Effective Tetradentate Compound Complexes against Leishmania spp. that Act on Critical Enzymatic Pathways of These Parasites

 , ,
, ,
Abstract
:1. Introduction
2. Results
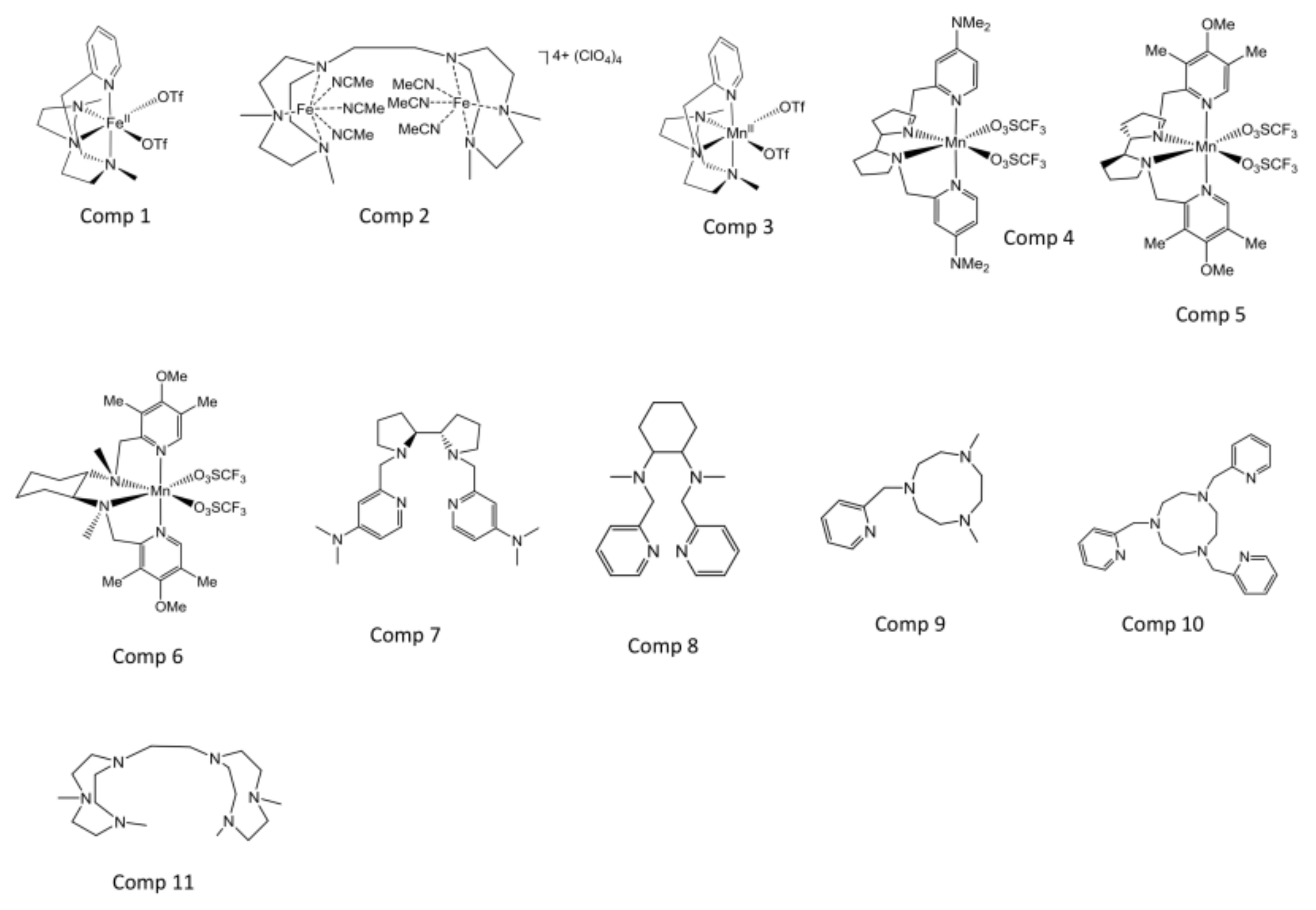
2.1. Selection of the Compounds Under Study
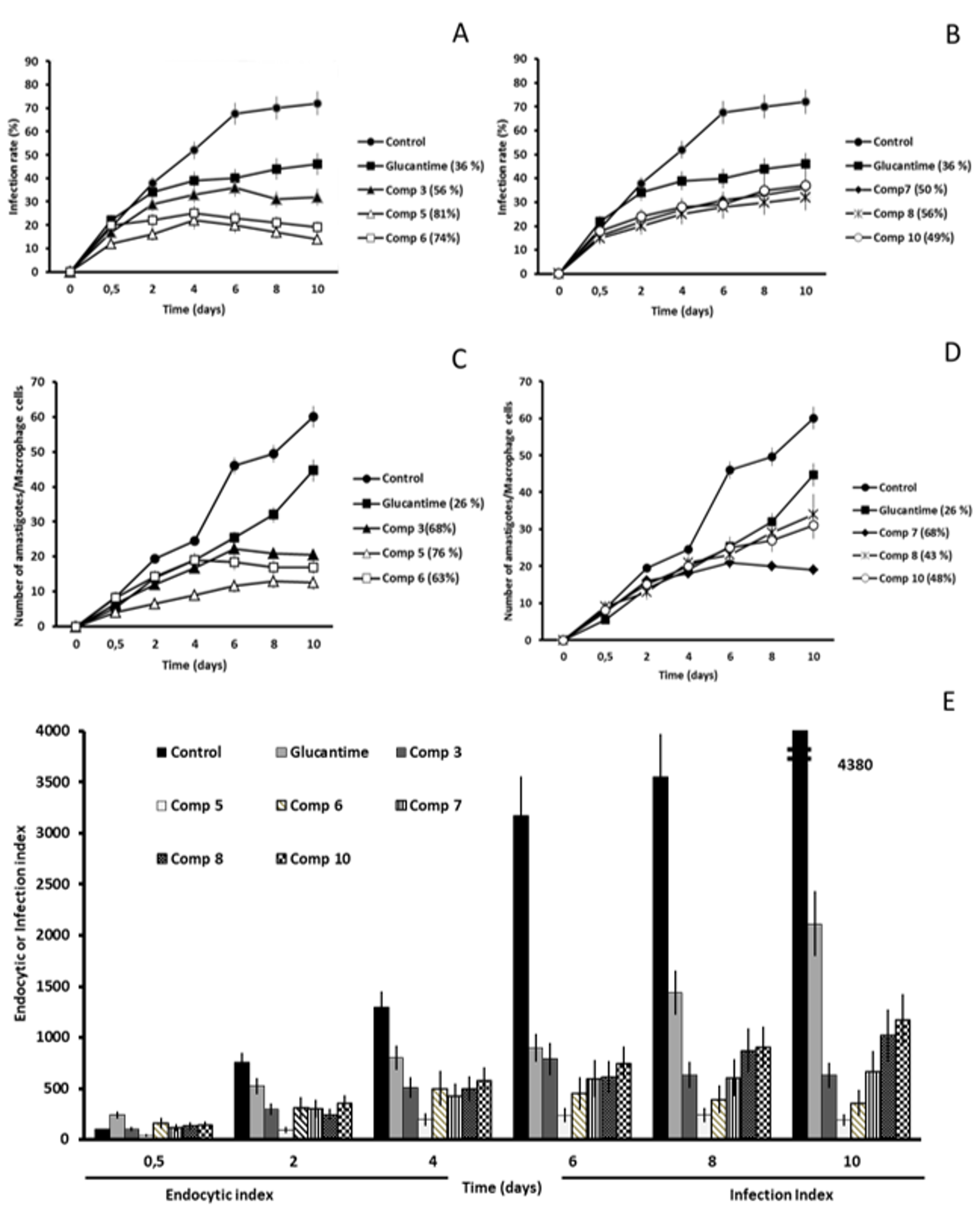
