
Larvicidal Compounds Extracted from Helicteres velutina K. Schum (Sterculiaceae) Evaluated against Aedes aegypti L.

 , , ,
, , , 
Abstract
:
1. Introduction
2. Results
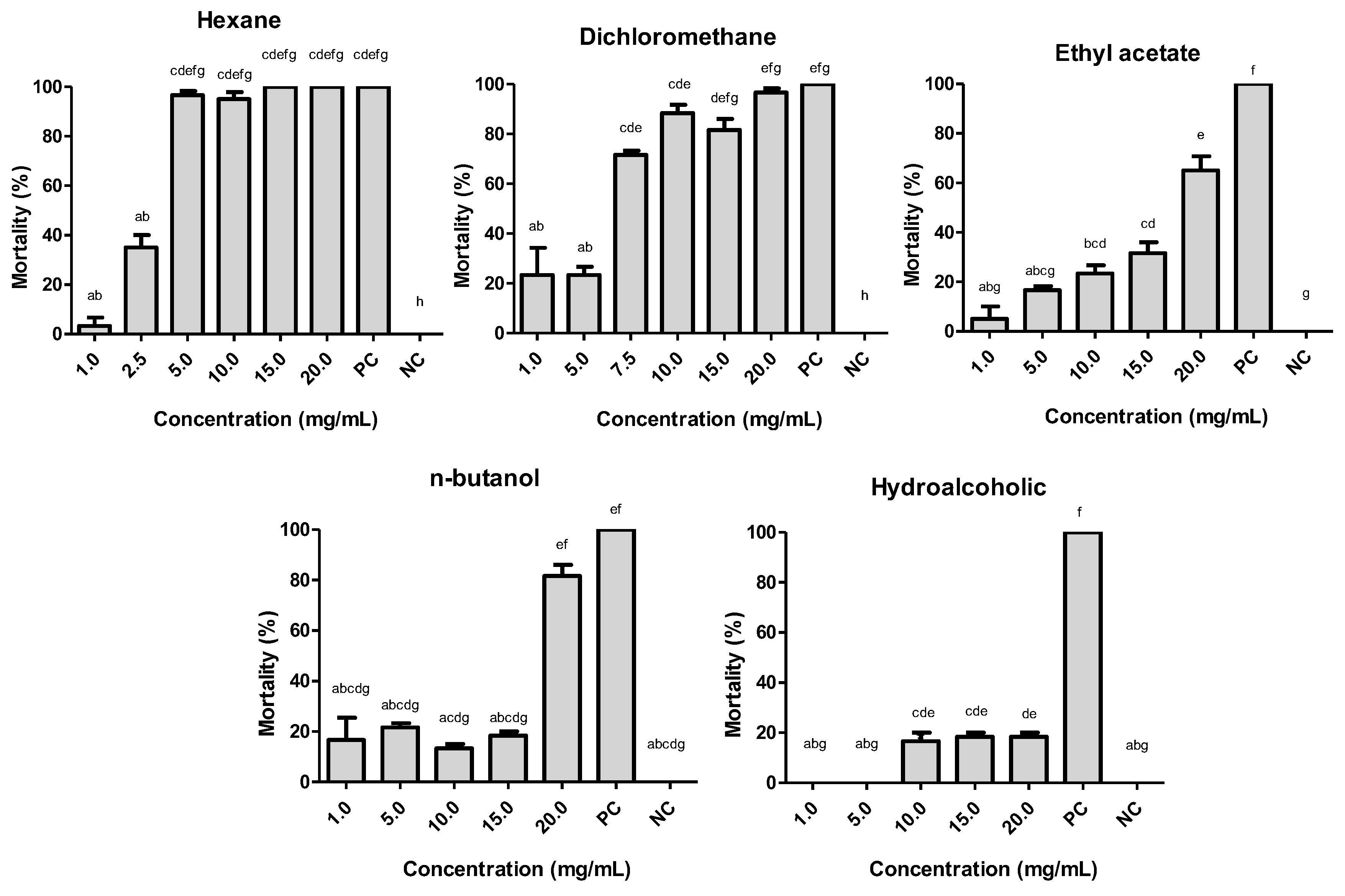
2.1. Larvicidal Activity of Helicteres velutina Fractions
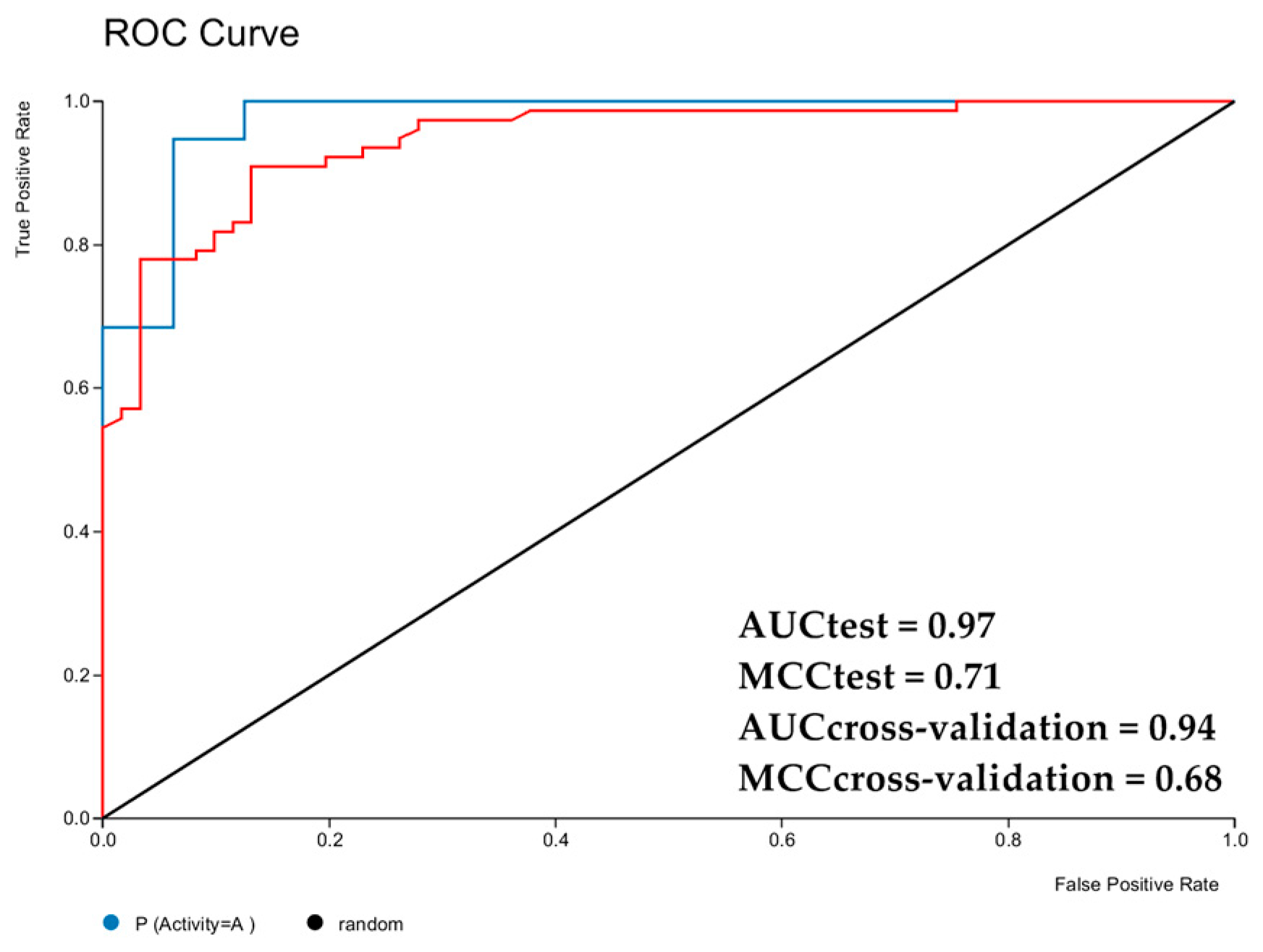
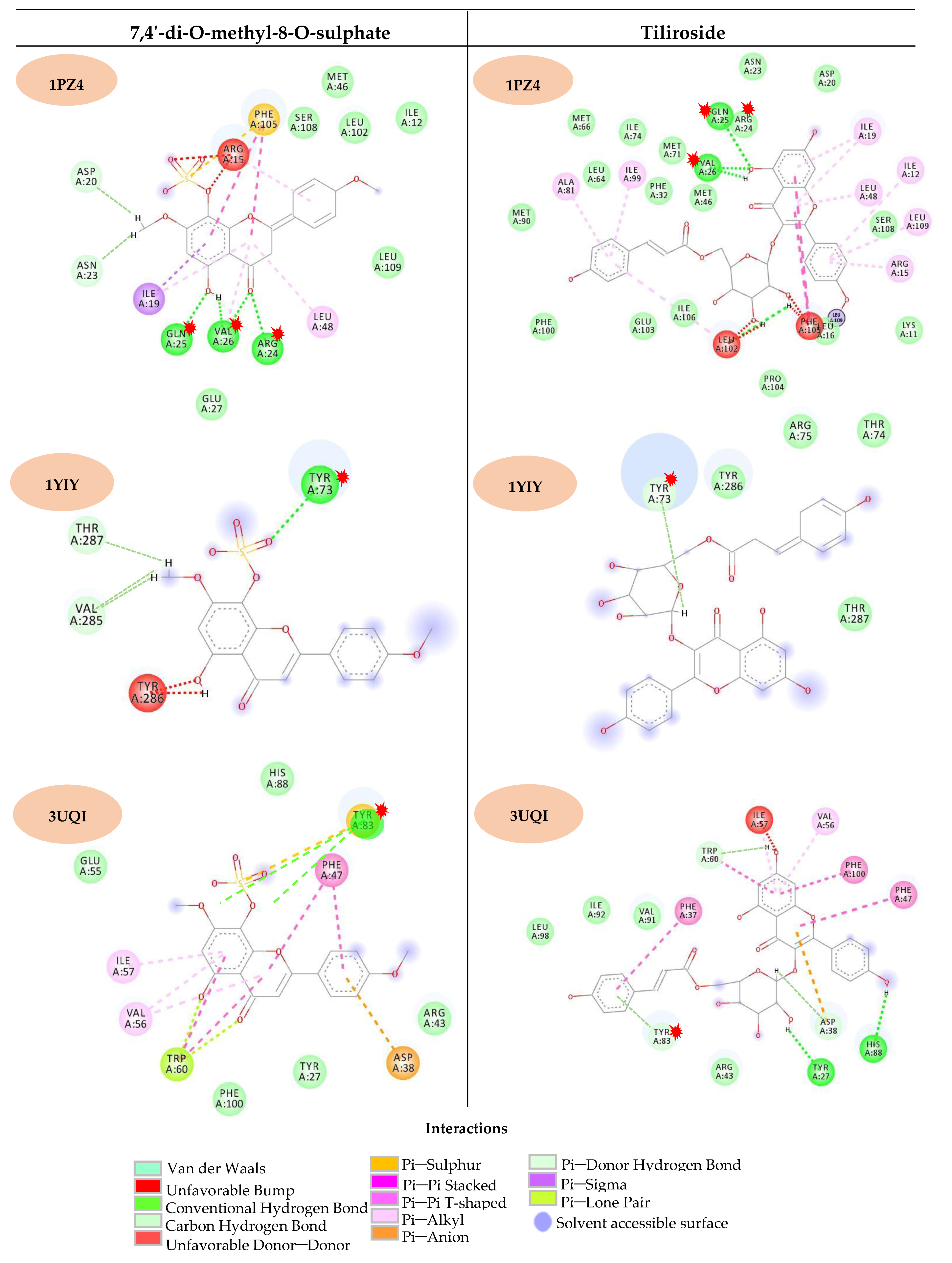
2.2. Computational Study
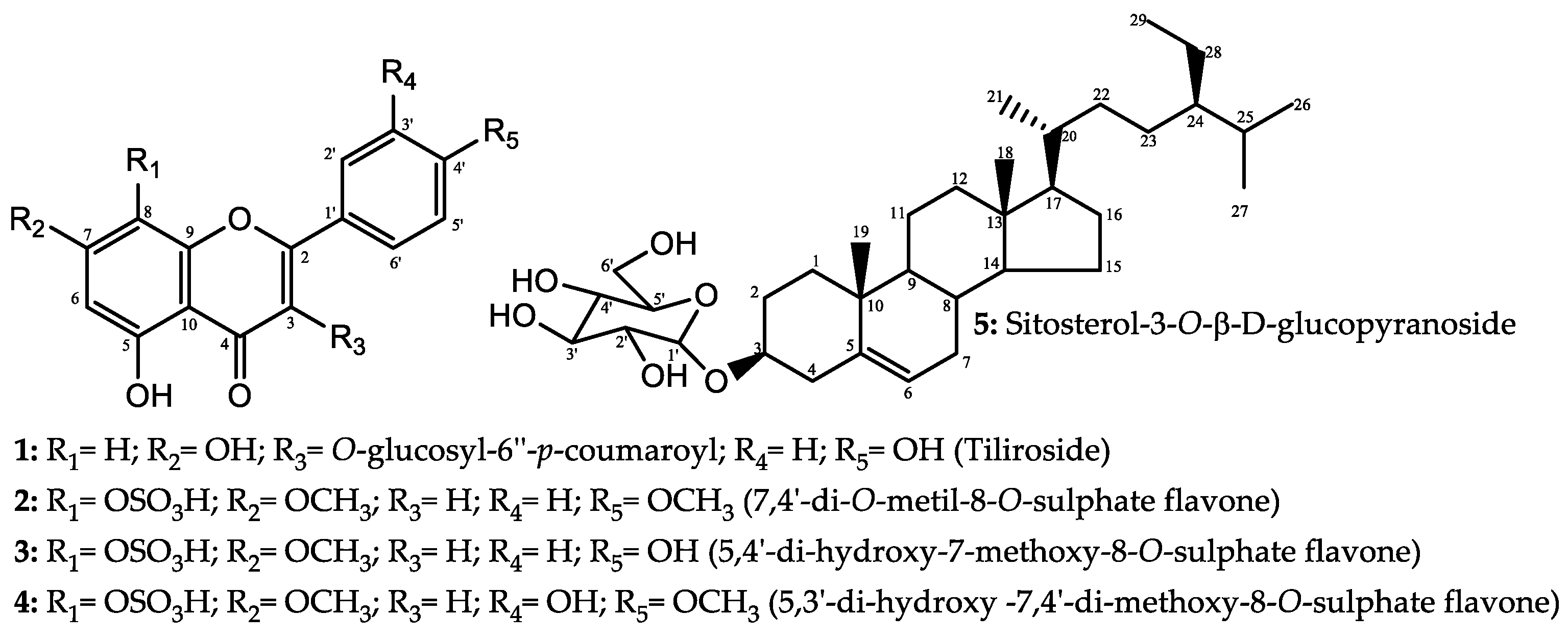
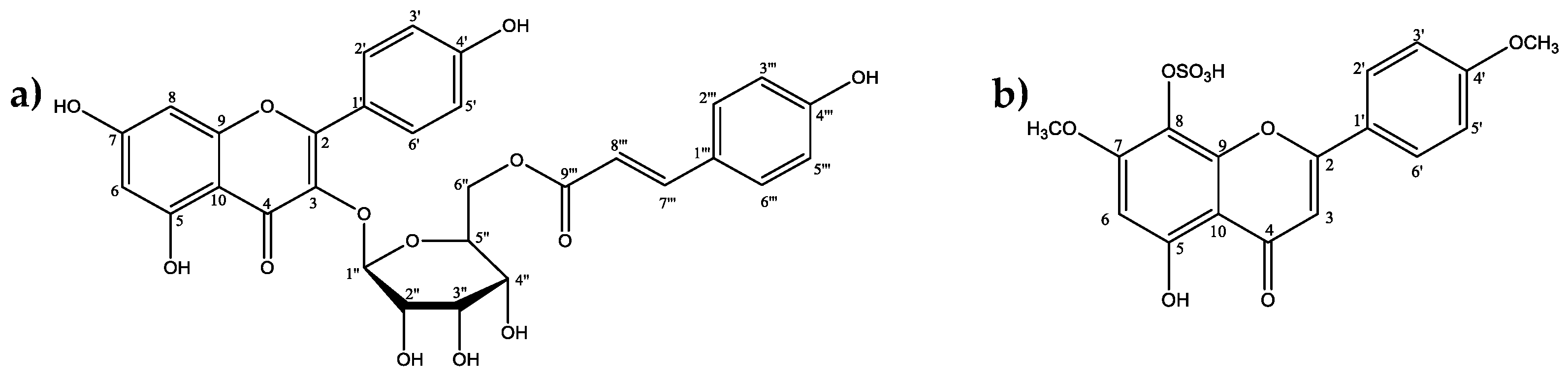
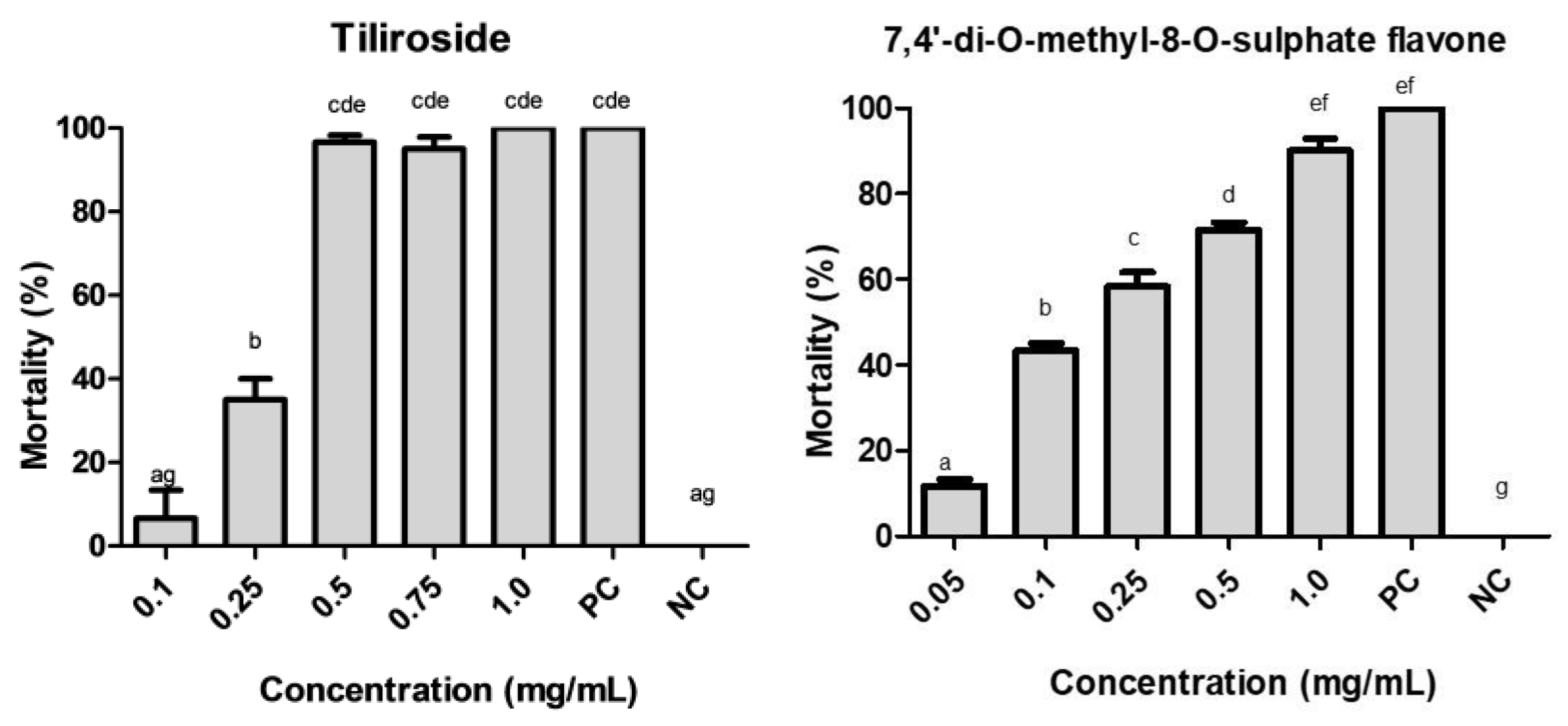
2.3. Larvicidal Activity of the Promising Substances
3. Discussion
4. Materials and Methods
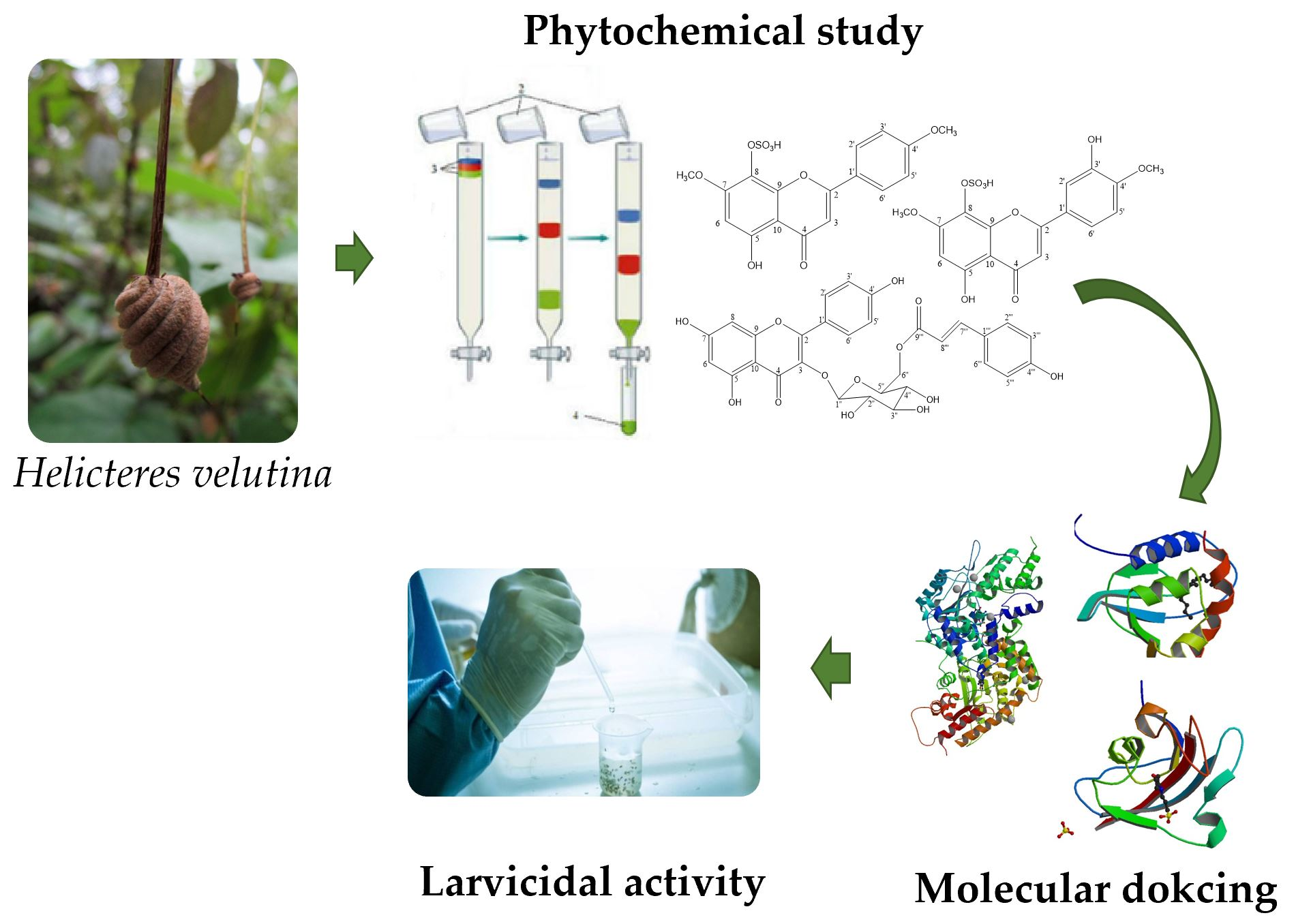
4.1. Collection, Extraction, and Compound Isolation
4.2. Virtual Screening Focused on Medicinal Chemistry
4.3. Molecular Docking
4.4. Larvicidal Activity
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Botas, G.S.; Cruz, R.A.S.; Almeida, F.B.; Duarte, J.L.; Araújo, R.S.; Souto, R.N.P.; Ferreira, R.; Carvalho, J.C.T.; Santos, M.G.; Rocha, L.; et al. Bacccharis reticularia DC. And Limonene Nanoemulsions: Promising larvicidal agentes for Aedes aegypti (Diptera: Culicidae) Control. Molecules 2017, 22, 1990. [Google Scholar] [CrossRef]
- Leta, S.; Beyene, T.J.; Clercq, E.M.; Amenu, K.; Revie, C. Global risk mapping for major diseases transmitted by Aedes aegypti and Aedes albopictus. Int. J. Infect. Dis. 2018, 67, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Mukandiwa, L.; Eloff, J.N.; Naidoo, V. Larvicidal activity of leaf extracts and seselin from Clausena anisata (Rutaceae) against Aedes aegypti. S. Afr. J. Bot. 2016, 100, 169–173. [Google Scholar] [CrossRef]
