
Metabolite Transformation and Enzyme Activities of Hainan Vanilla Beans During Curing to Improve Flavor Formation


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Change of the Metabolites
2.2. Principal Component Analysis (PCA)
2.3. Change of the Main Vanilla Flavors
2.3.1. Change of Vanillin
2.3.2. Change of Vanillic Acid
2.3.3. Change of p-hydroxybenzaldehyde
2.3.4. Change of p-hydroxybenzoic Acid
2.4. Change of Vanillin Precursors
2.4.1. Change of Glucovanillin
2.4.2. Change of Ferulic Acid
2.4.3. Change of p-coumaric Acid
2.4.4. Change of Vanillyl Alcohol
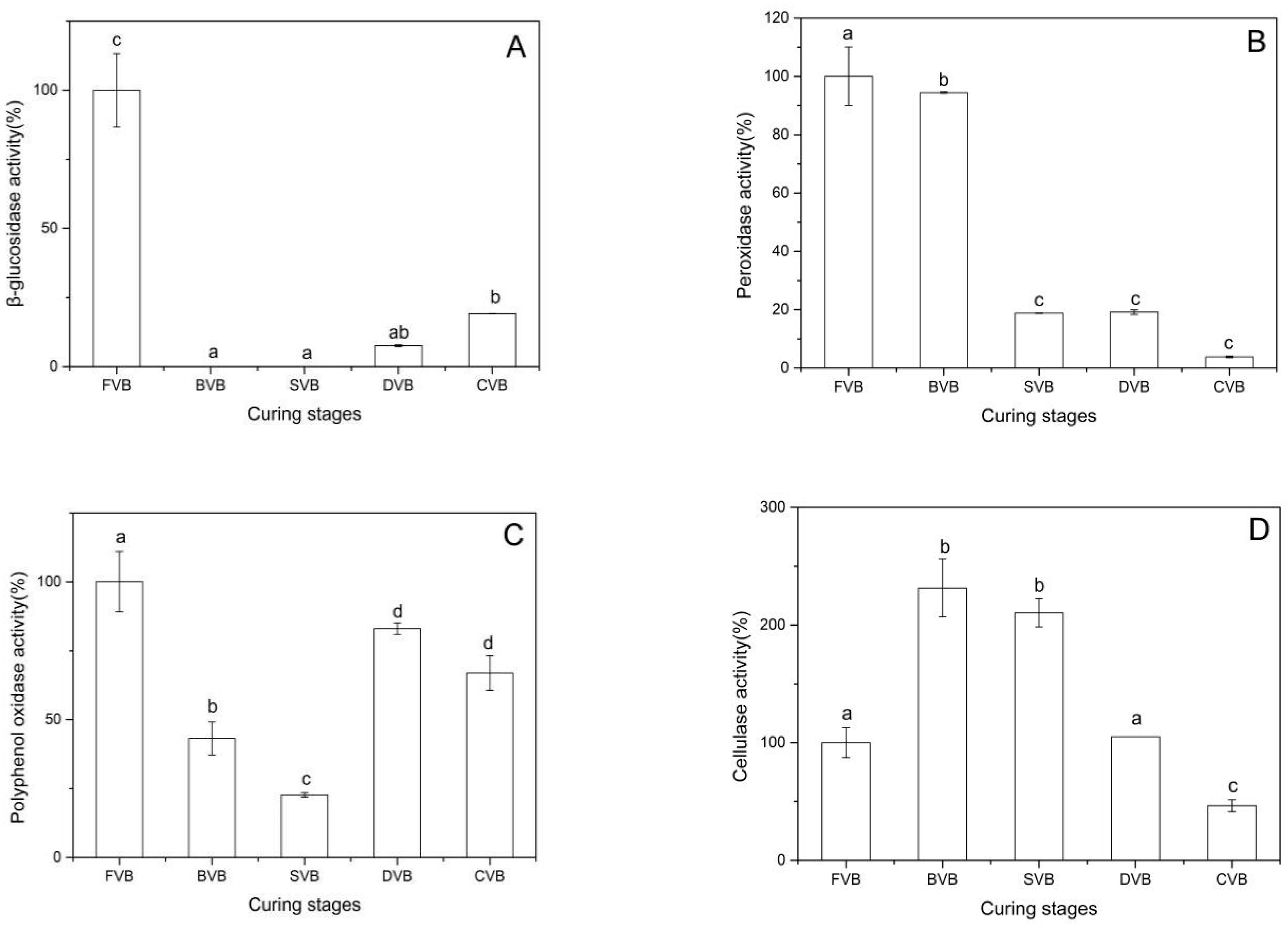
2.5. Enzymatic Activities in Vanilla Beans
2.5.1. Change of β-Glucosidase
2.5.2. Change of Peroxidase
2.5.3. Change of Polyphenol Oxidase
2.5.4. Change of Cellulase
3. Materials and Methods
3.1. Materials
3.2. Curing Process and Sampling
3.3. Sample Preparation and Extraction
3.4. HPLC Conditions
3.5. ESI-Q TRAP-MS/MS
3.6. Metabolite Identification and Quantification
3.7. Determination of the Main Vanilla Flavors
3.8. Determination of Vanillin Precursors
3.9. Determination of the Enzymatic Activities
3.10. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Ramachandra Rao, S.; Ravishankar, G.A. Vanilla flavour: Production by conventional and biotechnological routes. J. Sci. Food Agric. 2000, 80, 289–304. [Google Scholar] [CrossRef]
- Naidu, M.M.; Kumar, P.V.S.; Shyamala, B.N.; Sulochanamma, G.; Prakash, M.; Thakur, M.S. Enzyme-Assisted Process for Production of Superior Quality Vanilla Extracts from Green Vanilla Pods Using Tea Leaf Enzymes. Food Bioprocess Technol. 2012, 5, 527–532. [Google Scholar] [CrossRef]
- Hassan, S.; Araceli, P.S.; Denis, B.; Maria de los Angeles, V.-V.; Mayra, N.-G.; Delfino, R.-L. Identification of volatile compounds in cured Mexican vanilla (Vanilla planifolia G. Jackson) beans using headspace solid-phase microextraction with gas chromatography-mass spectrometry. Fruits 2016, 71, 407–418. [Google Scholar] [CrossRef][Green Version]
- Sreedhar, R.V.; Roohie, K.; Maya, P.; Venkatachalam, L.; Bhagyalakshmi, N. Biotic elicitors enhance flavour compounds during accelerated curing of vanilla beans. Food Chem. 2009, 112, 461–468. [Google Scholar] [CrossRef]
- Pérez Silva, A.; Gunata, Z.; Lepoutre, J.P.; Odoux, E. New insight on the genesis and fate of odor-active compounds in vanilla beans (Vanilla planifolia G. Jackson) during traditional curing. Food Res. Int. 2011, 44, 2930–2937. [Google Scholar] [CrossRef]
- Walton, N.J.; Mayer, M.J.; Narbad, A. Vanillin. Phytochemistry 2003, 63, 505–515. [Google Scholar] [CrossRef]
- Dong, Z.; Gu, F.; Xu, F.; Wang, Q. Comparison of four kinds of extraction techniques and kinetics of microwave-assisted extraction of vanillin from Vanilla planifolia Andrews. Food Chem. 2014, 149, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Funk, C.; Brodelius, P.E. Phenylpropanoid metabolism in suspension-cultures of Vanilla planifolia Andr. II Effects of precursor feeding and metabolic-inhibitors. Plant Physiol. 1990, 94, 95–101. [Google Scholar] [CrossRef]
