Differentiation of Taxonomically Closely Related Species of the Genus Acinetobacter Using Raman Spectroscopy and Chemometrics


Abstract
:1. Introduction
2. Results
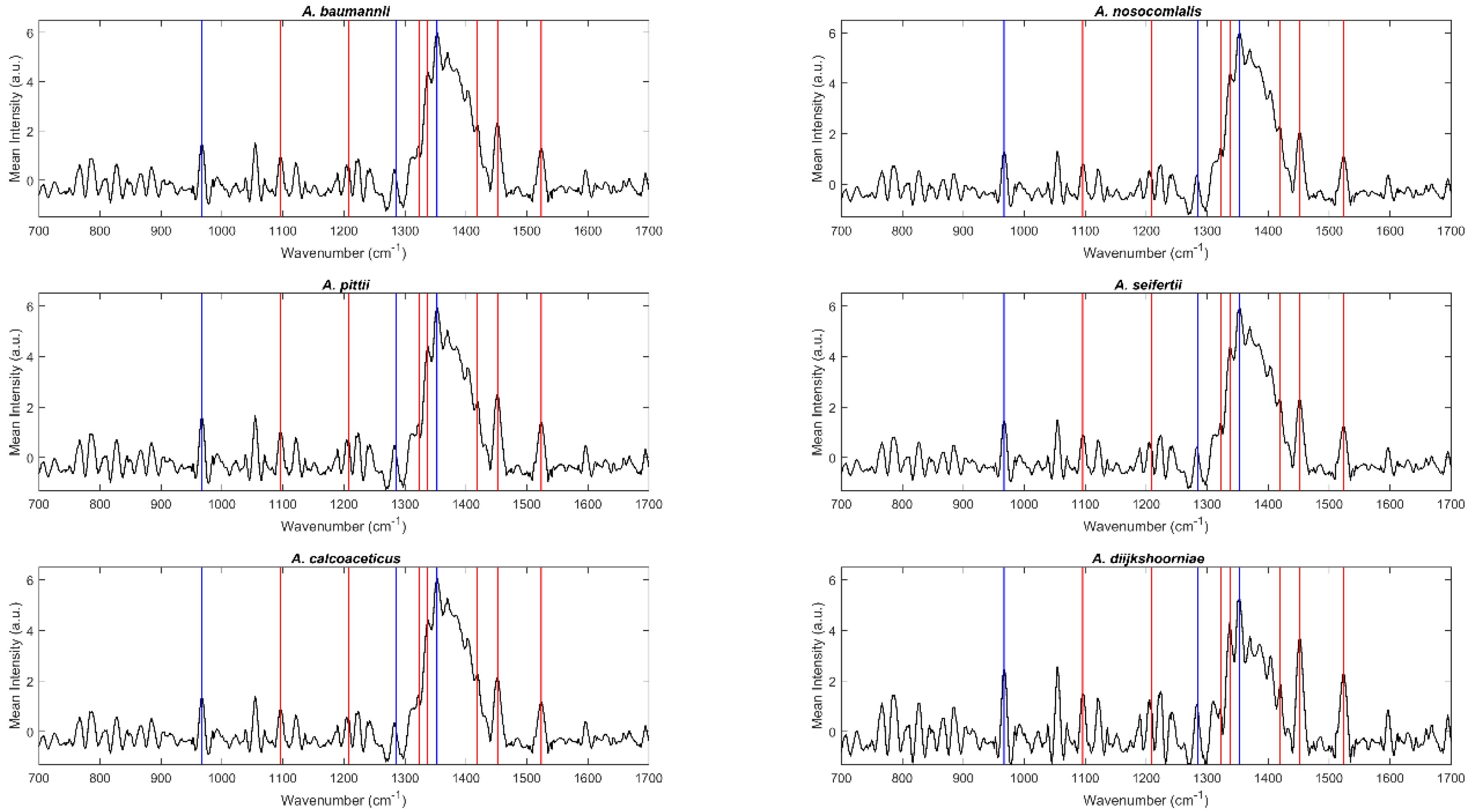
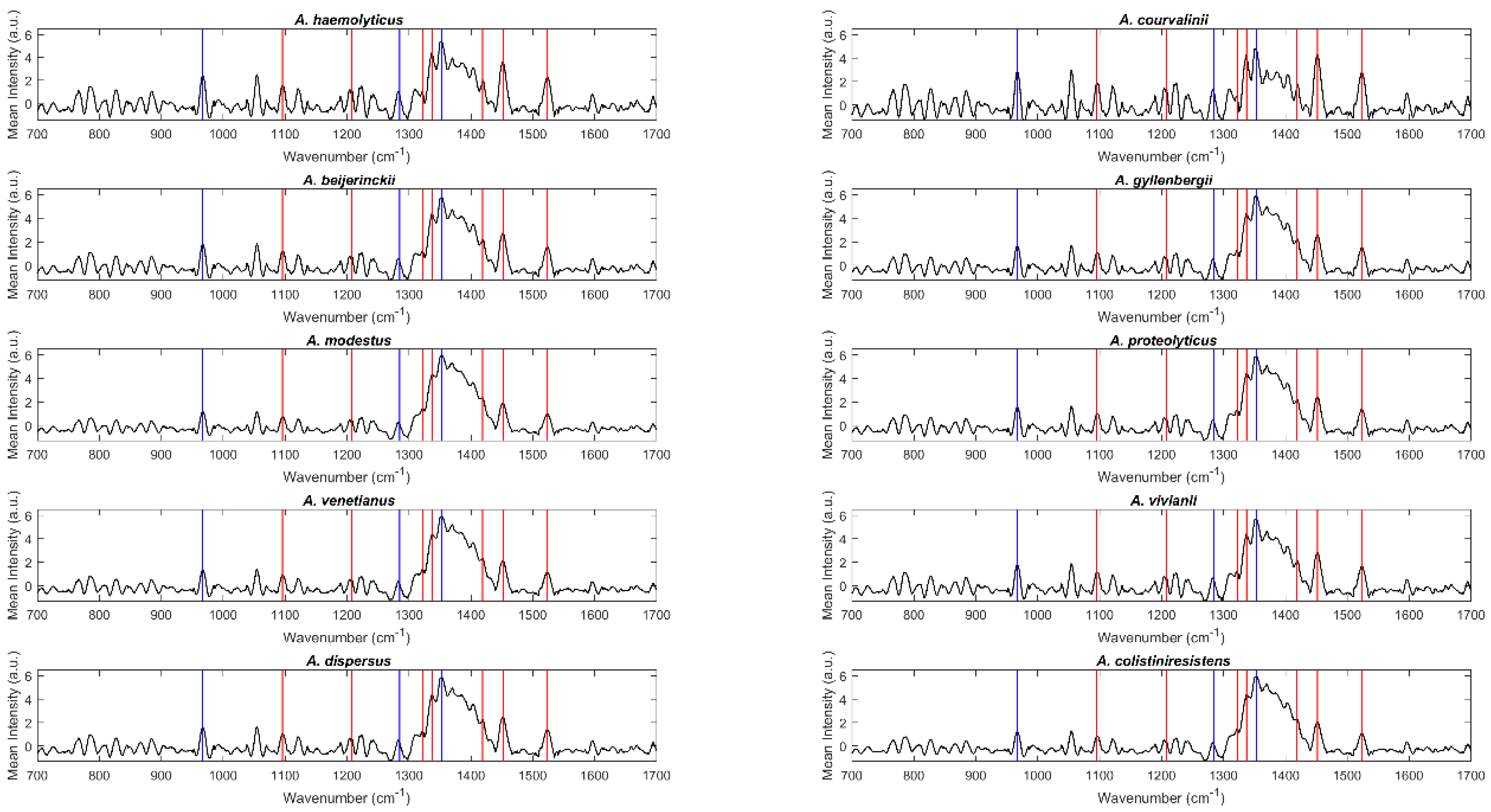
2.1. Spectral Analysis
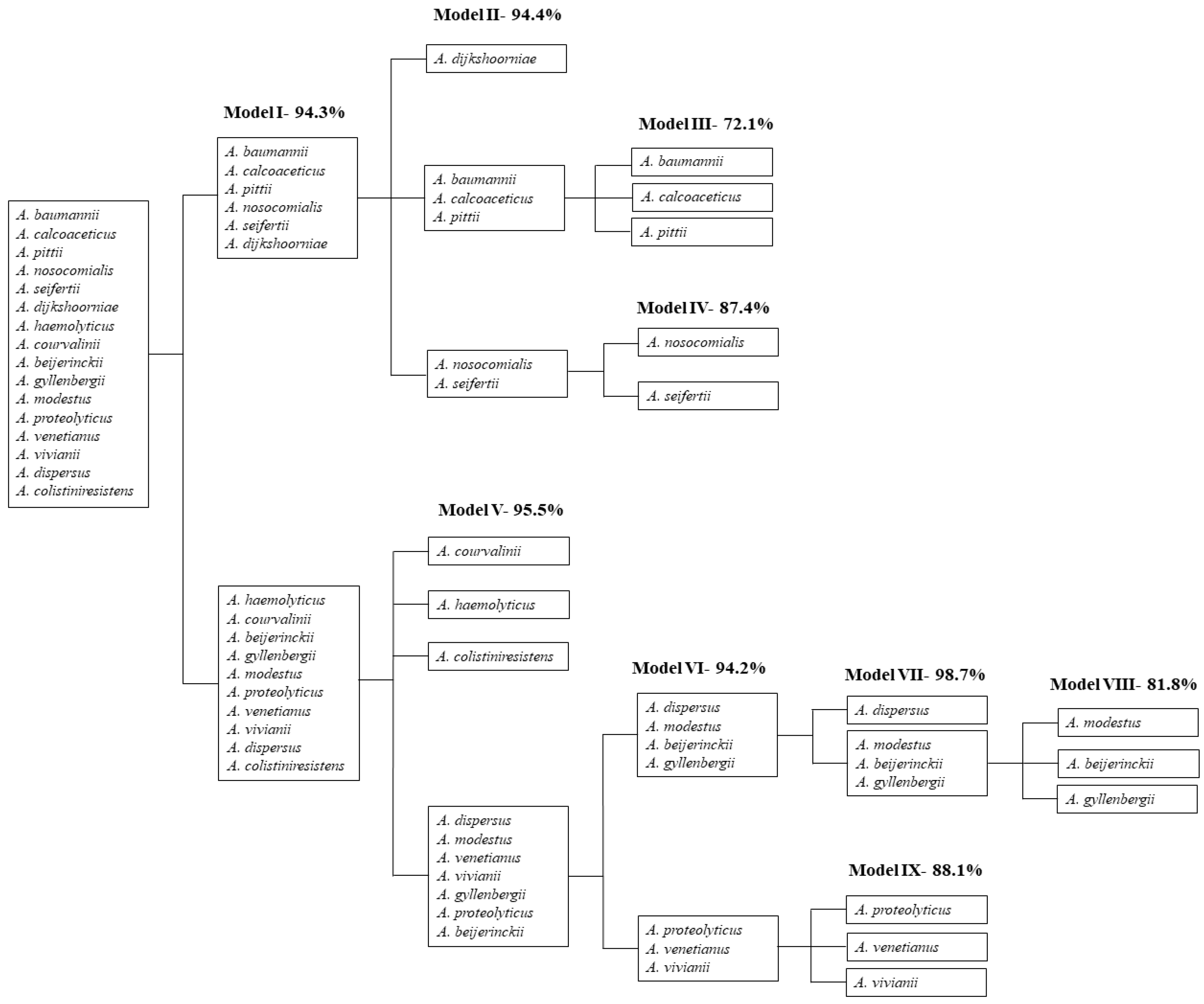
2.2. Discrimination of Acinetobacter Species
3. Discussion
4. Material and Methods
4.1. Bacterial Collection
4.2. Raman Spectroscopy
4.3. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Quintelas, C.; Ferreira, E.C.; Lopes, J.; Sousa, C. An overview of the evolution of infrared spectroscopy applied to bacterial typing. Biotechnol. J. 2018, 13. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Al-Qadiri, H.M.; Lin, M.; Rasco, B.A. Application of mid-infrared and Raman spectroscopy to the study of bacteria. Food Bioproc. Technol. 2011, 4, 919–935. [Google Scholar] [CrossRef]
- Vaz, M.; Meirinhos-Soares, L.; Sousa, C.; Ramirez, M.; Melo-Cristino, J.; Lopes, J. Serotypes discrimination of encapsulated Streptococcus pneumoniae strains by Fourier-transform infrared spectroscopy. J. Microbiol. Methods 2013, 93, 102–107. [Google Scholar] [CrossRef] [PubMed]
- van de Vossenberg, J.; Tervahauta, H.; Maquelin, K.; Blokker-Koopmans, C.H.W.; Uytewaal-Aarts, M.; van der Kooij, D.; van Wezel, A.P.; van der Gaag, B. Identification of bacteria in drinking water with Raman. Anal. Methods 2013, 5, 2679–2687. [Google Scholar] [CrossRef]
- Sousa, C.; Silva, L.; Grosso, F.; Nemec, A.; Lopes, J.; Peixe, L. Discrimination of the Acinetobacter calcoaceticus-Acinetobacter baumannii complex species by Fourier transform infrared spectroscopy. Eur. J. Clin. Microbiol. Infect. Dis. 2014, 33, 1345–1353. [Google Scholar] [CrossRef] [PubMed]
- Ghebremedhin, M.; Heitkamp, R.; Yesupriya, S.; Bradford, C.; Crane, N.J. Accurate and rapid differentiation of Acinetobacter baumannii strains by Raman spectroscopy: A comparative study. J. Clin. Microbiol. 2017, 55, 2480–2490. [Google Scholar] [CrossRef] [PubMed]
