Total Alkaloids from Bamboo Shoots and Bamboo Shoot Shells of Pleioblastus amarus (Keng) Keng f. and Their Anti-Inflammatory Activities

Abstract
1. Introduction
2. Results and Discussion
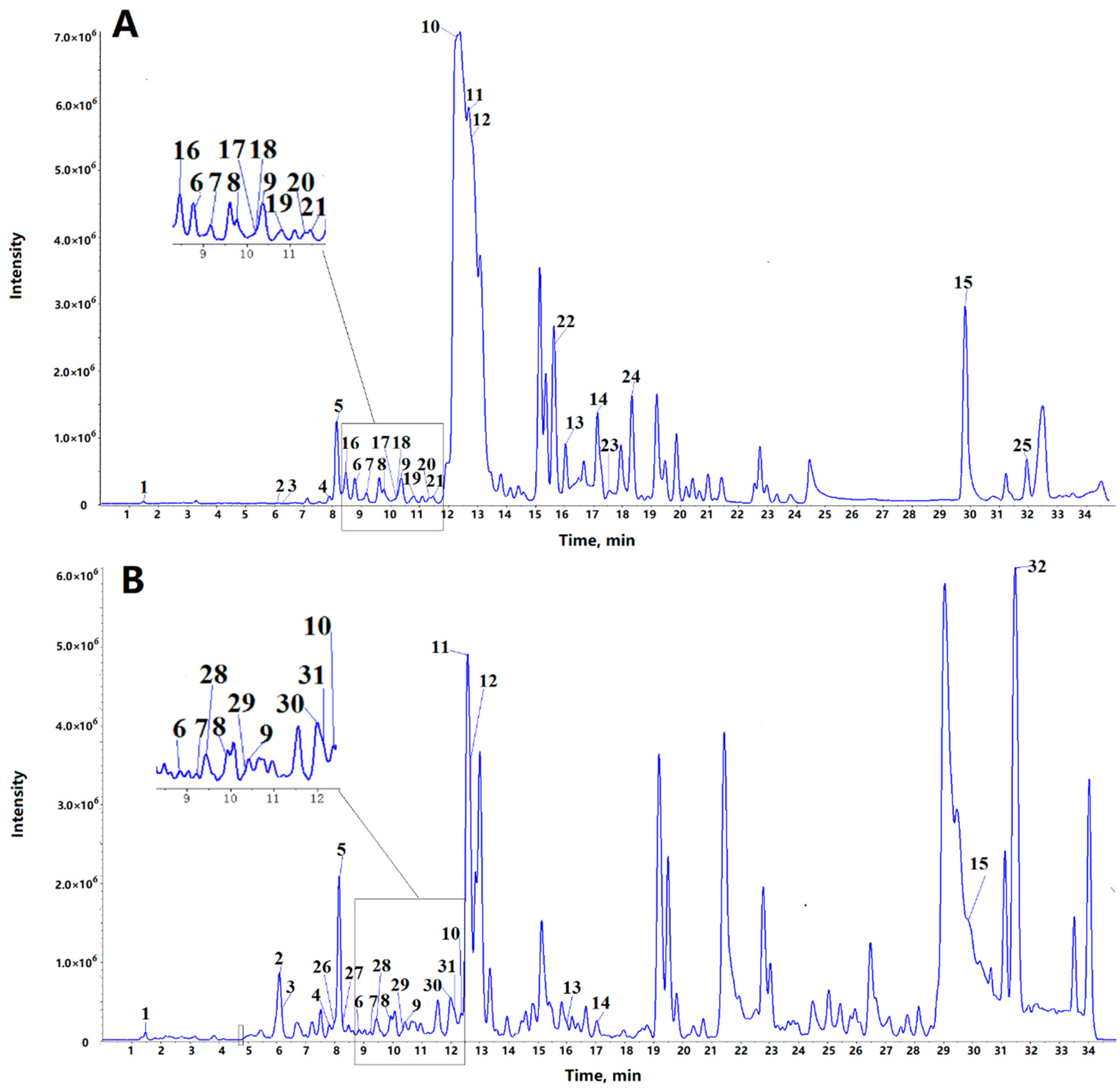
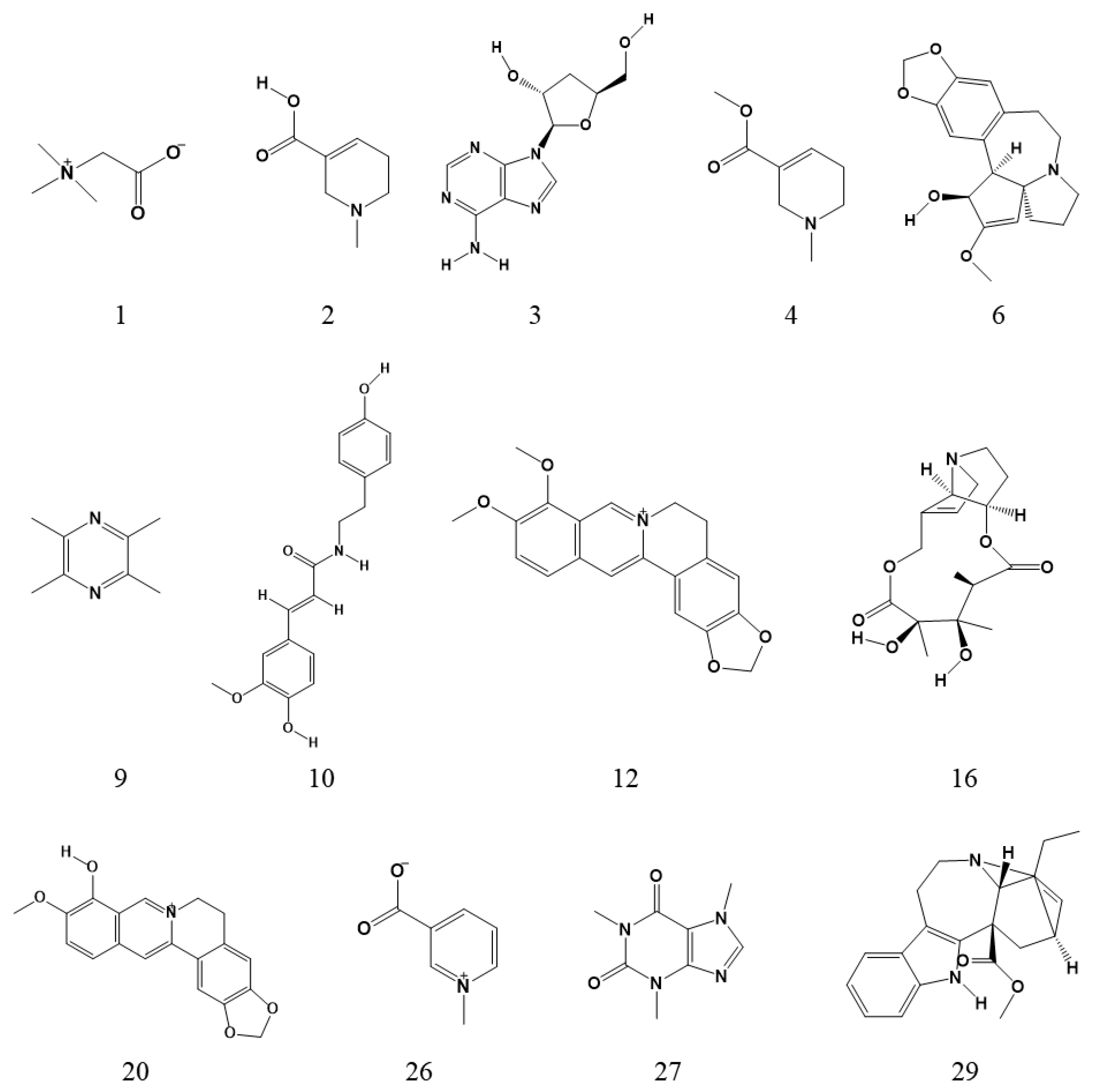
2.1. Identification of Alkaloids
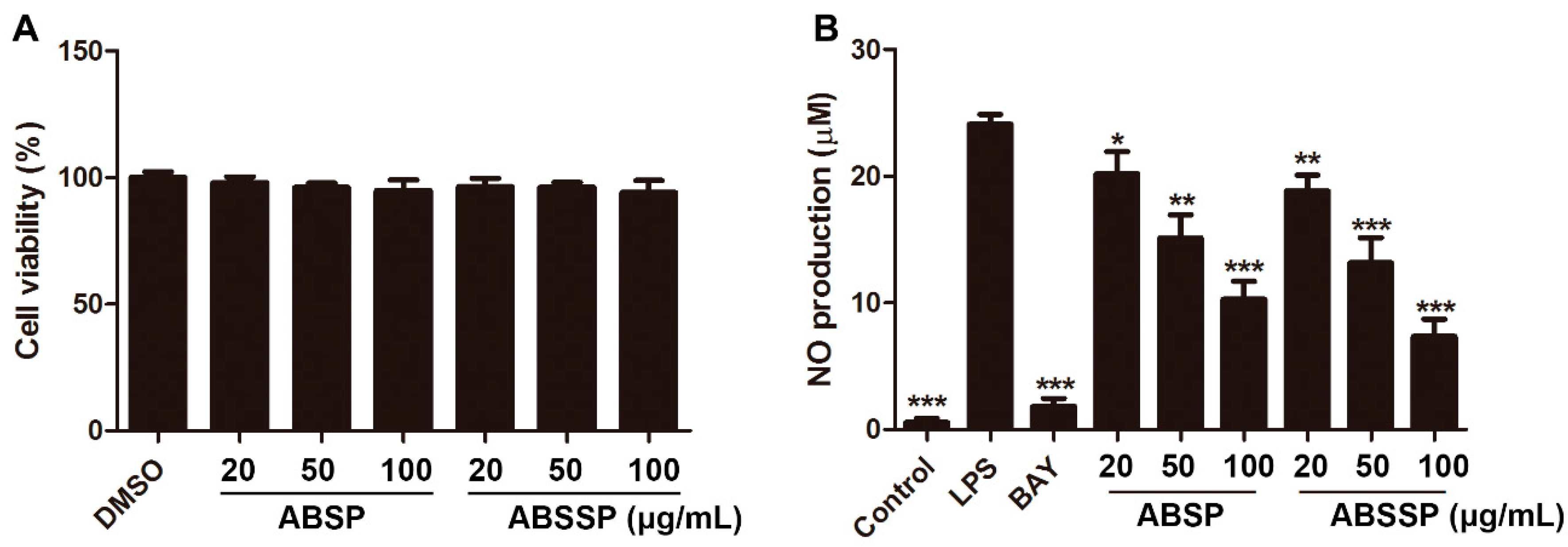
2.2. ABSP and ABSSP Inhibit LPS-Induced Nitric Oxide Production in RAW264.7 Cells
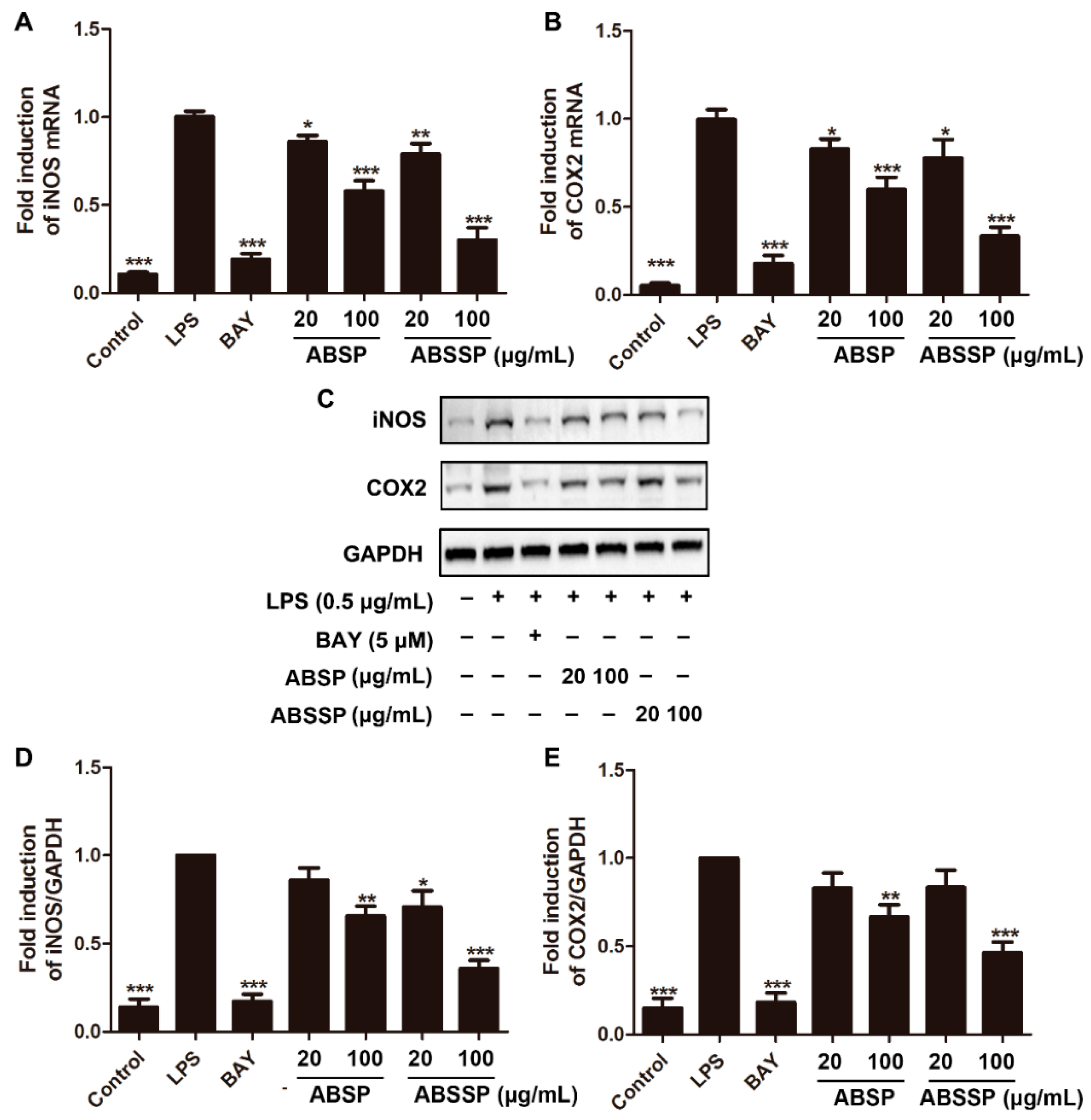
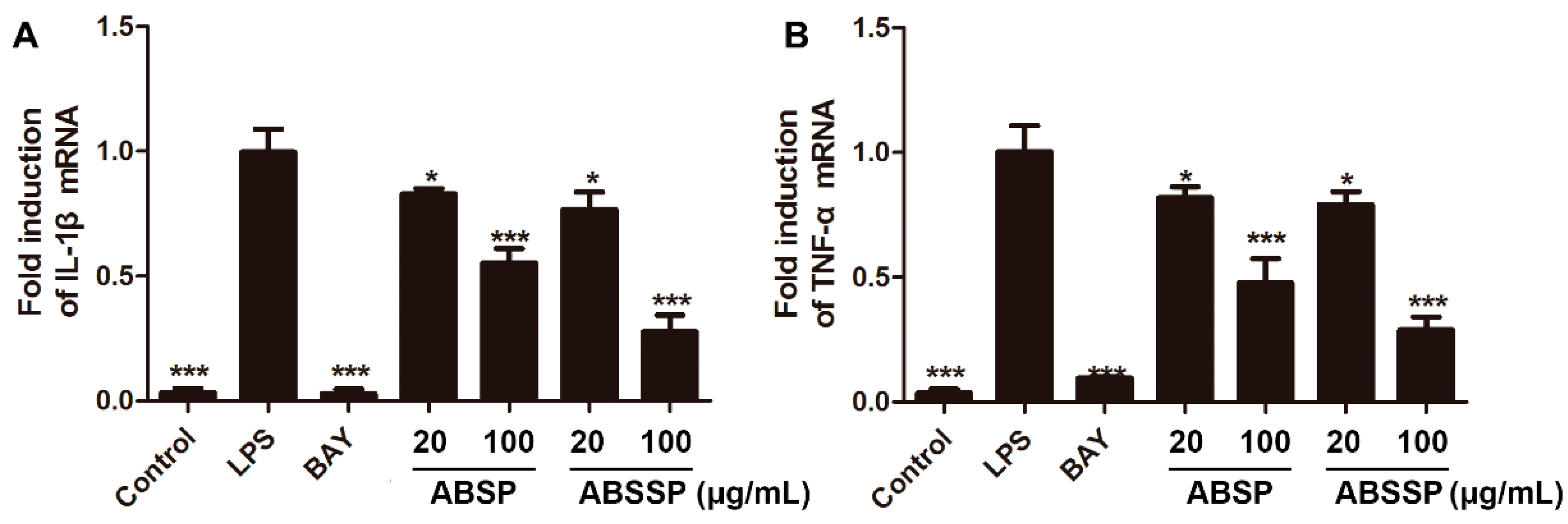
2.3. ABSP and ABSSP Suppress the mRNA Expression of Proinflammatory Cytokines in LPS-Exposed RAW264.7 Cells
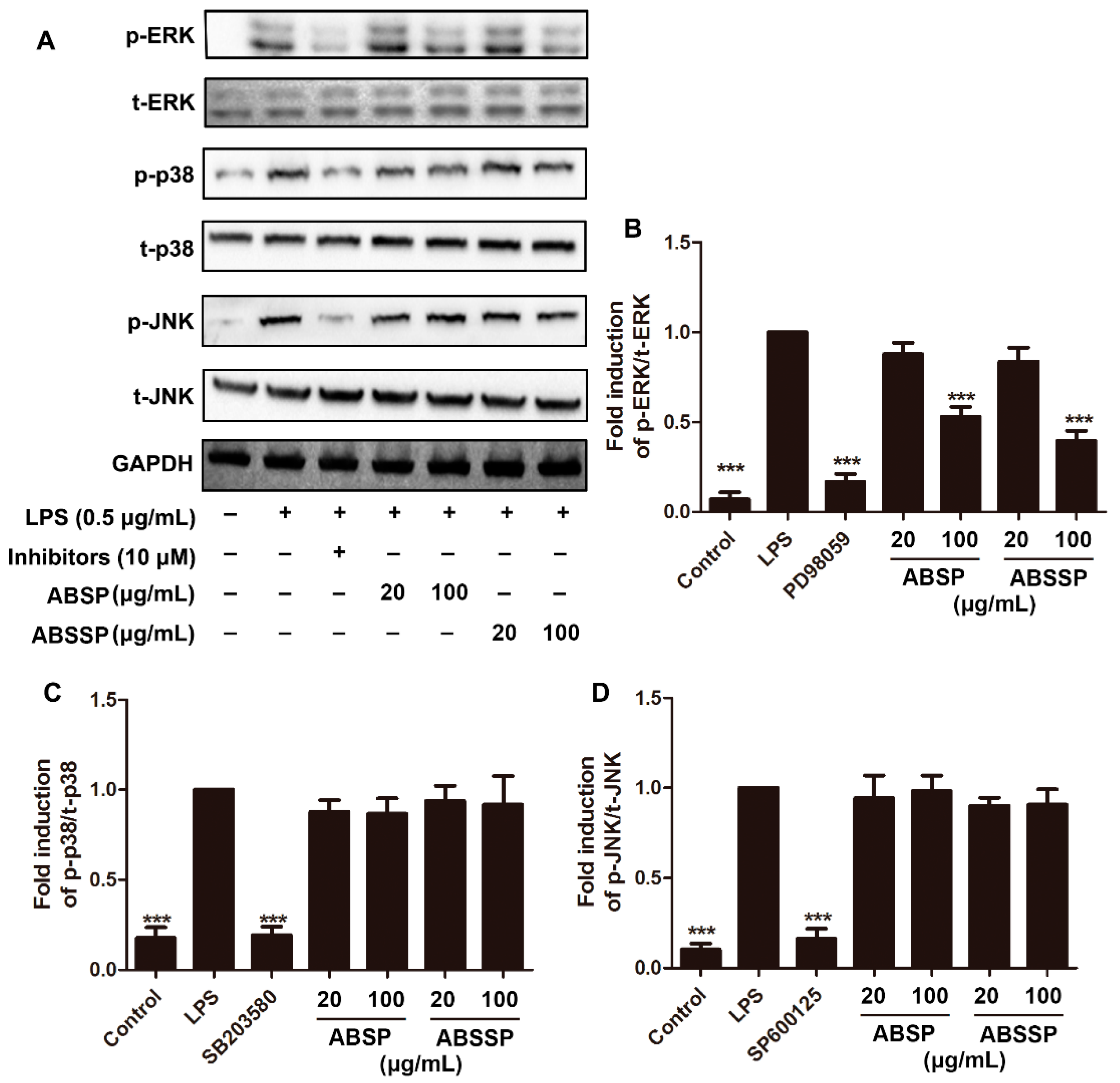
2.4. ABSP and ABSSP Block LPS-Induced ERK Activation in RAW264.7 Cells
3. Materials and Methods
3.1. Plant Materials and Extract of Alkaloids
3.2. Conditions of UHPLC/QTOF-MS/MS
3.3. Drugs and Chemicals
3.4. Cell Culture
3.5. Determination of Cell Viability
3.6. Measurement of Nitric Oxide (NO)
3.7. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
3.8. Western Blot Analysis
3.9. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Shi, C.; Yuan, S.; Zhang, H.; Zhang, T.; Wang, L.; Xu, Z. Cell-mediated immune responses and protective efficacy against infection with Mycobacterium tuberculosis induced by Hsp65 and hIL-2 fusion protein in mice. Scand. J. Immunol. 2010, 69, 140–149. [Google Scholar] [CrossRef] [PubMed]
- Deng, J.-S.; Huang, G.-J.; Huang, S.-S.; Wang, S.-Y.; Chang, Y.-S.; Kuo, Y.-H. Bioassay Guided Isolation and Identification of Anti-inflammatory Active Compounds from the Root of Ficus formosana. J. Agric. Food Chem. 2013, 61, 11008–11015. [Google Scholar]
- Véronique, B.; Michael, K. Is NF-κB a good target for cancer therapy? Hopes and pitfalls. Nat. Rev. Drug Discov. 2009, 8, 33–40. [Google Scholar]
- Leen, C.; Geert, L.V. Inflammation and the Metabolic Syndrome: The Tissue-Specific Functions of NF-κB. Trends Cell Biol. 2017, 27, 417–429. [Google Scholar]
- Hua, Y.; Drew, P.; Richard, J. STATs in cancer inflammation and immunity: A leading role for STAT3. Nat. Rev. Cancer 2009, 9, 798–809. [Google Scholar]