- Funk, C.; Brodelius, P.E. Phenylpropanoid metabolism in suspension-cultures of Vanilla planifolia Andr. III conversion of 4-methoxycinnamic acids into 4-hydroxybenzoic acids. Plant Physiol. 1990, 94, 102–108. [Google Scholar] [CrossRef]
- Funk, C.; Brodelius, P.E. Phenylpropanoid metabolism in suspension-cultures of Vanilla planifolia Andr. IV Induction of vanillic acid formation. Plant Physiol. 1992, 99, 256–262. [Google Scholar] [CrossRef]
- Gallage, N.J.; Hansen, E.H.; Kannangara, R.; Olsen, C.E.; Motawia, M.S.; Jorgensen, K.; Holme, I.; Hebelstrup, K.; Grisoni, M.; Moller, B.L. Vanillin formation from ferulic acid in Vanilla planifolia is catalysed by a single enzyme. Nat. Commun. 2014, 5, 4037. [Google Scholar] [CrossRef] [PubMed]
- Kundu, A. Vanillin biosynthetic pathways in plants. Planta 2017, 245, 1069–1078. [Google Scholar] [CrossRef]
- Yang, H.; Barros-Rios, J.; Kourteva, G.; Rao, X.; Chen, F.; Shen, H.; Liu, C.; Podstolski, A.; Belanger, F.; Havkin-Frenkel, D.; et al. A re-evaluation of the final step of vanillin biosynthesis in the orchid Vanilla planifolia. Phytochemistry 2017, 139, 33–46. [Google Scholar] [CrossRef] [PubMed]
- Dixon, R.A. Vanillin biosynthesis—Not as simple as it seems? In Handbook of Vanilla Science and Technology; Havkin-Frenkel, D., Belanger, F.C., Eds.; WileyBlackwell: Oxford, UK, 2011; pp. 292–298. [Google Scholar]
- Mäkelä, M.R.; Marinović, M.; Nousiainen, P.; Liwanag, A.J.; Benoit, I.; Sipilä, J.; Hatakka, A.; de Vries, R.P.; Hildén, K.S. Aromatic metabolism of filamentous fungi in relation to the presence of aromatic compounds in plant biomass. Adv. Appl. Microbiol. 2015, 91, 63–137. [Google Scholar] [PubMed]
- Dignum, M.J.W.; Kerler, J.; Verpoorte, R. Vanilla production: Technological, chemical and biosynthetic aspects. Food Rev. Int. 2001, 17, 199–219. [Google Scholar] [CrossRef]
- Zenk, M.H. Biosynthese von vanillin in Vanilla planifolia Andr. Z. Pflanzenphysiol. 1965, 53, 404–414. [Google Scholar]
- French, C.J.; Vance, C.P.; Towers, G.H.N. Conversion of para-coumaric acid to para-hydroxybenzoic acid by cell free extracts of potato-tubers and polyporus-hispidus. Phytochemistry 1976, 15, 564–566. [Google Scholar] [CrossRef]
- Dignum, M.J.W.; van der Heijden, R.; Kerler, J.; Winkel, C.; Verpoorte, R. Identification of glucosides in green beans of Vanilla planifolia Andrews and kinetics of vanilla beta-glucosidase. Food Chem. 2004, 85, 199–205. [Google Scholar] [CrossRef]
- Kaur, B.; Chakraborty, D. Biotechnological and Molecular Approaches for Vanillin Production: A Review. Appl. Biochem. Biotechnol. 2013, 169, 1353–1372. [Google Scholar] [CrossRef]