- Sousa, C.; Silva, L.; Grosso, F.; Lopes, J.; Peixe, L. Development of a FTIR-ATR based model for typing clinically relevant Acinetobacter baumannii clones belonging to ST98, ST103, ST208 and ST218. J. Photochem. Photobiol. B 2014, 133, 108–114. [Google Scholar] [CrossRef] [PubMed]
- Sousa, C.; Botelho, J.; Grosso, F.; Silva, L.; Lopes, J.; Peixe, L. Unsuitability of MALDI-TOF MS to discriminate Acinetobacter baumannii clones under our routine experimental conditions. Front. Microbiol. 2015, 6, 481. [Google Scholar] [CrossRef] [PubMed]
- Maquelin, K.; Dijkshoorn, L.; van der Reijdenb, T.J.K.; Puppels, G.J. Rapid epidemiological analysis of Acinetobacter strains by Raman spectroscopy. J. Microbiol. Methods 2006, 64, 126–131. [Google Scholar] [CrossRef]
- Dijkshoorn, L.; Nemec, A.; Seifert, H. An increasing threat in the hospital: Multidrug resistant Acinetobacter baumannii. Nat. Rev. Microbiol. 2007, 5, 939–951. [Google Scholar] [CrossRef]
- Cosgaya, C.; Marí-Almirall, M.; Van Assche, A.; Fernández-Orth, D.; Mosqueda, N.; Telli, M.; Huys, G.; Higgins, P.G.; Seifert, H.; Lievens, B.; et al. Acinetobacter dijkshoorniae sp. nov., a member of the Acinetobacter calcoaceticus-Acinetobacter baumannii complex mainly recovered from clinical samples in different countries. Int. J. Syst. Evol. Microbiol. 2016, 66, 4105–4111. [Google Scholar] [PubMed]
- Nemec, A.; Krizova, L.; Maixnerova, M.; van der Reijden, T.J.K.; Deschaght, P.; Passet, V.; Vaneechoutte, M.; Brisse, S.; Dijkshoorn, L. Genotypic and phenotypic characterization of the Acinetobacter calcoaceticus-Acinetobacter baumannii complex with the proposal of Acinetobacter pittii sp. nov. (formerly Acinetobacter genomic species 3) and Acinetobacter nosocomialis sp. nov. (formerly Acinetobacter genomic species 13TU). Res. Microbiol. 2011, 162, 393–404. [Google Scholar] [PubMed]