- Martínez-Soto, D.; Ruiz-Herrera, J. Functional analysis of the MAPK pathways in fungi. Rev. Iberoam. Micol. 2017, 34, 192–202. [Google Scholar] [CrossRef] [PubMed]
- Matthews, C.P.; Colburn, N.H.; Young, M.R. AP-1 a target for cancer prevention. Curr. Cancer Drug Targets 2007, 7, 317–324. [Google Scholar] [CrossRef] [PubMed]
- Yao, B.; Wang, S.; Xiao, P.; Wang, Q.; Hea, Y.; Zhang, Y.; He, Y. MAPK signaling pathways in eye wounds: Multifunction and cooperation. Exp. Cell Res. 2017, 359, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Reis, F.S.; Martins, A.; Vasconcelos, M.H.; Morales, P.; Ferreira, I.C. Functional foods based on extracts or compounds derived from mushrooms. Trends Food Sci. Technol. 2017, 66, 48–62. [Google Scholar] [CrossRef]
- Wani, T.A.; Shah, A.G.; Wani, S.M.; Wani, I.A.; Masoodi, F.A.; Nissar, N.; Shagoo, M.A. Suitability of Different Food Grade Materials for the Encapsulation of Some Functional Foods Well Reported for Their Advantages and Susceptibility. Crit. Rev. Food Sci. Nutr. 2016, 56, 2431–2454. [Google Scholar] [CrossRef]
- Guo, Y.-G.; Ding, Y.-H.; Wu, G.-J.; Zhu, S.-L.; Sun, Y.-F.; Yan, S.-K.; Qian, F.; Jin, H.-Z.; Zhang, W.-D. Three new alkaloids from Xylopia vielana and their antiinflammatory activities. Fitoterapia 2018, 127, 96–100. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.L.; Liu, Y.P.; Zhang, C.; Zhao, W.H.; Shi, S.; He, D.N.; Zhang, P.; Liu, X.H.; Han, T.T.; Fu, Y.H. Carbazole alkaloids from Clausena hainanensis with their potential antiproliferative activities. Bioorg. Chem. 2017, 76, 359–364. [Google Scholar] [CrossRef] [PubMed]
- Wei, R.; Ma, Q.; Li, T.; Liu, W.; Sang, Z.; Li, M.; Liu, S. Carbazole alkaloids with antiangiogenic activities from Clausena sanki. Bioorg. Chem. 2018, 77, 387–392. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.-L.; Wang, L.; Wang, E.-J.; Zhang, G.-L.; Chen, B.; Wang, M.-K.; Li, F. Flavonoid glycosides and alkaloids from the embryos of Nelumbo nucifera seeds and their antioxidant activity. Fitoterapia 2018, 125, 184–190. [Google Scholar] [CrossRef] [PubMed]