- Ravidran, J.; Samuel, T.; Alex, E.; William, J. Adulticial activity of Ageratum houstonianum Mill. (Asteraceae) leaf extracts against three vector mosquito species (Diptera: Culicidae). Asian Pac. J. Trop. Dis. 2012, 2, 177–179. [Google Scholar] [CrossRef]
- Nunes, F.C.; Leite, J.A.; Oliveira, L.H.G.; Sousa, P.A.P.S.; Menezes, M.C.; Moraes, J.P.S.; Mascarenhas, S.R.; Braga, V.A. The larvicidal activity of Agave sisalana against L4 larvae of Aedes aegypti is mediated by internal necrosis and inhibition of nitric oxide production. Parasitol. Res. 2015, 114, 543–549. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, L.H.G.; Lacerda, D.; Nunes, F.C. Effects of hecogenin on larvicidal activity against Aedes aegypti mosquito, the dengue vector. BMC Proc. 2014, 8, P33. [Google Scholar] [CrossRef]
- Oliveira, L.H.G.; Sousa, P.A.P.S.; Hilario, F.F.; Nascimento, G.J.; Morais, J.P.S.; Medeiros, E.P.; Sousa, M.F.; Nunes, F.C. Agave sisalana extract induces cell death in Aedes aegypti hemocytes increasing nitric oxide production. Asian Pac. J. Trop. Biomed. 2016, 6, 396–399. [Google Scholar] [CrossRef]