- Nemec, A.; Krizova, L.; Maixnerova, M.; Sedo, O.; Brisse, S.; Higgins, P.G. Acinetobacter seifertii sp. nov., a member of the Acinetobacter calcoaceticus-Acinetobacter baumannii complex isolated from human clinical specimens. Int. J. Syst. Evol. Microbiol. 2015, 65, 934–942. [Google Scholar] [CrossRef]
- Nemec, A.; Radolfova-Krizova, L.; Maixnerova, M.; Vrestiakova, E.; Jezek, P.; Sedo, O. Taxonomy of haemolytic and/or proteolytic strains of the genus Acinetobacter with the proposal of Acinetobacter courvalinii sp. nov. (genomic species 14 sensu Bouvet & Jeanjean), Acinetobacter dispersus sp. nov. (genomic species 17), Acinetobacter modestus sp. nov., Acinetobacter proteolyticus sp. nov. and Acinetobacter vivianii sp. nov. Int. J. Syst. Evol. Microbiol. 2016, 66, 1673–1685. [Google Scholar] [PubMed]
- Nemec, A.; Radolfova-Krizova, L.; Maixnerova, M.; Sedo, O. Acinetobacter colistiniresistens sp. nov. (formerly genomic species 13 sensu Bouvet and Jeanjean and genomic species 14 sensu Tjernberg and Ursing), isolated from human infections and characterised by intrinsic resistance to polymyxins. Int. J. Syst. Evol. Microbiol. 2017, 67, 2134–2141. [Google Scholar] [CrossRef] [PubMed]
- Šedo, O.; Nemec, A.; Křížová, L.; Kačalová, M.; Zdráhal, Z. Improvement of MALDI-TOF MS profiling for the differentiation of species within the Acinetobacter calcoaceticus-Acinetobacter baumannii complex. Syst. Appl. Microbiol. 2013, 36, 572–578. [Google Scholar] [CrossRef] [PubMed]
- Šedo, O.; Radolfová-Křížová, L.; Nemec, A.; Zdráhal, Z. Limitations of routine MALDI-TOF mass spectrometric identification of Acinetobacter species and remedial actions. J. Microbiol. Methods 2018, 154, 79–85. [Google Scholar] [CrossRef]
- de Oliveira, F.S.S.; Giana, H.E.; Silveira, L. Discrimination of selected species of pathogenic bacteria using near-infrared Raman spectroscopy and principal components analysis. J. Biomed. Opt. 2012, 17. [Google Scholar] [CrossRef]
- Paret, M.L.; Sharma, S.K.; Green, L.M.; Alvarez, A.M. Biochemical characterization of Gram-positive and Gram-negative plant-associated bacteria with micro-Raman spectroscopy. Appl. Spectrosc. 2010, 64, 433–441. [Google Scholar] [CrossRef]