- Lukhoba, C.W.; Simmonds, M.S.; Paton, A.J. Plectranthus: A review of ethnobotanical uses. J. Ethnopharmacol. 2006, 103, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Salminen, K.A.; Meyer, A.; Jeřábková, L.; Korhonen, L.E.; Rahnasto, M.; Juvonen, R.O.; Imming, P.; Raunio, H. Inhibition of human drug metabolizing cytochrome P450 enzymes by plant isoquinoline alkaloids. Phytomedicine 2011, 18, 533–538. [Google Scholar] [CrossRef] [PubMed]
- Gu, X.-Y.; Shen, X.-F.; Wang, L.; Wu, Z.-W.; Li, F.; Chen, B.; Zhang, G.-L.; Wang, M.-K. Bioactive steroidal alkaloids from the fruits of Solanum nigrum. Phytochemistry 2018, 147, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Wang, L.; Di, S.-N.; Xu, Q.; Ren, Q.-C.; Chen, S.-Z.; Huang, N.; Jia, D.; Shen, X.-F. Steroidal alkaloid solanine A from Solanum nigrum Linn. exhibits anti-inflammatory activity in lipopolysaccharide/interferon γ-activated murine macrophages and animal models of inflammation. Biomed. Pharmacother. 2018, 105, 606–615. [Google Scholar] [CrossRef] [PubMed]
- Changkija, S. Folk Medicinal Plants of the Nagas in India. Asian Folk. Stud. 1999, 58, 205. [Google Scholar] [CrossRef]
- Jin, Y.; Yuan, K. Studies on the Functional Components and Bioactivity and the Relativity of Bamboo Shoots and Shells. Appl. Mech. Mater. 2012, 108, 314–319. [Google Scholar] [CrossRef]
- Gong, W.; Xiang, Z.; Ye, F.; Zhao, G. Composition and structure of an antioxidant acetic acid lignin isolated from shoot shell of bamboo (Dendrocalamus Latiforus). Ind. Crop Prod. 2016, 91, 340–349. [Google Scholar] [CrossRef]
- Yang, Y.F. A study on the flavonoid compound in bamboo shoots of three pleioblastus species. J. Bamboo Res. 2009, 28, 56–60. [Google Scholar]
