ± 0.00 a |
| Alcohols | |||||
| 1-Pentanol | 0.96 ± 0.18 d | 11.53 ± 0.03 c | 10.30 ± 0.09 c | 23.37 ± 0.00 b | 69.16 ± 2.70 a |
| 1-Hexanol | 4.03 ± 1.51 e | 20.74 ± 0.05 c | 83.40 ± 0.93 a | 10.56 ± 0.04 d | 69.68 ± 50.97 b |
| 1-Octen-3-ol | 7.37 ± 2.15 d | 77.27 ± 0.27 b | 88.0 ± 0.93 a | 61.41 ± 0.04 c | 65.08 ± 16.45 c |
| 1-Heptanol | 1.99 ± 0.74 e | 14.20 ± 0.04 d | 23.48 ± 0.25 c | 56.02 ± 0.03 b | 142.15 ± 4.87 a |
| 1-Octanol | 7.88 ± 5.21 c | 5.55 ± 0.05 e | 6.24 ± 0.05 d | 12.88 ± 0.04 b | 39.19 ± 4.41 a |
| 2-Octen-1-ol, (E)- | - | 3.08 ± 0.02 b | 3.90 ± 0.03 b | 0.04 ± 0.00 c | 47.04 ± 3.52 a |
| 1-Nonanol | 0.82 ± 0.30 d | 2.52 ± 0.01 c | 3.36 ± 0.03 c | 76.97 ± 7.44 a | 16.09 ± 0.99 b |
| 2-Penten-1-ol, (Z)- | - | 0.72 ± 0.01 c | 1.74 ± 0.03 c | 9.55 ± 0.00 b | 55.07 ± 5.35 a |
| 1-Penten-3-ol | 10.93 ± 4.95 d | 161.64 ± 0.39 b | 120.12 ± 1.19 c | 129.63 ± 0.20 c | 138.87 ± 2.84 a |
| Others | |||||
| Nonanoic acid, ethyl ester | 0.90 ± 0.79 b | 0.02 ± 0.00 d | 0.75 ± 0.13 c | 0.02 ± 0.00 d | 2.63 ± 0.22 a |
| Hexanoic acid | - | 1.64 ± 0.28 c | 1.76 ± 0.03 c | 2.71 ± 0.04 b | 9.73 ± 0.00 a |
| Benzaldehyde | 0.05 ± 0.02 b | 0.23 ± 0.06 a | 0.06 ± 0.01 b | 0.10 ± 0.04 a | 0.21 ± 0.03 a |
| Naphthalene | 0.05 ± 0.00 b | 0.13 ± 0.02 a | 0.08 ± 0.02 a,b | 0.10 ± 0.08 a | 0.02 ± 0.00 b |
| Naphthalene,2 methyl- | 0.03 ± 0.01 b | 0.04 ± 0.01 b | 0.02 ± 0.00b b | 0.20 ± 0.00 b | 0.57 ± 0.25 a |
| Parameter | T2b | T21 | T22 |
|---|---|---|---|
| Hexanal | −0.25 | −0.80 ** | 0.39 |
| Heptanal | −0.18 | −0.21 | 0.48 |
| Octanal | −0.17 | −0.40 | 0.41 |
| Nonanal | −0.02 | −0.34 | 0.18 |
| 2,6-Nonadienal,(E,Z)- | −0.14 | −0.38 | 0.43 |
| Tetradecanal | −0.14 | −0.38 | 0.43 |
| Propanal | −0.06 | −0.52 * | 0.23 |
| 4-Heptenal, (Z)- | −0.16 | −0.40 | 0.44 |
| Pentadecanal- | −0.16 | −0.40 | 0.44 |
| Acetone | 0.53 * | −0.21 | −0.46 |
| 2-Heptanone | 0.22 | −0.46 | −0.01 |
| 2-Nonanone | −0.11 | −0.47 | 0.39 |
| 2-Undecanone | −0.60 * | −0.62 * | 0.56 * |
| 2,3-Pentanedione | 0.05 | −0.15 | −0.21 |
| 1-Pentanol | 0.02 | −0.54 * | 0.28 |
| 1-Hexanol | −0.54 * | −0.71 ** | 0.51 |
| 1-Octen−3-ol | 0.01 | 0.88 ** | −0.16 |
| 1-Heptanol | 0.03 | −0.56 * | 0.28 |
| 1-Octanol | −0.04 | −0.38 | 0.37 |
| 2-Octen-1-ol, (E)- | −0.17 | −0.42 | 0.43 |
| 1-Nonanol | 0.55 * | −0.25 | −0.39 |
| 2-Penten-1-ol, (Z)- | −0.05 | −0.43 | 0.36 |
| 1-Penten-3-ol | 0.27 | −0.82 ** | −0.33 |
| Nonanoic acid, ethyl ester | −0.40 | −0.21 | 0.63 * |
| Hexanoic acid | −0.03 | −0.56 * | 0.31 |
| Benzaldehyde | 0.29 | 0.44 | −0.27 |
| Naphthalene | 0.37 | −0.06 | −0.65 ** |
| Naphthalene,2 methyl- | 0.05 | 0.43 | 0.28 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.-Y.; Xie, J. Study on the Volatile Organic Compounds and Its Correlation with Water Dynamics of Bigeye Tuna (Thunnus obesus) during Cold Storage. Molecules 2019, 24, 3119. https://doi.org/10.3390/molecules24173119
Wang X-Y, Xie J. Study on the Volatile Organic Compounds and Its Correlation with Water Dynamics of Bigeye Tuna (Thunnus obesus) during Cold Storage. Molecules. 2019; 24(17):3119. https://doi.org/10.3390/molecules24173119
Chicago/Turabian StyleWang, Xin-Yun, and Jing Xie. 2019. "Study on the Volatile Organic Compounds and Its Correlation with Water Dynamics of Bigeye Tuna (Thunnus obesus) during Cold Storage" Molecules 24, no. 17: 3119. https://doi.org/10.3390/molecules24173119
APA StyleWang, X.-Y., & Xie, J. (2019). Study on the Volatile Organic Compounds and Its Correlation with Water Dynamics of Bigeye Tuna (Thunnus obesus) during Cold Storage. Molecules, 24(17), 3119. https://doi.org/10.3390/molecules24173119