- Murugan, K.; Kumar, P.M.; Kovedan, K.; Amerasan, D.; Subrmaniam, J.; Hwang, J. Larvicidal, pupicidal, repellent and aduticidal activity of Citrus sinensis orange peel extracts against Anopheles stephensi, Aedes aegypti and Culex quinquefasciatus (Diptera: Culicidae). Parasitol. Res. 2012, 111, 1757–1769. [Google Scholar] [CrossRef]
- Rojas-Pizón, P.A.; Silva-Fernandez, J.J.; Dussán, J. Laboratory and simulated-field bioassays for assessing mixed cultures of Lysinibacillus sphaericus against Aedes aegypti (Diptera: Culicidae) larvae resistant to temephos. Appl. Entomol. Zool. 2018, 53, 183–191. [Google Scholar] [CrossRef]
- Souza, T.M.; Farias, D.F.; Soares, B.M.; Viana, M.P.; Lima, G.P.G.; Machado, L.K.A.; Morais, S.M.; Carvalho, A.F.U. Toxicity of Brazilian plant seed extracts to two strains of Aedes aegypti (Diptera: Culicidae) and Nontarget animals. Entomol. Soc. Am. 2011, 48, 846–851. [Google Scholar] [CrossRef] [PubMed]
- Vivekanandhan, P.; Senthil-Nathan, S.; Shivakumar, M.S. Larvicidal, pupicidal and adult smoke toxic effects of Acanthospermum hispidum (DC) leaf crude extracts against mosquito vectors. Physilogical Mol. Plant Pathol. 2018, 101, 156–162. [Google Scholar] [CrossRef]
- Govindarajan, M. Mosquito larvicidal and ovicidal activity of Cardiospermum halicacabum Linn. Family: Sapindaceae) leaf extract against Culex quinquefasciatus (Say) and Aedes aegypti (Linn.) (Diptera: Culicidae). Eur. Rev. Med. Pharmacol. Sci. 2011, 15, 787–794. [Google Scholar] [PubMed]