- Luo, X.; Wang, Q.; Zheng, B.; Lin, L.; Chen, B.; Zheng, Y.; Xiao, J. Hydration properties and binding capacities of dietary fibers from bamboo shoot shell and its hypolipidemic effects in mice. Food Chem. Toxicol. 2017, 109, 1003–1009. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Zhang, S.; Wang, Q.; Lu, X.; Lin, L.; Tian, Y.; Xiao, J.; Zheng, B. Characterization and hypoglycemic activity of a beta-pyran polysaccharides from bamboo shoot (Leleba oldhami Nakal) shells. Carbohydr. Polym. 2016, 144, 438–446. [Google Scholar] [CrossRef] [PubMed]
- Thomas, R.; Jebin, N.; Saha, R.; Sarma, D. Antioxidant and antimicrobial effects of kordoi (Averrhoa carambola) fruit juice and bamboo (Bambusa polymorpha) shoot extract in pork nuggets. Food Chem. 2016, 190, 41–49. [Google Scholar] [CrossRef]
- Zhang, Y.; Xu, P.; Xie, Y.H.; Du, W.P.; Jiang, P.; Yao, Z.Z.; Li, B. Phyllostachys chemical and its resistance to liver damage components research. J. Anhui Agric. Sci. 2013, 31, 12280–12282. [Google Scholar]
- Hamscher, G.; Priess, B.; Nau, H.; Panariti, E. Determination of Colchicine Residues in Sheep Serum and Milk Using High-Performance Liquid Chromatography Combined with Electrospray Ionization Ion Trap Tandem Mass Spectrometry. Anal. Chem. 2005, 77, 2421–2425. [Google Scholar] [CrossRef]
- Soufli, I.; Toumi, R.; Rafa, H.; Touil-Boukoffa, C.; Soufli, R.T.I. Overview of cytokines and nitric oxide involvement in immuno-pathogenesis of inflammatory bowel diseases. World J. Gastrointest. Pharmacol. Ther. 2016, 7, 353–360. [Google Scholar] [CrossRef]
- Ali, A.M.; Habeeb, R.A.; El-Azizi, N.O.; Khattab, D.A.; Abo-Shady, R.A.; Elkabarity, R.H. Higher nitric oxide levels are associated with disease activity in Egyptian rheumatoid arthritis patients. Rev. Bras. Reum. 2014, 54, 446–451. [Google Scholar] [CrossRef]
- Asiimwe, N.; Yeo, S.G.; Kim, M.S.; Jung, J.Y.; Jeong, N.Y. Nitric oxide: Exploring the contextual link with alzheimer’s disease. Oxid. Med. Cell. Longev. 2016, 2016, 7205747. [Google Scholar] [CrossRef]
- Aktan, F. iNOS-mediated nitric oxide production and its regulation. Life Sci. 2004, 75, 639–653. [Google Scholar] [CrossRef] [PubMed]
- Garret, A.; Fitz, G. COX-2 and beyond: Approaches to prostaglandin inhibition in human disease. Nat. Rev. Drug Discov. 2003, 2, 879–890. [Google Scholar]