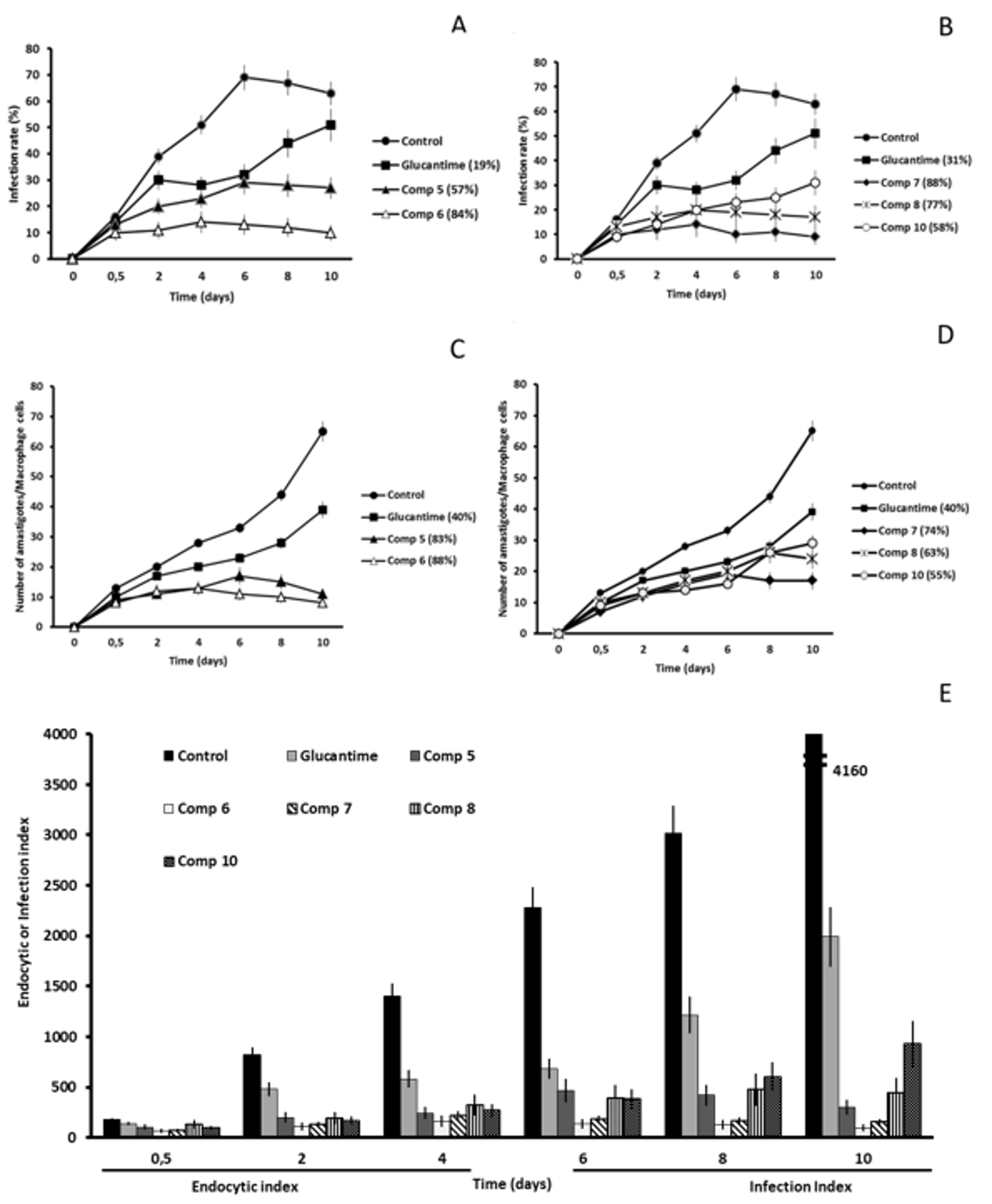
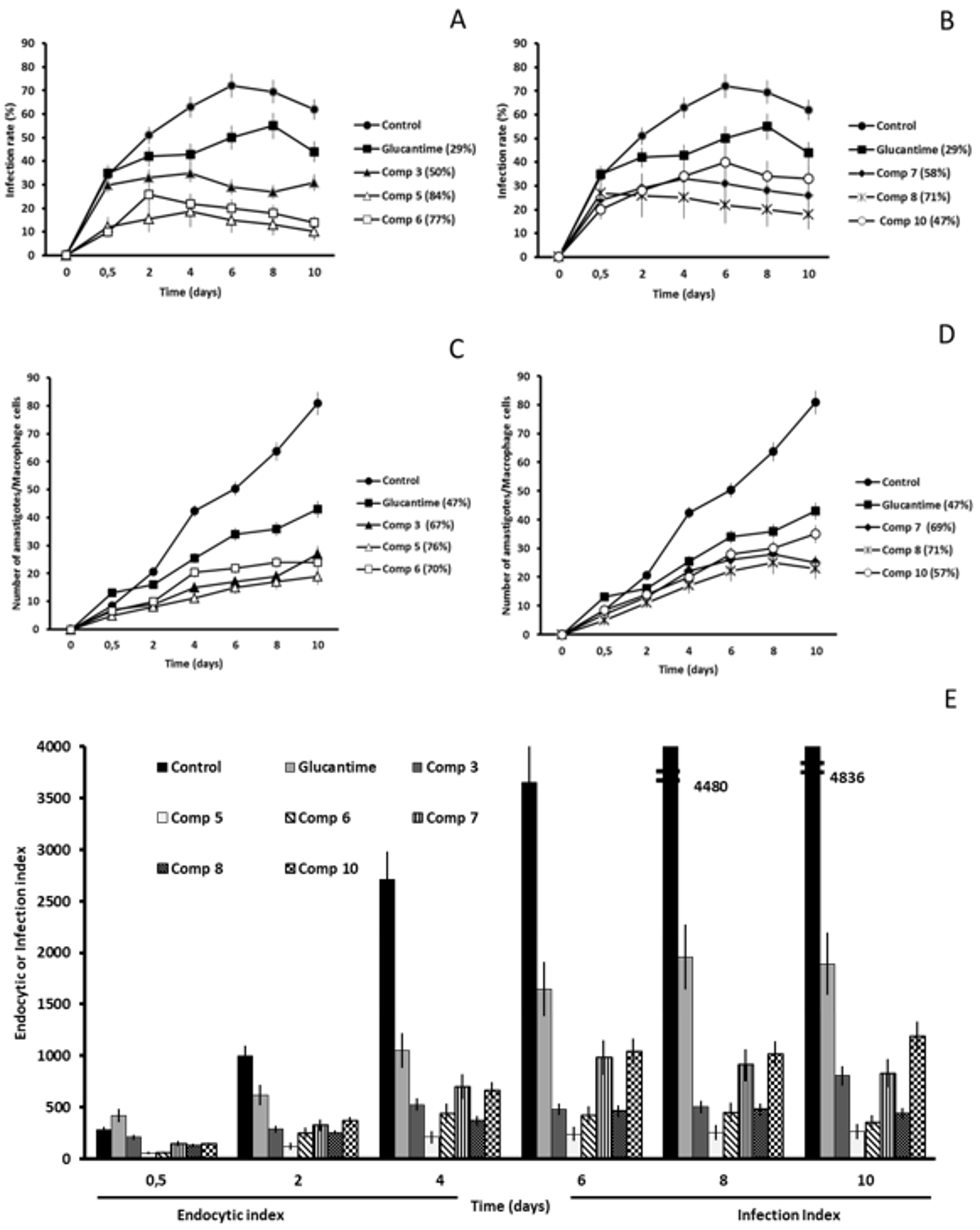
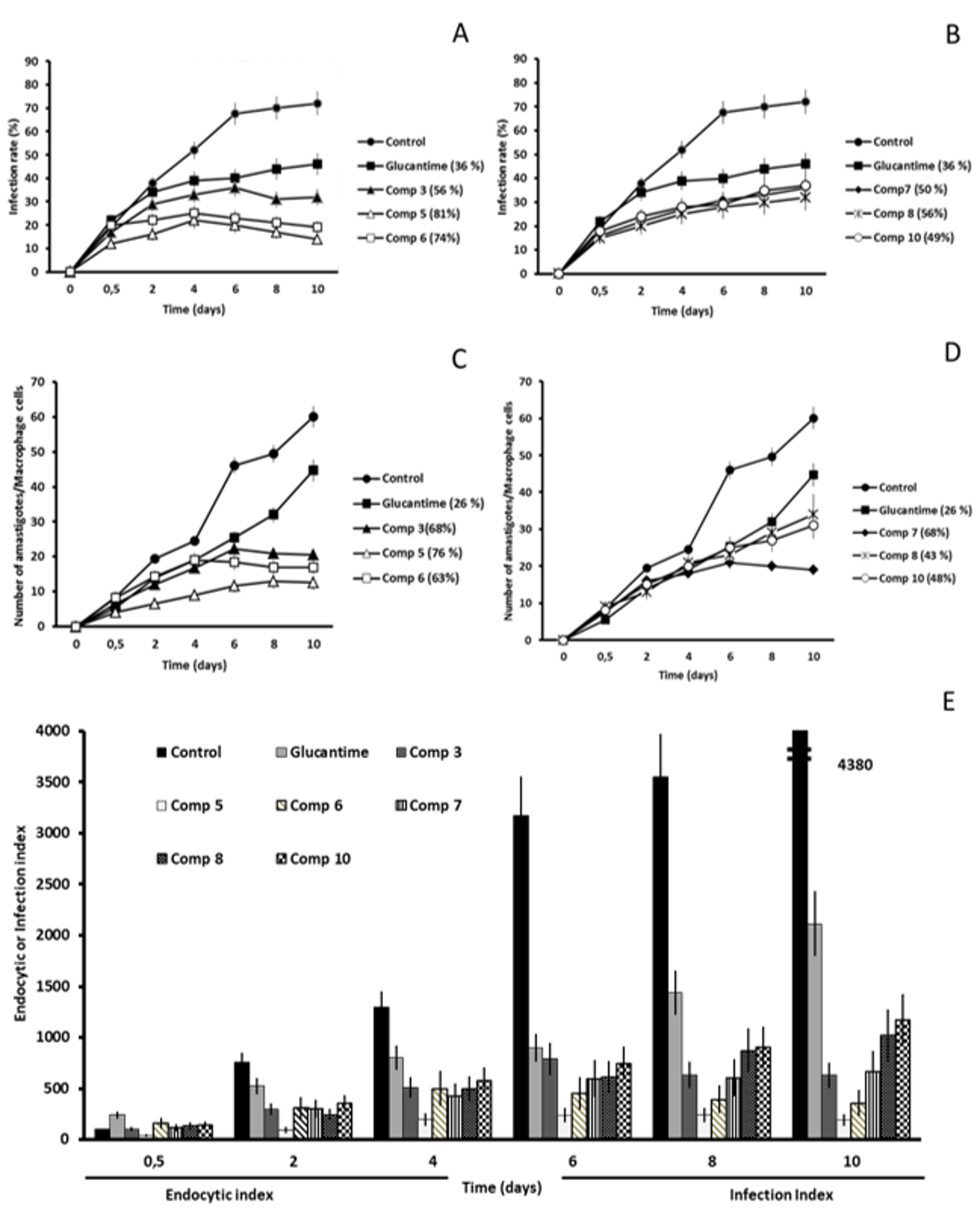
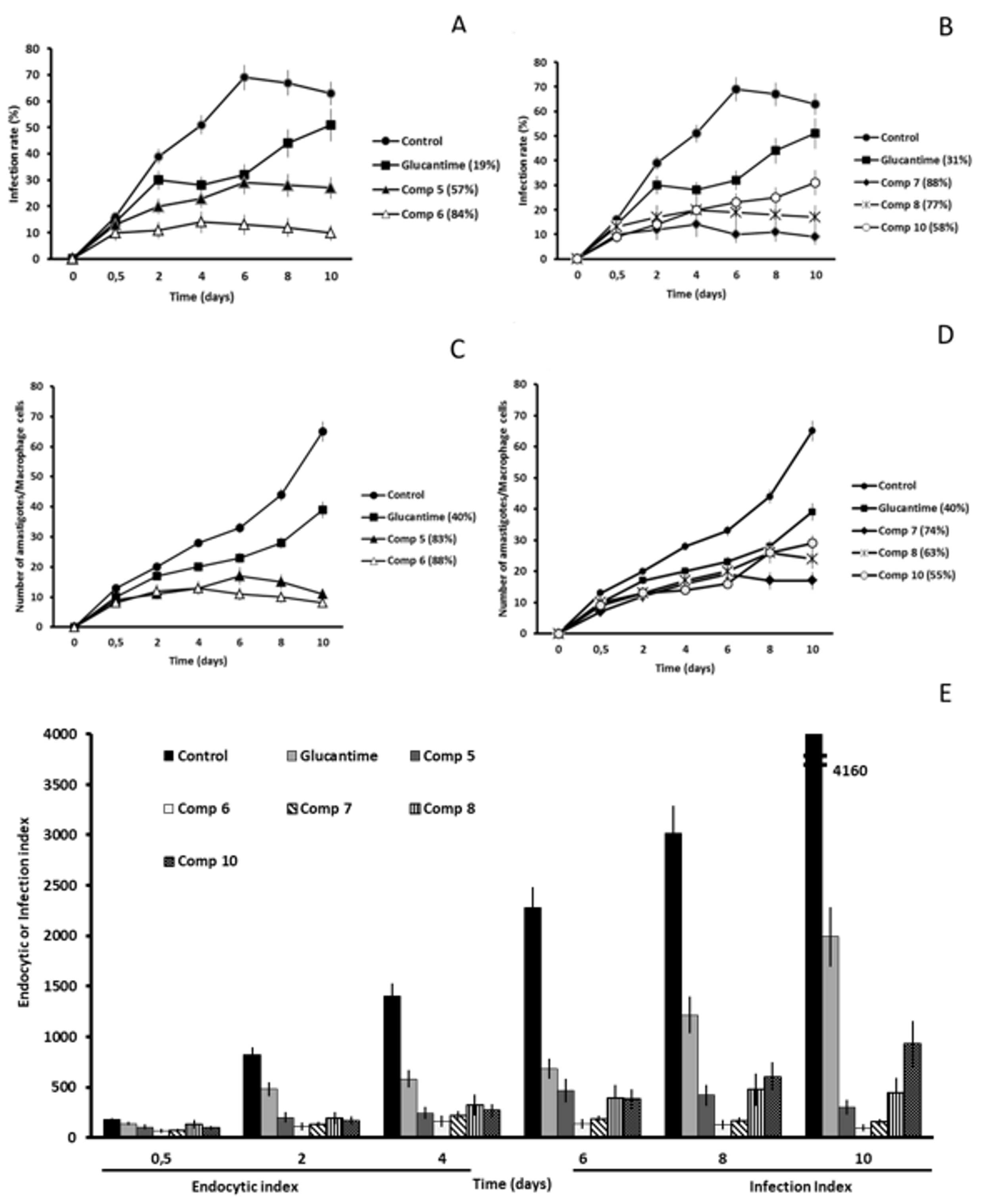
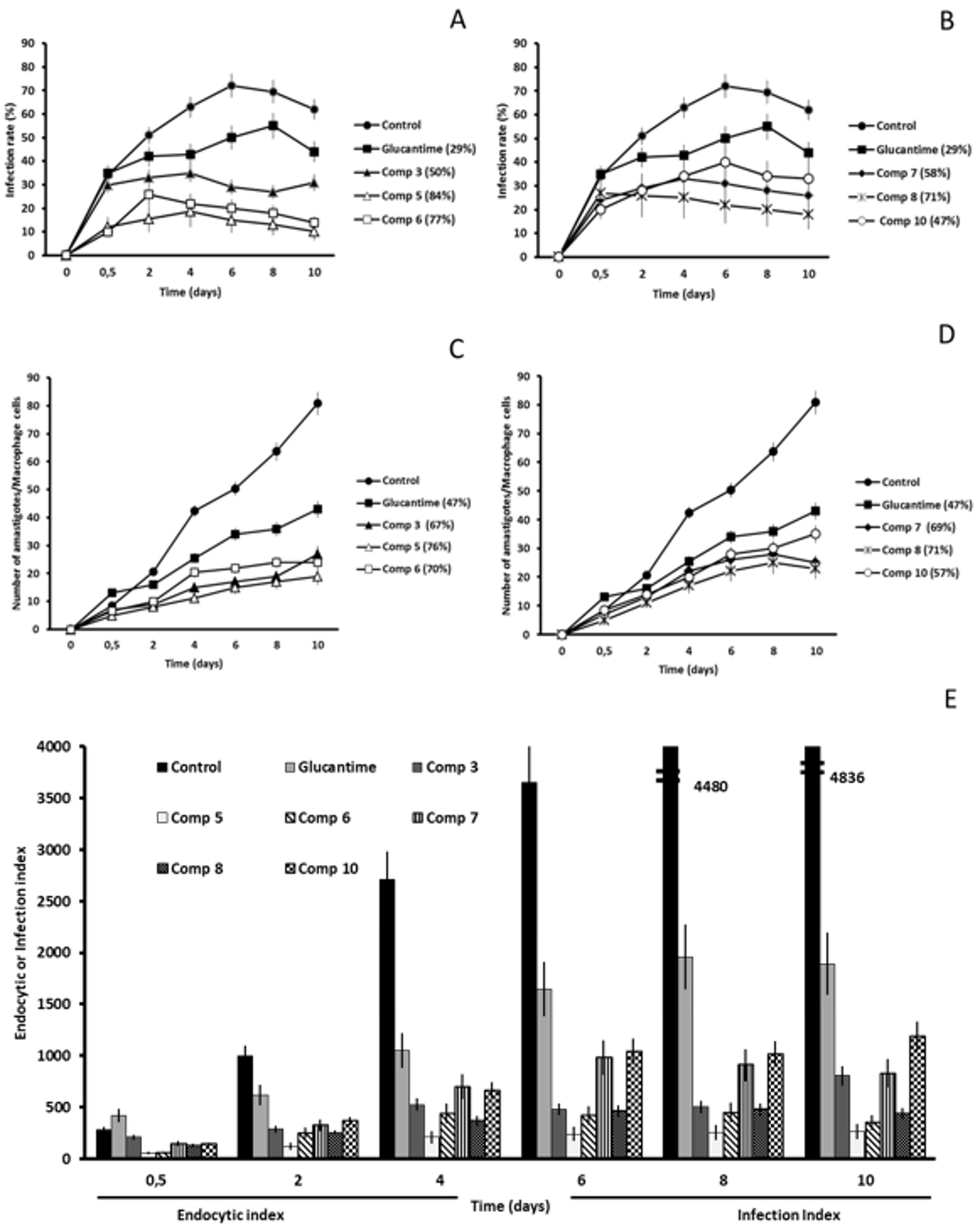
2.2. In Vitro Antileishmanial Activity
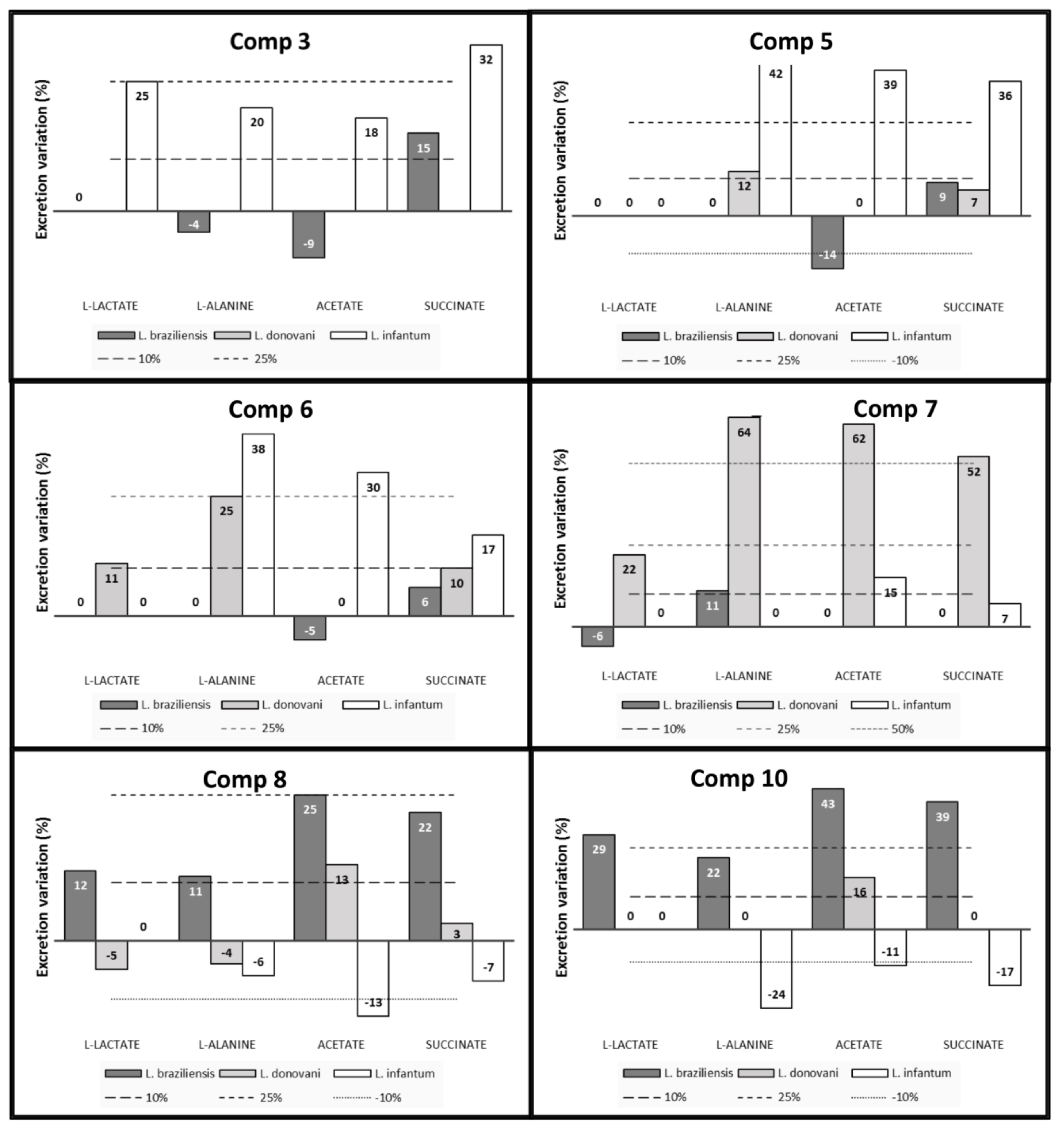