- Berger, A.J.; Zhu, Q. Identification of oral bacteria by Raman microspectroscopy. J. Mod. Optic. 2013, 50, 15–17. [Google Scholar] [CrossRef]
- Sengupta, A.; Mujacic, M.; Davis, E.J. Detection of bacteria by surface-enhanced Raman spectroscopy. Anal. Bioanal. Chem. 2006, 386, 1379–1386. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.Y.; Chen, Y.; Wang, H.H.; Huang, Y.L.; Chao, Y.C.; Tsai, K.T.; Cheng, W.C.; Chuang, C.Y.; Tsai, Y.H.; Huang, C.Y.; et al. Differentiation of bacteria cell wall using Raman scattering enhanced by nanoparticle array. J. Nanosci. Nanotechnol. 2012, 12, 5004–5008. [Google Scholar] [CrossRef] [PubMed]
- Sousa, C.; Botelho, J.; Silva, L.; Grosso, F.; Nemec, A.; Lopes, J.; Peixe, L. MALDI-TOF MS and chemometric based identification of Acinetobacter calcoaceticus-Acinetobacter baumannii complex species. Int. J. Med. Microbiol. 2014, 304, 669–677. [Google Scholar] [CrossRef] [PubMed]
- Nemec, A.; Radolfova-Krizova, L.; Maixnerova, M.; Nemec, M.; Clermont, D.; Bzdil, J.; Jezek, P.; Spanelova, P. Revising the taxonomy of the Acinetobacter lwoffii group: The description of Acinetobacter pseudolwoffii sp. nov. and emended description of Acinetobacter lwoffii. Syst. Appl. Microbiol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Geladi, P.; Kowalsky, B.R. Partial least-squares regression: A tutorial. Anal. Chim. Acta 1986, 185, 1–17. [Google Scholar] [CrossRef]
- Alsberg, B.K.; Kell, D.B.; Goodacre, R. Variable selection in discriminant partial least-squares analysis. Anal. Chem. 1998, 70, 4126–4133. [Google Scholar] [CrossRef] [PubMed]
- Sousa, C.; Grosso, F.; Meirinhos-Soares, L.; Peixe, L.; Lopes, J. Identification of carbapenem-resistant Acinetobacter baumannii clones using infrared spectroscopy. J. Biophotonics 2014, 7, 287–294. [Google Scholar] [CrossRef] [PubMed]