- O’Callaghan, G.; Houston, A. Prostaglandin E2 and the EP receptors in malignancy: Possible therapeutic targets? Br. J. Pharmacol. 2016, 172, 5239–5250. [Google Scholar] [CrossRef] [PubMed]
- Wojdasiewicz, P.; Poniatowski, Ł.A.; Szukiewicz, D. The Role of Inflammatory and Anti-Inflammatory Cytokines in the Pathogenesis of Osteoarthritis. Mediat. Inflamm. 2014, 2014, 561459. [Google Scholar] [CrossRef] [PubMed]
- Tracey, K.J. The inflammatory reflex. Nature 2010, 257, 121. [Google Scholar] [CrossRef] [PubMed]
- Siebert, S.; Tsoukas, A.; Robertson, J.; McInnes, I. Cytokines as Therapeutic Targets in Rheumatoid Arthritis and Other Inflammatory Diseases. Pharmacol. Rev. 2015, 67, 280–309. [Google Scholar] [CrossRef] [PubMed]
- Wada, T.; Penninger, J.M. Mitogen-activated protein kinases in apoptosis regulation. Oncogene 2004, 23, 2838–2849. [Google Scholar] [CrossRef] [PubMed]
- Broom, O.J.; Widjaya, B.; Troelsen, J.; Olsen, J.; Nielsen, O.H. Mitogen activated protein kinases: A role in inflammatory bowel disease? Clin. Exp. Immunol. 2010, 158, 272–280. [Google Scholar] [CrossRef] [PubMed]
- Tai, C.J.; Chang, S.J.; Leung, P.C.; Tzeng, C.R. ATP activates nuclear translocation of mitogen-activated protein kinases in human granulosa-luteal cells. J. Clin. Endocrinol. Metab. 2013, 13, 679–692. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are available from the authors. |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Rt (min) | [M + H]+ | Formula | Identification | MS/MS Fragments | ||
|---|---|---|---|---|---|---|---|
| m/z theo | m/z exp | Error (ppm) | |||||
| Common alkaloids in the bamboo shoots and shells of P. amarus | |||||||
| 1 | 1.47 | 118.0863 | 118.0865 | 1.7 | C5H11NO2 | Betaine a | 100.076 (C5H10NO+), 82.065 (C5H8N+), 58.069 (C3H8N+) |
| 2 | 6.15 | 142.0863 | 142.0863 | 0 | C7H11NO2 | Arecaidine a | 124.077 (C7H10NO+), 100.076 (C5H10NO+), 96.084, 69.035 |
| 3 | 6.23 | 252.1091 | 252.1089 | −0.8 | C10H13N5O3 | Cordycepin a | 136.062 (C5H6N5+), 119.036 (C5H3N4+) |