- Gu, F.; Chen, Y.; Hong, Y.; Fang, Y.; Tan, L. Comparative metabolomics in vanilla pod and vanilla bean revealing the biosynthesis of vanillin during the curing process of vanilla. Amb Express 2017, 7, 116. [Google Scholar] [CrossRef]
- Sreedhar, R.V.; Roohie, K.; Venkatachalam, L.; Narayan, M.S.; Bhagyalakshmi, N. Specific pretreatments reduce curing period of vanilla (Vanilla planifolia) beans. J. Agric. Food Chem. 2007, 55, 2947–2955. [Google Scholar] [CrossRef] [PubMed]
- Röling, W.F.; Kerler, J.; Braster, M.; Apriyantono, A.; Stam, H.; van Verseveld, H.W. Microorganisms with a taste for vanilla: Microbial ecology of traditional Indonesian vanilla curing. Appl. Environ. Microbiol. 2001, 67, 1995–2003. [Google Scholar] [CrossRef] [PubMed]
- General, T.; Mamatha, V.; Divya, V.; Appaiah, K.A.A. Diversity of yeast with β-glycosidase activity in vanilla (Vanilla planifolio) plant. Curr. Sci. 2009, 96, 1501–1505. [Google Scholar]
- Chen, Y.; Gu, F.; Li, J.; Xu, F.; He, S.; Fang, Y. Involvement of Colonizing Bacillus Isolates in Glucovanillin Hydrolysis during the Curing of Vanilla planifolia Andrews. Appl. Environ. Microbiol. 2015, 81, 4947–4954. [Google Scholar] [CrossRef] [PubMed]
- Dignum, M.J.W.; Kerler, J.; Verpoorte, R. Beta-glucosidase and peroxidase stability in crude enzyme extracts from green beans of Vanilla planifolia Andrews. Phytochem. Anal. 2001, 12, 174–179. [Google Scholar] [CrossRef] [PubMed]
- Dunn, W.B.; Ellis, D.I. Metabolomics: Current analytical platforms and methodologies. TrAC Trends Anal. Chem. 2005, 24, 285–294. [Google Scholar]
- Lee, J.-E.; Hwang, G.-S.; Lee, C.-H.; Hong, Y.-S. Metabolomics Reveals Alterations in Both Primary and Secondary Metabolites by Wine Bacteria. J. Agric. Food Chem. 2009, 57, 10772–10783. [Google Scholar] [CrossRef] [PubMed]
- Schmidtke, L.M.; Blackman, J.W.; Clark, A.C.; Grant-Preece, P. Wine Metabolomics: Objective Measures of Sensory Properties of Semillon from GC-MS Profiles. J. Agric. Food Chem. 2013, 61, 11957–11967. [Google Scholar] [CrossRef]
- Xu, F.; Zou, L.; Ong, C.N. Experiment-originated variations, and multi-peak and multi-origination phenomena in derivatization-based GC-MS metabolomics. TrAC Trends Anal. Chem. 2010, 29, 269–280. [Google Scholar] [CrossRef]
- Chen, L.; Zhou, L.; Chan, E.C.; Neo, J.; Beuerman, R.W. Characterization of the human tear metabolome by LC-MS/MS. J Proteome Res 2011, 10, 4876–4882. [Google Scholar] [CrossRef]
- Palama, T.L. Metabolic Changes in Different Developmental Stages of Vanilla planifolia Pods. J. Agric. Food Chem. 2009, 17, 7651–7658. [Google Scholar] [CrossRef] [PubMed]
- Frenkel, C.; Ranadive, A.S.; Vázquez, J.T.; Havkin-Frenkel, D. Curing of vanilla. In Handbook of Vanilla Science and Technology; Havkin-Frenkel, D., Belanger, F.C., Eds.; WileyBlackwell: Oxford, UK, 2011; pp. 79–103. [Google Scholar]
- Dignum, M.J.W.; Kerler, J.; Verpoorte, R. Vanilla curing under laboratory conditions. Food Chem. 2002, 79, 165–171. [Google Scholar] [CrossRef]