- Munusamy, R.G.; Appadurai, D.R.; Kuppusamy, S.; Michael, G.P.; Savarimuthu, I. Ovicidal and larvicidal activities of some plant extracts against Aedes aegypti L. and Culex quinquefasciatus Say (Diptera: Culicida). Asian Pac. J. Trop. Dis. 2016, 6, 468–471. [Google Scholar] [CrossRef]
- Chase, M.W.; Christenhusz, M.J.M.; Fay, M.F.; Byng, J.W.; Judd, W.S.; Soltis, D.E.; Soltis, D.E.; Mabberley, D.J.; Sennikov, A.N.; Soltis, P.S.; et al. APG IV—Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV. Bot. J. Linn. Soc. 2016, 181, 1–20. [Google Scholar] [CrossRef]
- Santos, E.A.; Carvalho, C.M.; Costa, A.L.S.; Conceição, A.S.; Moura, F.B.P.; Santana, A.E.G. Bioactivity evaluation of plants extracts used in indigenous medicine against the snail, Biomphalaria glabrata, and the larvae of Aedes aegypti. Evid. Based Complement. Altern. Med. 2012, 1, 1–9. [Google Scholar] [CrossRef]
- Fernandes, D.A.; Souza, M.S.R.; Teles, Y.C.F.; Oliveira, L.H.G.; Lima, J.B.; Conceição, A.S.; Nunes, F.C.; Silva, T.M.S.; Souza, M.F.V. New sulphated flavonoids and larvicidal activity of Helicteres velutina K. Schum (Sterculiaceae). Molecules 2018, 23, 2784. [Google Scholar] [CrossRef] [PubMed]
- Cechinel-Filho, V.; Yunes, R.A. Estratégias para a obtenção de compostos farmacologicamente ativos a partir de plantas medicinais. Conceitos sobre modificação estrutural para otimização da atividade. Quím. Nova 1997, 21, 99–105. [Google Scholar] [CrossRef]