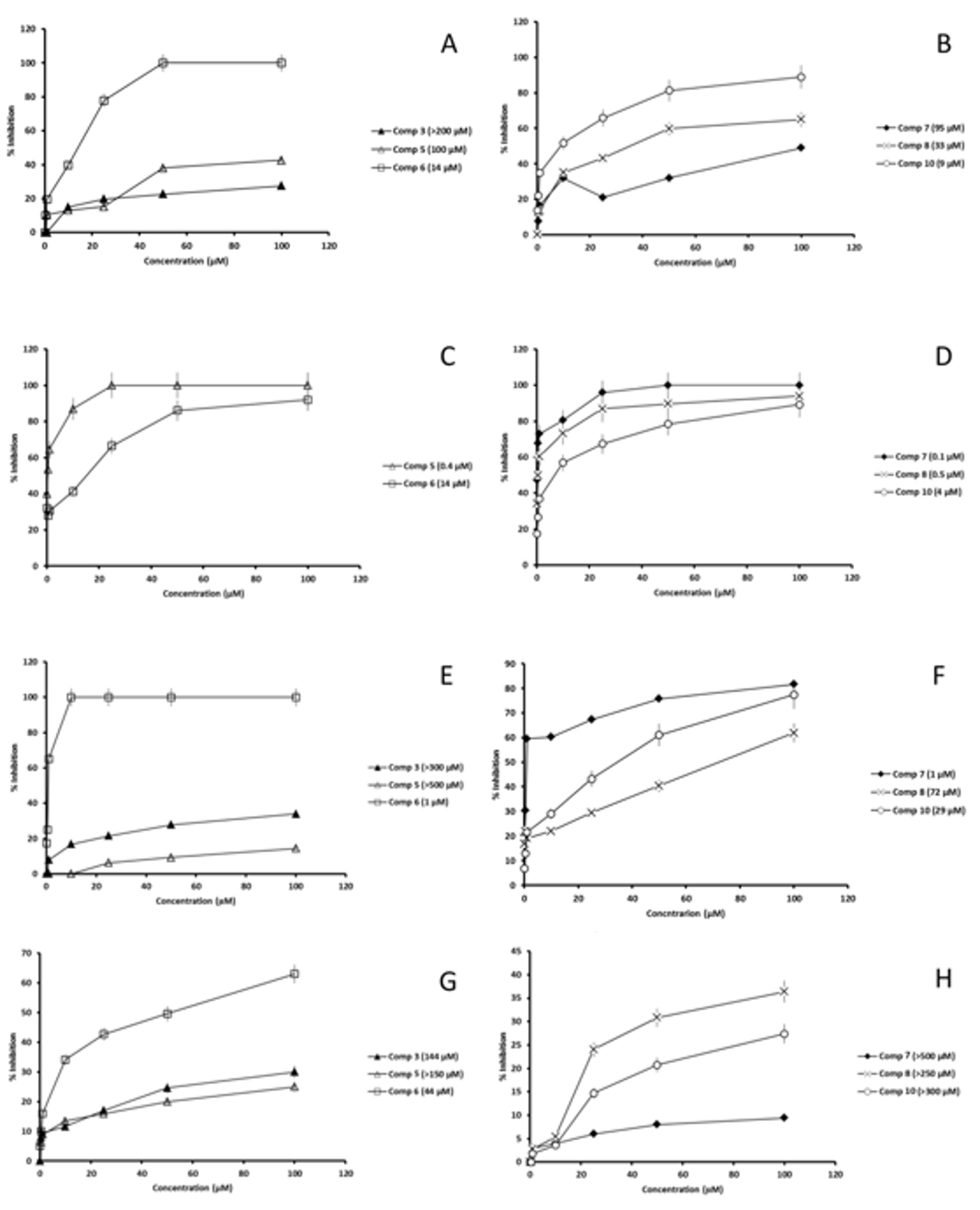
2.3. Studies on the Mechanism of Action
2.3.1. SOD Inhibition
2.3.2. 1H-NMR
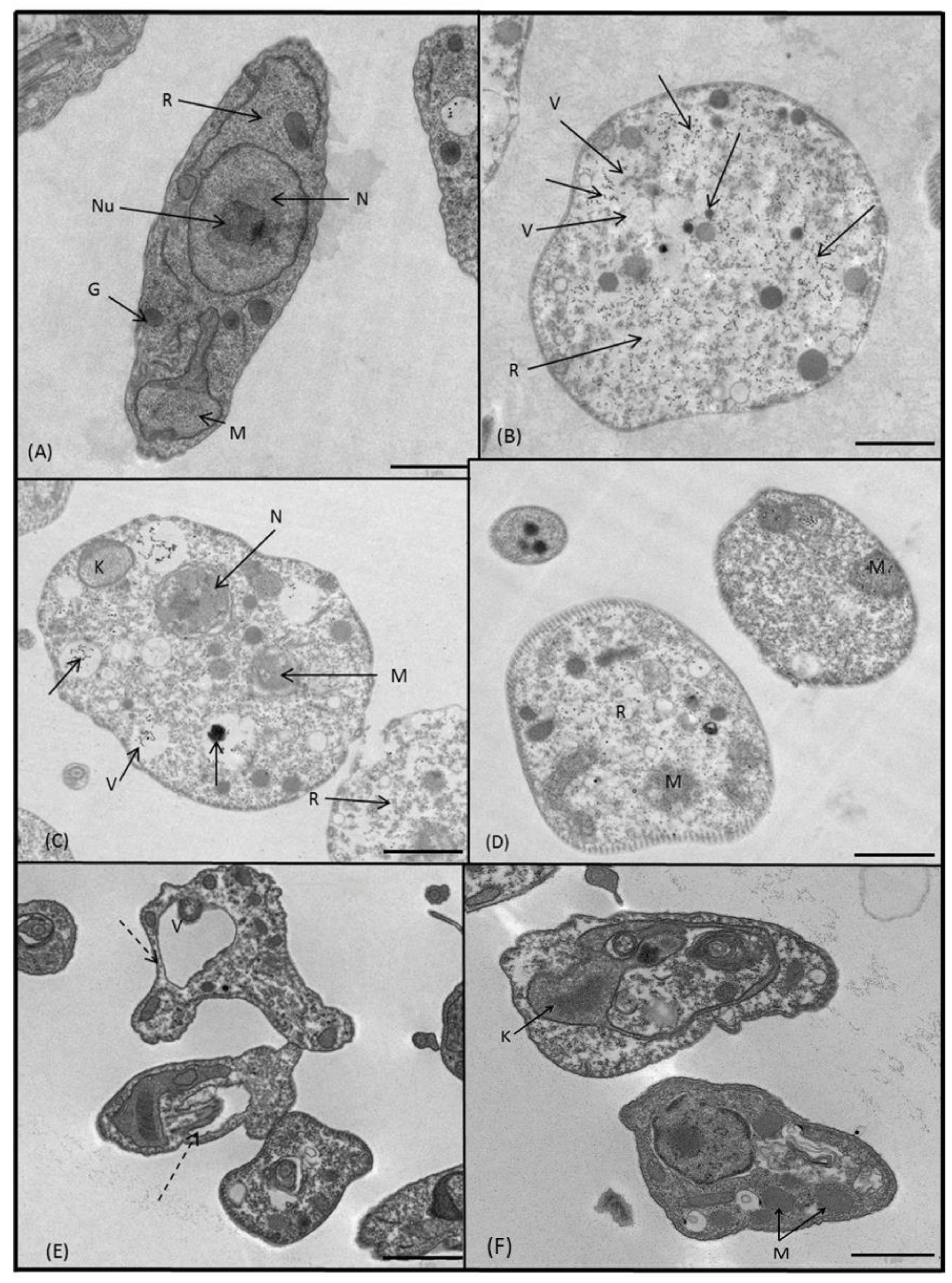
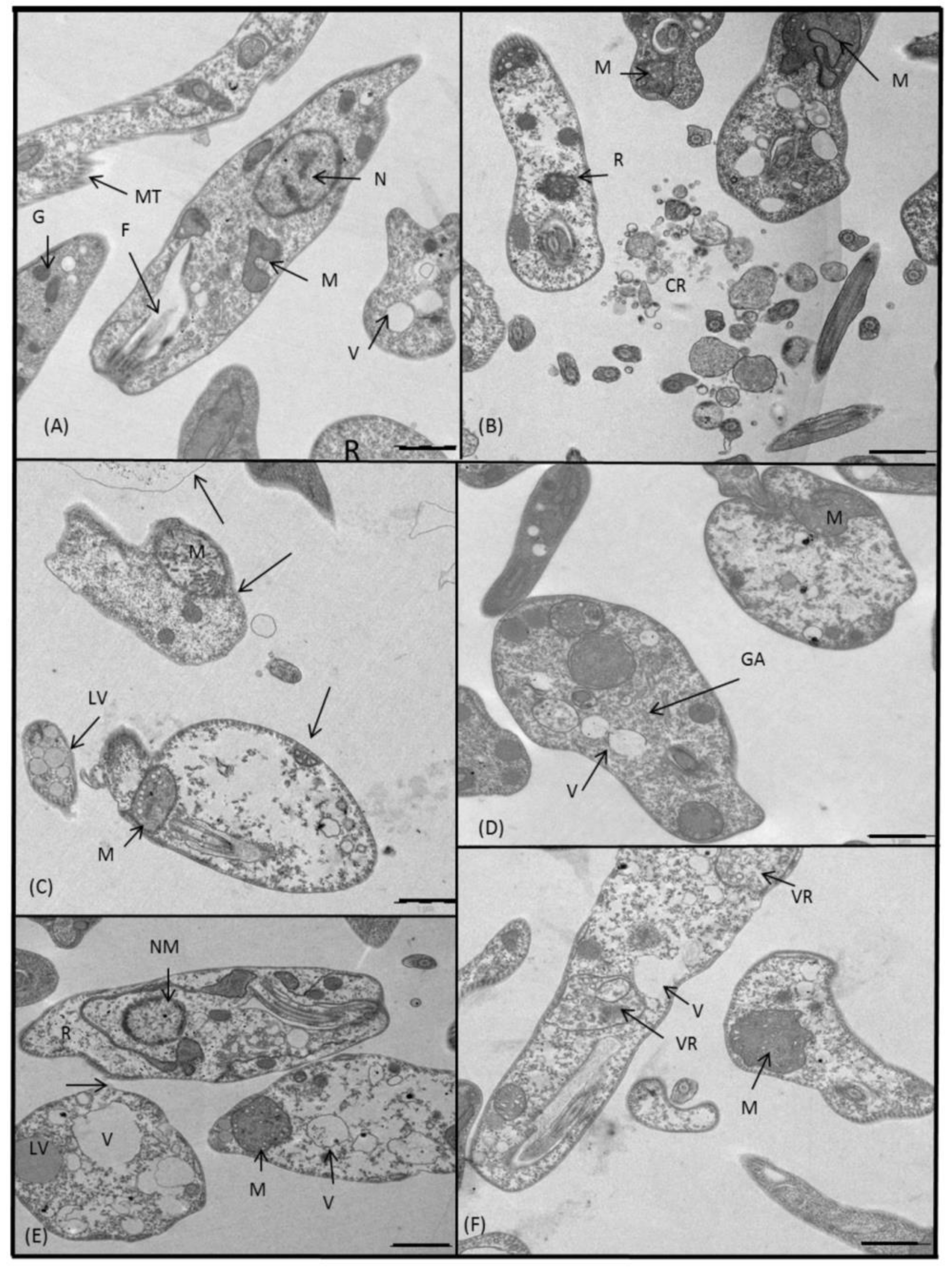
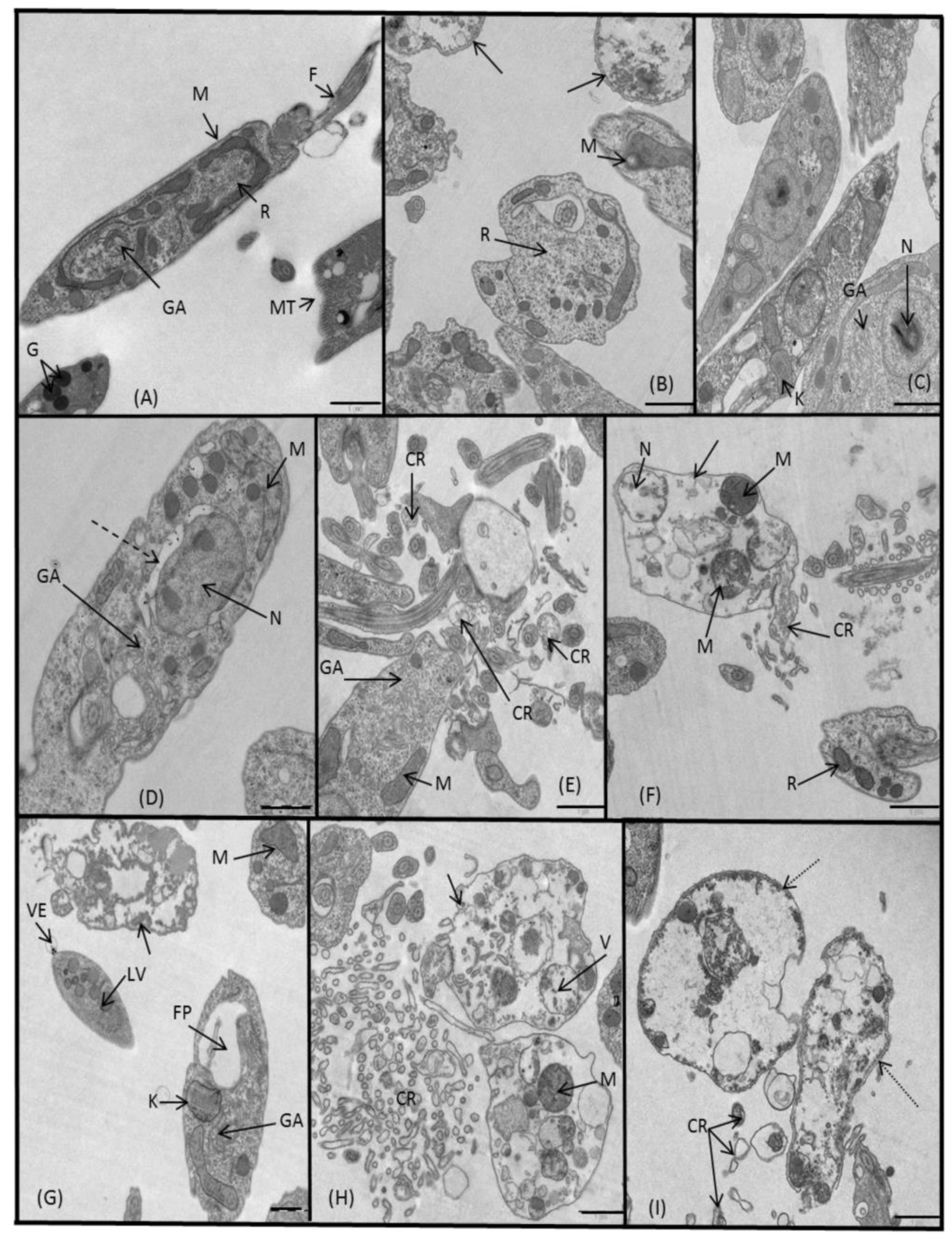
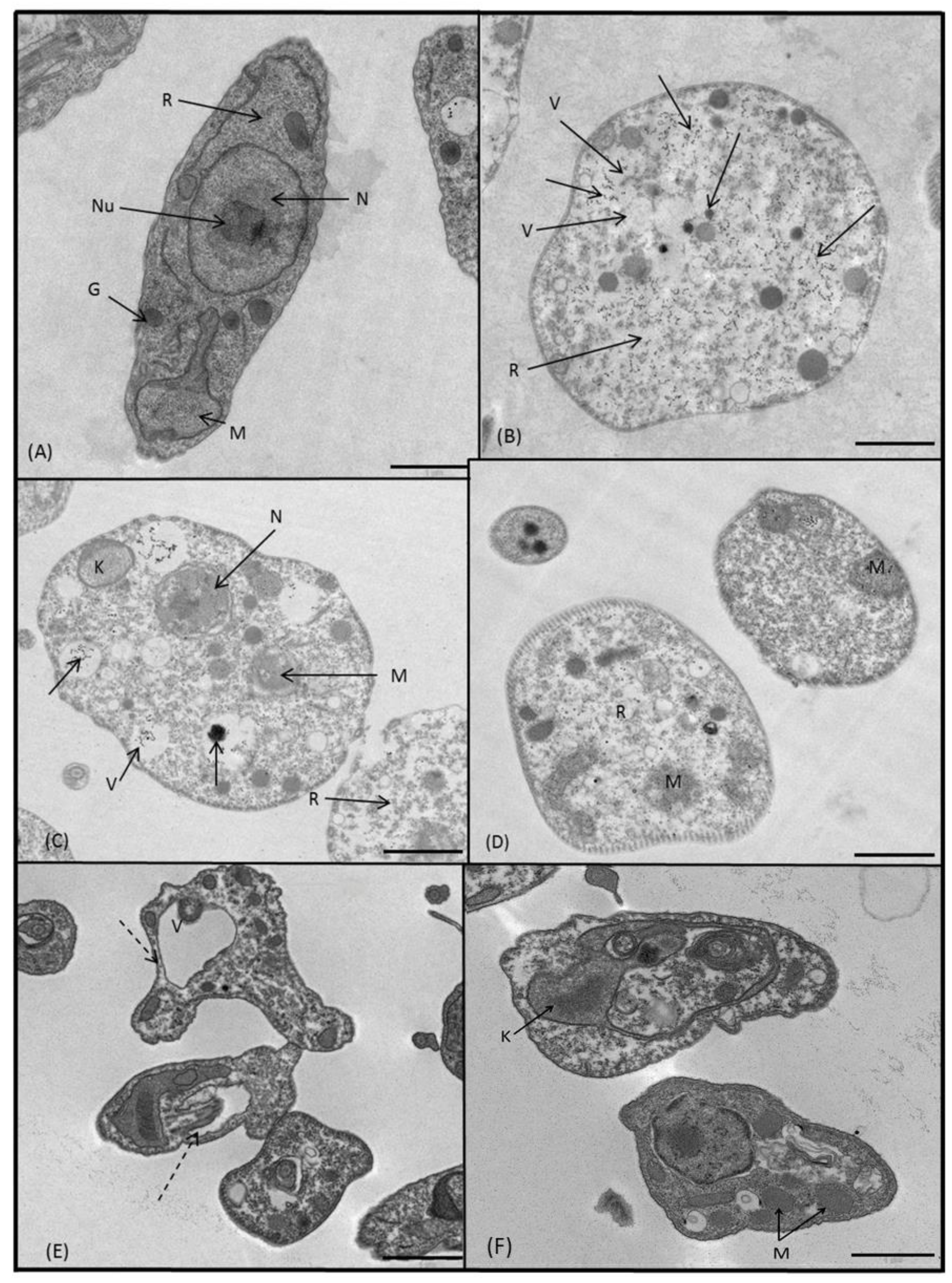