- Nemec, A.; Musílek, M.; Maixnerová, M.; De Baere, T.; van der Reijden, T.J.K.; Vaneechoutte, M.; Dijkshoorn, L. Acinetobacter beije-rinckii sp. nov. and Acinetobacter gyllenbergii sp. nov., haemolytic organisms isolated from humans. Int. J. Syst. Evol. Microbiol. 2009, 59, 118–124. [Google Scholar] [CrossRef]
- Vaneechoutte, M.; Nemec, A.; Musílek, M.; van der Reijden, T.J.K.; van den Barselaar, M.; Tjernberg, I.; Calame, W.; Fani, R.; De Baere, T.; Dijkshoorn, L. Description of Acinetobacter venetianus ex Di Cello et al. 1997 sp. nov. Int. J. Syst. Evol. Microbiol. 2009, 59, 1376–1381. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are not available from the authors. |




{kind=link}
{kind=link}
{kind=link}
| PLSDA Model | Spectra Pre-Processing * | Number of Latent Variables (LVs) | Clusters | Species | % of Correct Assignments |
|---|---|---|---|---|---|
| A | A. baumannii | ||||
| A. calcoaceticus | |||||
| A. pittii | |||||
| A. nosocomialis | |||||
| A. seifertii | |||||
| I | 5 | A. dijkshoorniae | 94.3% | ||
| B | A. haemolyticus | ||||
| A. courvalinii | |||||
| A. beijerinckii | |||||
| A. gyllenbergii | |||||
| A. modestus | |||||
| A. proteolyticus | |||||
| A. venetianus | |||||
| A. vivianii | |||||
| A. dispersus | |||||
| A. colistiniresistens |
| PLSDA Model | Spectra Pre-Processing * | Number of LVs | Clusters ** | Species | % of Correct Assignments |
|---|---|---|---|---|---|
| II-cluster A isolates | --- | 6 | 1 | A. dijkshoorniae | 94.4% |
| AI | A. baumannii | ||||
| A. calcoaceticus | |||||
| A. pittii | |||||
| AII | A. nosocomialis | ||||
| A. seifertii | |||||
| III-cluster AI isolates | --- | 5 | 1 | A. baumannii | 72.1% |
| 2 | A. calcoaceticus | ||||
| 3 | A. pittii | ||||
| IV-cluster AII isolates | --- | 5 | 1 | A. nosocomialis | 87.4% |
| 2 | A. seifertii |
| PLSDA Model | Spectra Pre-Processing * | Number of LVs | Clusters ** | Species | % of Correct Assignments |
|---|---|---|---|---|---|
| V-cluster B isolates | Standard normal variate (SNV) + Savitzky-Golay (SavGol) (7,2,1) | 6 | 1 | A. courvalinii | 95.5% |