- Santos, A.E. Evaluation of the Antioxidant Potential and Chemical Characterization of the Chromatographic Fractions and Ethanolic Extract of the Leaves of Bauhinia longifolia (Bong) Steudel. Bachelor’s Thesis, Universidade Federal do Rio de Janeiro, Ilha do Fundão, Brazil, 2008. Degree in Chemistry (Work Completion Course). [Google Scholar]
- Barros, R.P.C.; Cunha, E.V.L.; Catão, R.M.R.; Scotti, L.; Souza, M.S.R.; Brás, A.A.Q.; Scotti, M.T. Virtual screening of secondary metabolites of the genus Solanum with potencial antimicrobial activity. Braz. J. Pharmacogn. 2018, 28, 686–691. [Google Scholar] [CrossRef]
- Alves, M.F.; Ferreira, L.A.M.; Gadelha, F.A.A.F.; Ferreira, L.K.D.P.; Felix, M.B.; Scotti, M.T.; Scotti, L.; Oliveira, K.M.; Santos, S.G.; Diniz, M.F.F. Toxicological evaluation in silico and in vivo of secondary metabolites of Cissampelos sympodialis in Mus musculus mice following inhalation. Nat. Prod. Res. 2017, 4, 1–7. [Google Scholar] [CrossRef]
- Chibli, L.A.; Schmidt, T.J.; Nonato, M.C.; Calil, F.A.; Costa, F.B. Natural products as inhibitors of Leishmania major dihydroorotate dehydrogenase. Eur. J. Med. Chem. 2018, 157, 852–866. [Google Scholar] [CrossRef]
- Talete, S.R.L. Dragon—Software for Molecular Descriptor Claculation—Version 6.0. 2013. Available online: http://www.talete.mi.it/ (accessed on 25 November 2018).