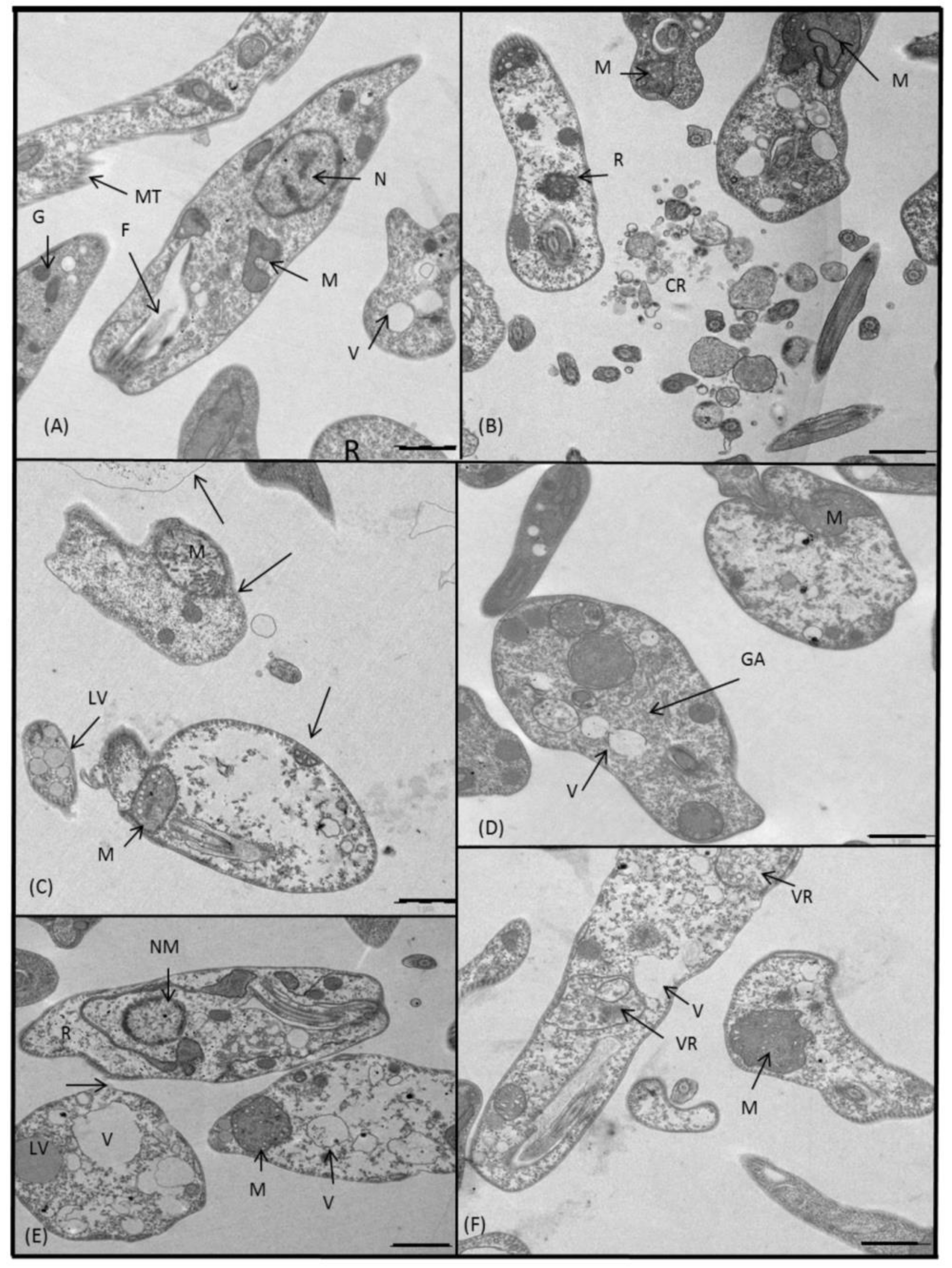
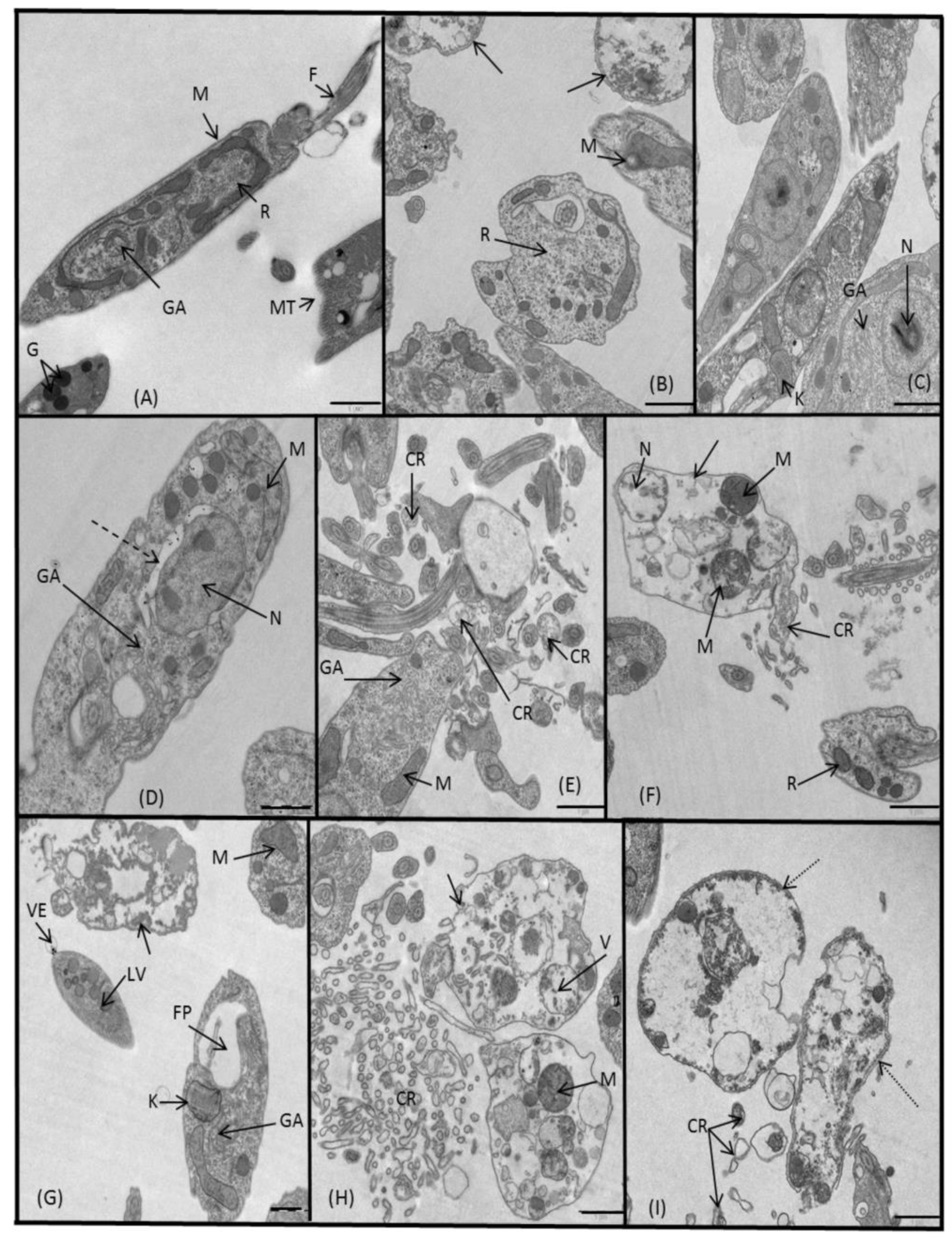
2.3.3. Microscopy
3. Discussion
4. Materials and Methods
4.1. Compounds Under Study
Synthesis of Compound 2
4.2. Parasite Strain and Culture
4.3. In Vitro Screening Assays
4.3.1. Cell Culture and Cytotoxicity Tests