| 4 | 7.85 | 156.1019 | 156.1017 | −1.3 | C8H13NO2 | Arecoline a | 138.093 (C8H12NO+), 110.103 (C7H12N+), 86.061, 69.037 |
| 5 | 8.07 | 238.1438 | 238.1439 | 0.4 | C13H19NO3 | - c | 164.107 (C9H10NO2+), 121.065, 103.055, 93.070, 91.055, 77.040 |
| 6 | 8.88 | 316.1543 | 316.1545 | 0.6 | C18H21NO4 | Cephalotaxine a | 283.041, 267.019, 169.098, 150.091, 133.064, 121.065, 103.054, 93.071, 91.054, 77.040 |
| 7 | 9.25 | 245.1648 | 245.1648 | 0 | C15H20N2O | 2-(4-Piperidinylcarbonyl)-1,2,3,4-tetrahydroisoquinoline b | 162.090 (C10H12NO+), 125.070 (C6H9N2O+), 70.067 (C4H8N+), 121.064, 103.054, 93.069, 91.055, 77.039 |
| 8 | 9.81 | 176.0706 | 176.0701 | −2.8 | C10H9NO2 | - c | 158.059 (C10H8NO+), 130.065 (C9H8N+), 118.066 (C8H8N+), 128.049, 103.055, 91.055, 77.040 |
| 9 | 10.32 | 137.1073 | 137.1072 | −0.7 | C8H12N2 | Ligustrazine a | 121.077 (C7H9N2+), 96.081 (C6H10N+), 70.065 (C4H8N+), 94.067, 80.051, 69.072, 55.057 |
| 10 | 12.35 | 314.1387 | 314.1393 | 1.9 | C18H19NO4 | N-feruloyltyramine a | 177.055, 149.060, 145.029, 121.066, 117.035, 103.055, 93.071, 91.055, 89.040, 77.041 |
| 11 | 12.72 | 227.0815 | 227.0816 | 0.4 | C13H10N2O2 | - c | 210.055 (C13H8NO2+), 199.050 (C11H7N2O2+), 198.055 (C12H8NO2+), 197.071 (C12H9N2O+), 181.076 (C12H9N2+), 169.076 (C11H9N2+), 154.065 (C11H8N+), 153.065 (C7H9N2O2+), 184.076, 144.080, 127.055, 115.055, 103.054, 77.039 |
| 12 | 12.75 | 336.1230 | 336.1232 | 0.6 | C20H17NO4 | Berberine a | 320.091 (C19H14NO4+), 306.076 (C18H12NO4+), 304.097 (C19H14NO3+), 292.097 (C18H14NO3+), 278.081 (C17H12NO3+), 262.086 (C17H12NO2+), 275.093 |
| 13 | 16.00 | 268.1332 | 268.1332 | 0 | C17H17NO2 | 4-Acetyl-N-(2,6-dimethylphenyl)benzamide b | 174.094, 147.046, 131.049, 121.066, 119.051, 105.073, 103.055, 91.055, 79.057, 77.041, 65.041 |
| 14 | 17.03 | 386.1598 | 386.1598 | 0 | C21H23NO6 | Colchiceine b [27] | 386.162 (C21H24NO6+), 368.149 (C21H22NO5+), 358.164 (C20H24NO5+), 342.134 (C19H20NO5+), 328.118 (C18H18NO5+), 326.139 (C19H20NO4+), 310.112 (C18H16NO4+), 162.089 (C10H12NO+), 308.131, 280.138, 179.071, 105.034 |