- Matthews, B.W. Comparison of the predicted and observed secondary structure of T4 phage lysozyme. Biochim. Biophys. Acta Prot. Struct. 1975, 405, 442–451. [Google Scholar] [CrossRef]
- Silva, F.C. Análises ROC; Federal University of Pernambuco: Recife, Brazil, 2006. [Google Scholar]
- Zhang, S.; Golbraikh, A.; Oloff, S.; Kohn, H.; Tropsha, A.J. A novel automated lazy learning QSAR (ALL-QSAR) approach: Method development, applications, and virtual screening of chemical databases using validated ALL-QSAR models. Chem. Inf. Model. 2006, 46, 1984–1995. [Google Scholar] [CrossRef] [PubMed]
- Han, Q.; Gao, Y.G.; Robison, H.; Ding, H.; Wilson, S.; Li, J. Crystal structures of Aedes aegypti kynurenine aminotransferase. FEBS J. 2005, 272, 2198–2206. [Google Scholar] [CrossRef] [PubMed]
- Dyer, D.H.; Lovell, S.; Thoden, J.B.; Holden, H.M.; Rayment, I.; Lan, Q. The structural determination of an Insect Sterol Carrier Protein-2 with a Ligand-bound C16 fatty acid at 1.35-Å resolution. J. Biol. Chem. 2003, 278, 39085–39091. [Google Scholar] [CrossRef] [PubMed]
- Rajan, S.; Saw, K.Q.; Nguyen, Q.T.; Baek, K.; Yoon, H.S. High-resolution crystal structure of FKBP12 from Aedes aegypti. Protein Sci. 2012, 21, 1080–1084. [Google Scholar] [CrossRef] [PubMed]
- Lorenzo, V.; Lucio, A.S.; Tavares, J.F.; Filho, J.M.; Lima, T.K.; Rocha, J.D.; Scotti, M.T. Structure and Ligand Based approaches to evaluate aporphynic alkaloids from Annonaceae as Multi-target agente against Leishmania donovani. Curr. Pharm. Des. 2016, 22, 5196–5203. [Google Scholar] [CrossRef] [PubMed]
- Acevedo, C.H.; Scotti, L.; Scotti, M.T. In silico studies designed to select sesquiterpene lactones with potential antichagasic activity from an in-house Asteraceae Database. Chemmedchem 2018, 13, 634–645. [Google Scholar] [CrossRef] [PubMed]
- Pontual, E.V. Extrato de Flores de Moringa oleífera: Atividade Larvicida e Efeito Sobre a Tripsina e Acetilcolinesterase de Larvas de Aedes aegypti. Master’s Thesis, Universidade Federal do Pernambuco, Recife, Brasil, 2010. [Google Scholar]
- Murugan, K.; Murugan, P.; Noortheen, A. Larvicidal and repellent potencial of Albizzia amara Boivin and Ocimum basilicum Linn against dengue vector, Aedes aegypti (Insecta: Diptera: Culicidae). Bioresour. Technol. 2007, 98, 198–201. [Google Scholar] [CrossRef]
- Kovedan, K.; Murugan, K.; Kumar, P.M.; Thiyagarajan, P.; William, S.J. Ovicidal, repellent, adulticidal and field evaluations of plant extract against dengue, malaria and filarial vectors. Parasitol. Res. 2013, 112, 1205–1219. [Google Scholar] [CrossRef]
- Kumar, N.P.; Suresh, A.; Vanamail, P.; Sabesan, S.; Krishnamoorthy, K.G.; Mathew, J.; Jose, V.T.; Jambulingam, P. Chikungunya virus outbreak in Kerala, Inidia, 2007: A seroprevalence study. Mem. Inst. Oswaldo Cruz 2011, 106, 912–916. [Google Scholar] [CrossRef]
- Pluempanupat, S.; Kumrugsee, N.; Pluempanupat, W.; Ngamkitpinyo, K.; Chavasari, W.; Bullangtopi, V.; Koul, O. Laboratory evaluation of Dalbergia oliveri (Fabaceae: Fabales) extracts ans isolated isoflavonoids on Aedes aegypti (Diptera: Culicidae) mosquitoes. Ind. Crops Ans Prod. 2013, 44, 563–658. [Google Scholar] [CrossRef]
- Reegan, A.D.; Gandhi, M.R.; Paulraj, M.G.; Balakrishna, K.; Ignacimunthu, S. Effect of niloticin, a protolimonoid isolated from Limonia acidissima L. (Rutaceae) on the immature stages of dengue vector Aedes aegypti L. (Diptera: Culidae). Acta Trop. 2014, 139, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Sá, S.; Chaul, L.T.; Alves, V.F.; Fiuza, T.S.; Tresvenzol, B.G.V.; Ferri, P.H.; Borges, L.L.; Paula, J.R. Phytochemistry and antimicrobial activity of Campomanesia adamantium. Braz. J. Pharmacogn. 2018, 28, 303–311. [Google Scholar] [CrossRef]
- Krzyzaniak, L.M.; Antonelli-Ushirobira, T.M.; Panizzon, G.; Sereia, A.L.; Souza, J.R.P.; Zequi, J.A.C.; Novello, C.R.; Lopes, G.C.; Medeiros, D.C.M.; Silva, D.B.; et al. Larvicidal activity against Aedes aegypti and chemical characterization of the inflorescences of Tagetes patula. Evid. Based Complement. Altern. Med. 2017, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, G.L.; Cardoso, S.K.; Lara-Júnior, C.R.; Vieira, T.M.; Guimarães, E.F.; Figueiredo, L.S.; Martins, E.R.; Moreira, D.L.; Kaplan, M.A.C. Chemical study and larvicidal activity against Aedes aegypti of essencial oil of Piper aduncum L. (Piperaceae). An. Acad. Bras. Ciências 2013, 85, 1227–1234. [Google Scholar] [CrossRef] [PubMed]
- Heinzmann, B.M.; Barros, F.M.C. Potencial das plantas nativas brasileiras para o desenvolvimento de fitomedicamentos tendo como exemplo Lippia alba (Mill.) N.E. Brown (Verbenaceae). Saúde St. Maria 2007, 33, 43–48. [Google Scholar]