- Pardio, V.T.; Mariezcurrena, M.D.; Waliszewski, K.N.; Sánchez, V.; Janczur, M.K. Effects of killing conditions of vanilla (Vanilla planifolia, Andrews) pods during the curing process on aroma composition of pod ethanol extract. Int. J. Food Sci. Technol. 2009, 44, 2417–2423. [Google Scholar] [CrossRef]
- Wang, D.; Zhang, L.; Huang, X.; Wang, X.; Yang, R.; Mao, J.; Wang, X.; Wang, X.; Zhang, Q.; Li, P. Identification of Nutritional Components in Black Sesame Determined by Widely Targeted Metabolomics and Traditional Chinese Medicines. Molecules 2018, 23, 1180. [Google Scholar] [CrossRef] [PubMed]
- Yoshio, T.; Tetsuya, M.; Ryo, N.; Makoto, K.; Kazuki, S.; Seiichi, O.; Ning, W.; Miyako, K. Metabolomic Evaluation of the Quality of Leaf Lettuce Grown in Practical Plant Factory to Capture Metabolite Signature. Front. Plant Sci. 2018, 9, 665. [Google Scholar]
- Hansen, E.H.; Møller, B.L.; Kock, G.R.; Bünner, C.M.; Kristensen, C.; Jensen, O.R.; Okkels, F.T.; Olsen, C.E.; Motawia, M.S.; Hansen, J. De novo biosynthesis of vanillin in fission yeast (Schizosaccharomyces pombe) and baker’s yeast (Saccharomyces cerevisiae). Appl. Environ. Microbiol. 2009, 75, 2765–2774. [Google Scholar] [CrossRef] [PubMed]
- Havkin-Frenkel, D.; French, J.C.; Pak, F.; Frenkel, C. Inside vanilla: Vanilla planifolia’s botany, curing options and future market prospects. Perfum. Flavor. 2005, 30, 36–55. [Google Scholar]
- Ranadive, A.S. Vanillin and related flavor compounds in vanilla extracts made from beans of various global origins. J. Agric. Food Chem. 1992, 40, 1922–1924. [Google Scholar] [CrossRef]
- Odoux, E. Vanilla curing. In Vanilla; Odoux, E., Grisoni, M., Eds.; CRC: Boca Raton, FL, USA, 2011; Volume 11, pp. 173–185. [Google Scholar]
- Odoux, E. Glucosylated aroma precursors and glucosidase(s) in vanilla bean (Vanilla planifolia G. Jackson). Fruits 2006, 61, 171–184. [Google Scholar] [CrossRef][Green Version]
- Li, X.; Yang, J.; Li, X.; Gu, W.; Huang, J.; Zhang, K.-Q. The metabolism of ferulic acid via 4-vinylguaiacol to vanillin by Enterobacter sp. Px6-4 isolated from Vanilla root. Process. Biochem. 2008, 43, 1132–1137. [Google Scholar] [CrossRef]
- Gallage, N.; Møller, B.L. Vanillin–bioconversion and bioengineering of the most popular plant flavor and its de novo biosynthesis in the vanilla orchid. Mol. Plant 2015, 8, 40–57. [Google Scholar] [CrossRef] [PubMed]
- Podstolski, A.; Havkin-Frenkel, D.; Malinowski, J.; Blount, J.W.; Kourteva, G.; Dixon, R.A. Unusual 4-hydroxybenzaldehyde synthase activity from tissue cultures of the vanilla orchid Vanilla planifolia. Phytochemistry 2002, 61, 611–620. [Google Scholar] [CrossRef]
- Brillouet, J.M.; Odoux, É. In vivo kinetics of β-glucosidase towards glucovanillin and related phenolic glucosides in heat-treated vanilla pod (Vanilla planifolia, Orchidaceae). Fruits 2010, 65, 85–95. [Google Scholar] [CrossRef][Green Version]