| 15 | 29.89 | 400.3210 | 400.3211 | 0.2 | C26H41NO2 | - c | 138.090 (C8H12NO+), 164.071, 121.064, 93.070 |
| Alkaloids only in bamboo shoots of P. amarus | |||||||
| 16 | 8.24 | 326.1598 | 326.1601 | 0.9 | C16H23NO6 | Monocrotaline a | 150.091, 131.070, 121.065, 103.039, 85.029, 57.035 |
| 17 | 10.22 | 272.1281 | 272.1281 | 0 | C16H17NO3 | - c | 216.102 (C13H14NO2+), 194.081 (C10H12NO3+), 135.043, 121.067, 91.054 |
| 18 | 10.23 | 335.1750 | 335.1757 | 2.1 | C21H22N2O2 | - c | 188.107 (C12H14NO+), 215.112, 198.091, 121.065, 103.054, 95.061, 93.071, 91.055, 77.039 |
| 19 | 10.7 | 342.1340 | 342.134 | 0 | C19H19NO5 | Taspine’s derivate b | 310.108 (C18H16NO4+), 162.090 (C10H12NO+), 327.111, 292.097, 278.081, 266.081, 248.071, 220.0753, 189.078 |
| 20 | 11.42 | 322.1074 | 322.1074 | 0 | C19H15NO4 | Berberrubine a | 278.079 (C17H12NO3+), 307.085, 279.088, 264.065, 250.083 |
| 21 | 11.6 | 395.19653 | 395.1964 | −0.3 | C23H26N2O4 | - c | 377.203 (C23H25N2O3+), 367.165 (C21H23N2O4+), 311.175 (C19H23N2O2+), 275.139 (C15H19N2O3+), 247.103 (C13H15N2O3+), 162.091 (C10H12NO+), 136.076 (C8H10NO+), 229.098, 155.080, 121.065 |
| 22 | 15.68 | 306.1700 | 306.1697 | −1.0 | C17H23NO4 | - c | 260.131 (C15H18NO3+), 216.101 (C13H14NO2+), 181.112 (C9H16NO3+), 140.0706 (C7H10NO2+), 121.066, 113.026, 91.054 |
| 23 | 17.55 | 325.191 | 325.1911 | 0.3 | C20H24N2O2 | - c | 162.090 (C10H12NO+), 204.138, 203.154, 118.065, 105.033, 100.112, 77.039 |
| 24 | 18.5 | 370.129 | 370.1285 | −1.4 | C20H19NO6 | Taspine b | 352.119 (C20H18NO5+), 342.135 (C19H20NO5+), 310.109 (C18H16NO4+), 162.090 (C10H12NO+), 327.111, 292.097, 278.082, 266.081, 248.069, 220.075, 189.078, 130.065, 105.033 |
| 25 | 31.81 | 430.3316 | 430.3322 | 1.4 | C27H43NO3 | - c | 166.087 (C9H12NO2+), 120.082 (C8H10N+) |
| Alkaloids only in bamboo shoot shells of P. amarus | |||||||
| 26 | 7.95 | 138.0550 | 138.0549 | −0.7 | C7H7NO2 | Trigonelline a | 120.048 (C7H6NO+), 94.032 (C6H8N+), 82.068 (C5H8N+), 66.037 (C4H4N+), 65.042 |
| 27 | 8.27 | 195.0877 | 195.0875 | −1 | C8H10N4O2 | Caffeine a | 138.067 (C6H8N3O+), 136.051 (C6H6N3O+), 110.072 (C5H8N3+), 123.042, 83.062, 69.047 |