- Costa, M.S.; Pereira, M.J.B.; Oliveira, S.S.; Souza, P.T.; Dall’oglio, E.L.; Alves, T.C. Anonaceaes provocam mortalidade em larvas de Aedes aegypti (Linnaeus, 1762) (Diptera: Culicidae). Rev. Bras. Biociênc. 2013, 11, 184–190. [Google Scholar]
- Coelho, A.A.M.; de Paula, J.E.; Espindola, L.S. Atividade larvicida de extratos vegetais sobre Aedes aegypti (L.) (Diptera: Culicidae), em condições de laboratório. BioAssay 2009, 4, 1–6. [Google Scholar] [CrossRef]
- Govidarajan, M.; Sivakumar, R. Larvicidal, ovicidal and adulticidal efficacy of Erythrina indica (Lam.) (Family: Fabaceae) against Anopheles stephensi, Aedes aegypti and Culex quinquefasciatus (Diptera: Culicidae). Parasitol. Res. 2014, 113, 777–791. [Google Scholar] [CrossRef]
- Borah, R.; Kalita, M.C.; Talukdar, A.K. Herbal product for mosquito control and its process for preparation thereof. Indian Pat. Appl. 2014, 1, 1–25. [Google Scholar]
- Sukari, M.A.; Noor, H.S.M.; Bakar, N.H.A.; Ee, G.C.L.; Isamail, I.S.; Rahmani, M.; Abdul, A.B. Larvicidal carbazole alkaloids from Murraya koenigii against dengue fever mosquito Aedes aegypti Linnaeus. Asian J. Chem. 2013, 25, 7719–7721. [Google Scholar] [CrossRef]
- Arivoli, S.; Raveen, R.; Samuel, T. Larvicidal activity of Murraya koenigii (L.) Spreng (Rutaceae) hexane leaf extract isolated fractions against Aedes aegypti Linnaeus, Anopheles stephensi Liston and Culex quinquefasciatus Say (Diptera; Culicidae). J. Mosq. Res. 2015, 5, 1–8. [Google Scholar] [CrossRef]
- Guarido, M.M. Atividade Inseticida de Extratos de Nona foetida Mart. (Anonaceae) Sobre Estágios Imaturos de Aedes aegypti (Linnaeus, 1762) (Diptera: Culicidae). Master’s Thesis, Universidade do Paraná, Curitiba, Brazil, 2009. [Google Scholar]
- Torres, R.C.; Garbo, A.G.; Walde, R.Z.M.L. Larvicidal activity of Garcinia mangostana fruit wastes against dengue vector Aedes aegypti. J. Anim. Plant Sci. 2015, 25, 1187–1190. [Google Scholar]
- Govidarajan, M.; Karuppannan, P. Mosquito larvicidal and ovicidal properties of Eclipta alba (L.) Hassk (Asteraceae) against chikungunya vector, Aedes aegypti (Linn.) (Diptera: Culicidae). Asian Pac. J. Trop. Med. 2011, 4, 24–28. [Google Scholar] [CrossRef]
- Kumar, D.; Kumar, G.; Agrawal, V. Green synthesis of silver nanoparticles using Holarrhena antidysenterica (L.) Wall Bark extract and their larvicidal activity against dengue and filariasis vectors. Parasitol. Res. 2018, 117, 377–389. [Google Scholar] [CrossRef] [PubMed]
- Darmali, L.; Silva, C.B.; Campos, F.M.M.; Burci, L.M.; Nascimento, F.; Jesus, C.; Dias, J.F.G.; Dallarmi, M.M.; Gomes, O.M.; Zanin, S.M.W. Larvicidal activity of Dalbergia brasiliensis (Fabaceae-Papilioideae) on Aedes aegypti. Afr. J. Pharm. Pharmacol. 2015, 9, 881–885. [Google Scholar] [CrossRef]
- Teles, Y.C.F.; Souza, M.S.R.; Souza, M.F.V. Sulphated Flavonoids: Biosynthesis, Structures, and Biological Activities. Molecules 2018, 23, 480. [Google Scholar] [CrossRef]
- Teles, Y.C.F.; Horta, C.C.R.; Agra, M.F.; Siheri, W.; Boyd, M.; Igoli, J.O.; Gray, A.I.; Souza, M.F.V. New Sulphated Flavonoids from Wissadula periplocifolia (L.) C. Presl (Malvaceae). Molecules 2015, 20, 20161–20172. [Google Scholar] [CrossRef]
- Dimas, K.; Demetzos, C.; Mitaku, S.; Marselos, M.; Tzavaras, T.; Kokinopoulos, D. Citotoxic activity pf kaempferol glycosides against human leukaemic cells lines in vitro. Pharmacol. Res. 2000, 41, 85–88. [Google Scholar] [CrossRef]
- Matsuda, H.; Ninomiya, K.; Shimoda, H.; Yoshikawa, M. Hepatoprotective principles from the flowers of Tilia argentea (Linden): Structure requirements of tiliroside and mechanisms of action. Bioorg. Med. Chem. 2002, 10, 707–712. [Google Scholar] [CrossRef]
- Silva, G.C.; Pereira, A.C.; Rezende, B.A.; Silva, J.P.; Cruz, J.S.; Souza, M.F.V.; Gomes, R.A.; Teles, Y.C.F.; Cortes, S.F.; Lemos, V.S. Mechanism of the antihypertensive and vasorelaxant effects of the flavonoid tiliroside in resistance arteries. Planta Med. 2013, 79, 1003–1008. [Google Scholar] [CrossRef] [PubMed]
- Goto Tsuyoshi, H.; Mayuka, N.; Hiroyuki, N.; Akifumi, N.; Norihisa, M.Y.; Youichi, S.N. Tiliroside, a glycosidic flavonoid, inhibits carbohydrate digestion and glucose absorption in the gastrointestinal tract. Mol. Nutr. Food Res. 2012, 56, 435–445. [Google Scholar] [CrossRef] [PubMed]
- Sala, A.; Receio, M.C.; Schinella, G.R.; Manez, S.; Giner, R.M.; Cerdá-Nicolás, M.; Ríos, J.L. Assement of the radical scanvenger activity of tiliroside. Eur. J. Pharmacol. 2003, 461, 53–61. [Google Scholar] [CrossRef]