- Brillouet, J.-M.; Odoux, E.; Conejero, G. A set of data on green, ripening and senescent vanilla pod (Vanilla planifolia; Orchidaceae): Anatomy, enzymes, phenolics and lipids. Fruits 2010, 65, 221–235. [Google Scholar] [CrossRef]
- Waliszewski, K.N.; Marquez, O.; Pardio, V.T. Quantification and characterisation of polyphenol oxidase from vanilla bean. Food Chem. 2009, 117, 196–203. [Google Scholar] [CrossRef]
- Chen, W.; Gong, L.; Guo, Z.; Wang, W.; Zhang, H.; Liu, X.; Yu, S.; Xiong, L.; Luo, J. A Novel Integrated Method for Large-Scale Detection, Identification, and Quantification of Widely Targeted Metabolites: Application in the Study of Rice Metabolomics. Mol. Plant 2013, 6, 1769–1780. [Google Scholar] [CrossRef] [PubMed]
- Fraga, C.G.; Clowers, B.H.; Moore, R.J.; Zink, E.M. Signature-discovery approach for sample matching of a nerve-agent precursor using liquid chromatography-mass spectrometry, XCMS, and chemometrics. Anal. Chem. 2010, 82, 4165–4173. [Google Scholar] [CrossRef] [PubMed]
- Sinha, A.K.; Sharma, U.K.; Sharma, N. A comprehensive review on vanilla flavor: Extraction, isolation and quantification of vanillin and others constituents. Int. J. Food Sci. Nutr. 2008, 59, 299–326. [Google Scholar] [CrossRef] [PubMed]
- Sinha, A.K.; Verma, S.C.; Sharma, U.K. Development and validation of an RP-HPLC method for quantitative determination of vanillin and related phenolic compounds in Vanilla planifolia. J. Sep. Sci. 2007, 30, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Malik, C.P.; Singh, M.B. Plant Enzynoligy and Histoenzynology; Kalyani Publishers: New Delhi, India, 1980; pp. 53–72. [Google Scholar]
- Xia, J.; Sinelnikov, I.V.; Han, B.; Wishart, D.S. MetaboAnalyst 3.0-making metabolomics more meaningful. Nucleic Acids Res. 2015, 43, W251–W257. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are available from the authors. |






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cai, Y.; Gu, F.; Hong, Y.; Chen, Y.; Xu, F.; An, K. Metabolite Transformation and Enzyme Activities of Hainan Vanilla Beans During Curing to Improve Flavor Formation. Molecules 2019, 24, 2781. https://doi.org/10.3390/molecules24152781
Cai Y, Gu F, Hong Y, Chen Y, Xu F, An K. Metabolite Transformation and Enzyme Activities of Hainan Vanilla Beans During Curing to Improve Flavor Formation. Molecules. 2019; 24(15):2781. https://doi.org/10.3390/molecules24152781
Chicago/Turabian StyleCai, Yingying, Fenglin Gu, Yinghua Hong, Yonggan Chen, Fei Xu, and Kejing An. 2019. "Metabolite Transformation and Enzyme Activities of Hainan Vanilla Beans During Curing to Improve Flavor Formation" Molecules 24, no. 15: 2781. https://doi.org/10.3390/molecules24152781
APA StyleCai, Y., Gu, F., Hong, Y., Chen, Y., Xu, F., & An, K. (2019). Metabolite Transformation and Enzyme Activities of Hainan Vanilla Beans During Curing to Improve Flavor Formation. Molecules, 24(15), 2781. https://doi.org/10.3390/molecules24152781