| 28 | 9.46 | 328.1543 | 328.1544 | 0.3 | C19H21NO4 | - c | 208.098 (C11H14NO3+), 176.076 (C10H10NO2+), 273.112, 191.070, 121.065, 107.049, 103.055, 93.071 |
| 29 | 10.28 | 337.1911 | 337.1913 | 0.6 | C21H24N2O2 | Catharanthine a | 217.134, 121.065, 103.055, 97.077, 93.070, 77.039 |
| 30 | 11.99 | 353.1496 | 353.1495 | −0.3 | C20H20N2O4 | N-Feruloylserotonin b | 160.076 (C10H10NO+), 221.105, 177.055, 145.028, 117.033, 89.039 |
| 31 | 12.24 | 323.1390 | 323.1393 | 0.9 | C19H18N2O3 | N-p-Coumaroylserotonin b | 229.097 (C13H13N2O2+), 159.092 (C10H11N2+), 130.065 (C9H8N+), 295.143, 278.117, 245.093, 172.076, 158.059, 131.051, 121.065, 107.051, 103.054, 93.070, 91.0560, 77.040 |
| 32 | 31.5 | 404.3159 | 404.3162 | 0.7 | C25H41NO3 | N-Palmitoyl-L-phenylalanine b | 166.086 (C9H12NO2+), 120.081 (C8H10N+), 149.059, 131.049, 103.055 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ren, Y.; Ma, Y.; Zhang, Z.; Qiu, L.; Zhai, H.; Gu, R.; Xie, Y. Total Alkaloids from Bamboo Shoots and Bamboo Shoot Shells of Pleioblastus amarus (Keng) Keng f. and Their Anti-Inflammatory Activities. Molecules 2019, 24, 2699. https://doi.org/10.3390/molecules24152699
Ren Y, Ma Y, Zhang Z, Qiu L, Zhai H, Gu R, Xie Y. Total Alkaloids from Bamboo Shoots and Bamboo Shoot Shells of Pleioblastus amarus (Keng) Keng f. and Their Anti-Inflammatory Activities. Molecules. 2019; 24(15):2699. https://doi.org/10.3390/molecules24152699
Chicago/Turabian StyleRen, Yan, Yisha Ma, Zhidan Zhang, Liying Qiu, Huanhuan Zhai, Ruimeng Gu, and Yaping Xie. 2019. "Total Alkaloids from Bamboo Shoots and Bamboo Shoot Shells of Pleioblastus amarus (Keng) Keng f. and Their Anti-Inflammatory Activities" Molecules 24, no. 15: 2699. https://doi.org/10.3390/molecules24152699
APA StyleRen, Y., Ma, Y., Zhang, Z., Qiu, L., Zhai, H., Gu, R., & Xie, Y. (2019). Total Alkaloids from Bamboo Shoots and Bamboo Shoot Shells of Pleioblastus amarus (Keng) Keng f. and Their Anti-Inflammatory Activities. Molecules, 24(15), 2699. https://doi.org/10.3390/molecules24152699