- Oliveira, A.M.F.; Pinheiro, S.L.; Pereira, C.K.; Matias, W.N.; Gomes, R.A.; Chaves, O.S.; Souza, M.F.V.; Almeida, R.N.; Assis, T.S. Total phenolic contente and antioxidante activity of some Malvaceae Family species. Antioxidants 2012, 1, 33–43. [Google Scholar] [CrossRef] [PubMed]
- Maciel, J.K.S.; Chaves, O.S.; Brito-Filho, S.G.; Teles, Y.C.F.; Fernandes, M.G.; Assis, T.S.; Fernandes, P.D.; Andrade, A.P.; Felix, L.P.; Silva, T.M.S.; et al. New alcamide and anti-oxidant activity of Pilosocereus gounellei A. Weber ex K. Schum. Bly. Rowl. (Cactaceae). Molecules 2016, 21, 11. [Google Scholar] [CrossRef]
- Rajkumar, S.; Jebanesan, A. Bioactivity of flavonoid compounds from Poncirus trifólia L. (Family: Rutaceae) against the dengue vector, Aedes aegypti L. (Diptera). Parasitol. Res. 2008, 104, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Perumalsamy, H.; Jang, M.J.; Kim, J.R.; Kadarkarai, M.; Ahn, Y.J. Larvicidal activity and possible mode of action of four flavonoids and two fatty acids identified in Millettia pinnata seed toward three mosquito species. Parasit. Vectors 2015, 8, 2–14. [Google Scholar] [CrossRef]
- Gikonyo, N.K.; Mwangi, R.W.; Midiwo, J.O. Toxicity and growth-inhibitory activity of Polygonum senegalense (Meissn.) surface exudate against Aedes aegypti larvae. Insect Sci. Appl. 1998, 18, 229–234. [Google Scholar] [CrossRef]
- Garcez, W.S.; Garcez, F.R.; Silva, L.M.G.E.; Sarmento, U.C. Substâncias de origem vegetal com atividade larvicida contra Aedes aegypti. Rev. Virtual Quím. 2013, 5, 363–393. [Google Scholar]
- WHO—World Health Organization. Insecticide resistance and vector control. In WHO Technical Reports Series; World Health Organization: Geneva, Switzerland, 1970. [Google Scholar]
Sample Availability: Samples of all isolated compounds are available from the authors. |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Training | Cross Validation | Test | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Samples | Matches | % Matches | Samples | Matches | % Matches | Samples | Matches | % Matches | |
| Active | 77 | 76 | 98% | 77 | 65 | 84% | 19 | 14 | 73% |
| Inactive | 61 | 61 | 100% | 61 | 53 | 86% | 16 | 15 | 93% |
| Total | 138 | 137 | 99% | 138 | 118 | 85% | 35 | 29 | 82% |
| Protein ID | Classification | Ligand | Localisation |
|---|---|---|---|
| Aedes aegypti kynurenine aminotransferase (1YIY) | Transferase | 4′-Deoxy-4′-aminopyridoxal-5′-phosphate; Pyridoxamine-5′-phosphate | Adult mosquito head |
| Sterol Carrier Protein-2 (1PZ4) | Lipid binding | Palmitic acid | Large intestine of larvae |
| AaFKBP12 (3UQI) | Isomerase | 3 [N-morpholino]propane sulphonic acid | Adult mosquito |
| Protein | Energy, PDB Value (kJ/mol) | Energy, MolDock Value (kJ/mol) | Redocking, RMSD Value |
|---|---|---|---|
| 1YIY | −103.002 | −124.274 | 0.158 |
| 1PZ4 | −106.678 | −110.680 | 0.203 |
| 3UQI | −54.802 | −57.032 | 0.138 |
| Molecule | Protein | p | ps | Pc |
|---|---|---|---|---|
| Tiliroside | 1YIY | 0.79 | 1 | 0.79 |
| 1PZ4 | 0.79 | 1 | 0.79 | |
| 3UQI | 0.79 | 1 | 0.79 | |
| 7,4′-di-O-methyl-8-O-sulphate flavone | 1YIY | 0.66 | 0.81 | 0.73 |
| 1PZ4 | 0.66 | 0.575 | 0.63 | |
| 3UQI | 0.66 | 0.45 | 0.59 | |
| 5,4′–di-hydroxy-7-methoxy-8-O-sulphate flavone (mariahine) | 1YIY | 0.64 | 0.83 | 0.60 |
| 1PZ4 | 0.64 | 0 | 0.41 | |
| 3UQI | 0.64 | 0.53 | 0.53 | |
| 5,3′–di-hydroxy-7,4′-dimethoxy-8-O-sulphate flavone (condadine) | 1YIY | 0.70 | 0.86 | 0.66 |
| 1PZ4 | 0.70 | 0 | 0.45 | |
| 3UQI | 0.70 | 0.38 | 0.54 | |
| Sitosterol-3-O-β-d-glucopyranoside | 1YIY | 0.71 | 0.91 | 0.68 |
| 1PZ4 | 0.71 | 0.76 | 0.65 | |
| 3UQI | 10.71 | 0.64 | 0.62 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernandes, D.A.; Barros, R.P.C.; Teles, Y.C.F.; Oliveira, L.H.G.; Lima, J.B.; Scotti, M.T.; Nunes, F.C.; Conceição, A.S.; Vanderlei de Souza, M.d.F. Larvicidal Compounds Extracted from Helicteres velutina K. Schum (Sterculiaceae) Evaluated against Aedes aegypti L. Molecules 2019, 24, 2315. https://doi.org/10.3390/molecules24122315
Fernandes DA, Barros RPC, Teles YCF, Oliveira LHG, Lima JB, Scotti MT, Nunes FC, Conceição AS, Vanderlei de Souza MdF. Larvicidal Compounds Extracted from Helicteres velutina K. Schum (Sterculiaceae) Evaluated against Aedes aegypti L. Molecules. 2019; 24(12):2315. https://doi.org/10.3390/molecules24122315
Chicago/Turabian StyleFernandes, Diégina A., Renata P. C. Barros, Yanna C. F. Teles, Louise H. G. Oliveira, Jéssica B. Lima, Marcus T. Scotti, Fabíola C. Nunes, Adilva S. Conceição, and Maria de Fátima Vanderlei de Souza. 2019. "Larvicidal Compounds Extracted from Helicteres velutina K. Schum (Sterculiaceae) Evaluated against Aedes aegypti L." Molecules 24, no. 12: 2315. https://doi.org/10.3390/molecules24122315