| 2 | A. haemolyticus | ||||
| 3 | A. colistiniresistens | ||||
| BI | A. dispersus | ||||
| A. modestus | |||||
| A. venetianus | |||||
| A. vivianii | |||||
| A. gyllenbergii | |||||
| A. proteolyticus | |||||
| A. beijerinckii | |||||
| VI-cluster BI isolates | --- | 4 | BI-A | A. dispersus | 94.2% |
| A. modestus | |||||
| A. beijerinckii | |||||
| A. gyllenbergii | |||||
| BI-B | A. proteolyticus | ||||
| A. venetianus | |||||
| A. vivianii | |||||
| VII-cluster BI-A isolates | SNV + SavGol (7,2,1) | 5 | 1 | A. dispersus | 98.7% |
| BI-AII | A. modestus | ||||
| A. beijerinckii | |||||
| A. gyllenbergii | |||||
| VIII-cluster BI-AII isolates | --- | 4 | 1 | A. modestus | 81.8% |
| 2 | A. beijerinckii | ||||
| 3 | A. gyllenbergii | ||||
| VIX-cluster BI-B isolates | SNV + SavGol (7,2,1) | 4 | 1 | A. proteolyticus | 88.1% |
| 2 | A. venetianus | ||||
| 3 | A. vivianii |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Teixeira, A.M.; Nemec, A.; Sousa, C. Differentiation of Taxonomically Closely Related Species of the Genus Acinetobacter Using Raman Spectroscopy and Chemometrics. Molecules 2019, 24, 168. https://doi.org/10.3390/molecules24010168
Teixeira AM, Nemec A, Sousa C. Differentiation of Taxonomically Closely Related Species of the Genus Acinetobacter Using Raman Spectroscopy and Chemometrics. Molecules. 2019; 24(1):168. https://doi.org/10.3390/molecules24010168
Chicago/Turabian StyleTeixeira, A. Margarida, Alexandr Nemec, and Clara Sousa. 2019. "Differentiation of Taxonomically Closely Related Species of the Genus Acinetobacter Using Raman Spectroscopy and Chemometrics" Molecules 24, no. 1: 168. https://doi.org/10.3390/molecules24010168
APA StyleTeixeira, A. M., Nemec, A., & Sousa, C. (2019). Differentiation of Taxonomically Closely Related Species of the Genus Acinetobacter Using Raman Spectroscopy and Chemometrics. Molecules, 24(1), 168. https://doi.org/10.3390/molecules24010168