4.3.2. Promastigote Tests: Extracellular Forms
4.3.3. Amastigote Tests: Intracellular Forms
4.4. Infectivity Assay
4.5. Ultrastructural Alterations
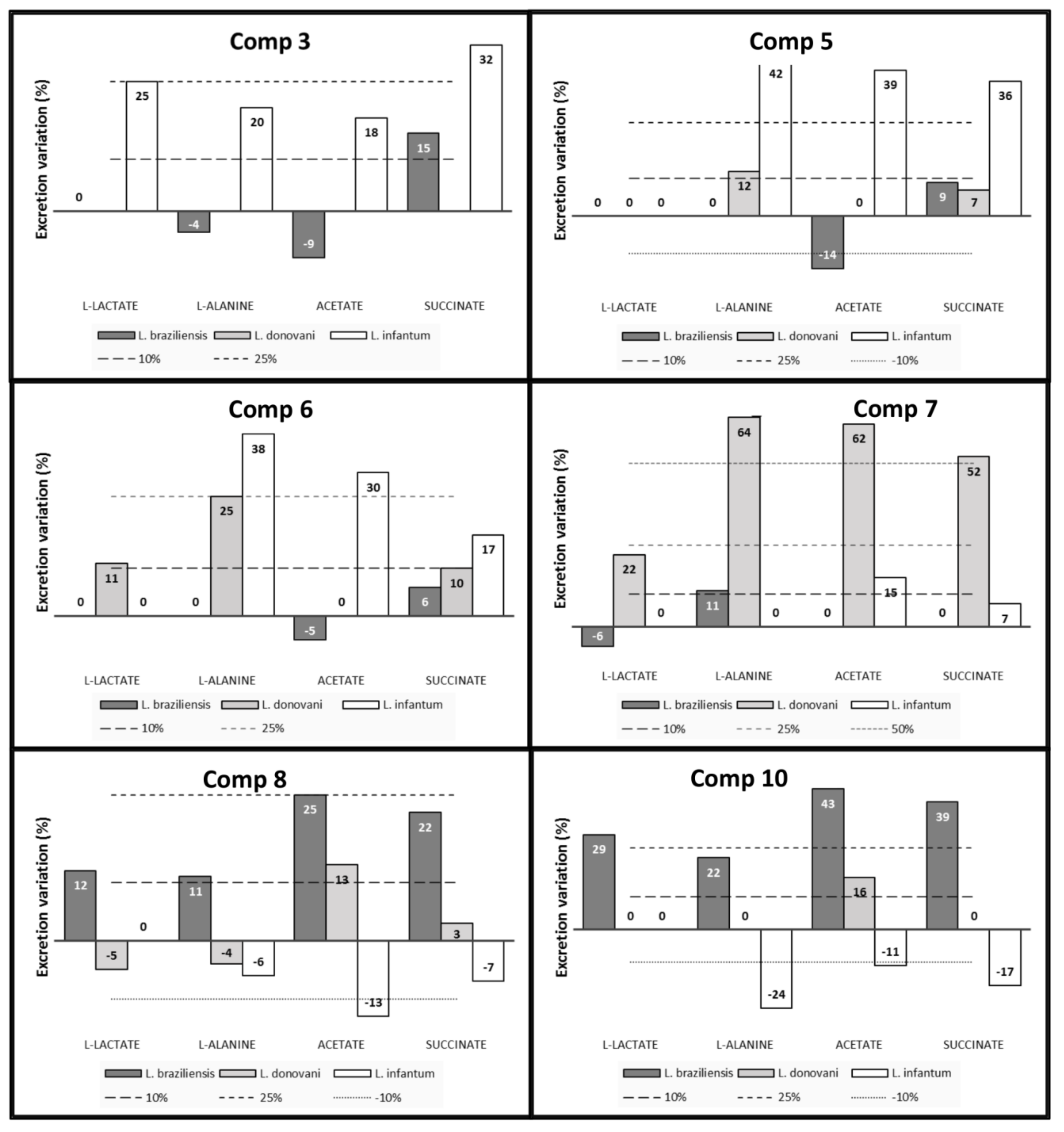
4.6. Metabolite Excretion
4.7. Fe-SOD Enzymatic Inhibition
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Available online: http://www.who.int/news-room/fact-sheets/detail/leishmaniasis (accessed on 11 October 2018).
- Mishra, J.; Dev, A.; Singh, N.; Somvanshi, R.; Singh, S. Evaluation of toxicity and therapeutic efficacy of a new liposomal formulation of amphotericin B in a mouse model. Indian J. Med. Res. 2013, 137, 767–776. [Google Scholar] [PubMed]
- Guerra, J.A.; Prestes, S.R.; Silveira, H.; Coelho, L.I.; Gama, P.; Moura, A.; Amato, V.; Barbosa, M.; Ferreira, L.C. Mucosal leishmaniasis caused by Leishmania (Viannia) braziliensis and Leishmania (Viannia) guyanensis in the Brazilian Amazon. PLoS Negl. Trop. Dis. 2011, 5. [Google Scholar] [CrossRef] [PubMed]
- Paniz Mondolfi, A.E.; Stavropoulos, C.; Gelanew, T.; Loucas, E.; Perez Alvarez, A.M.; Benaim, G.; Polsky, B.; Schoenian, G.; Sordillo, E.M. Successful Treatment of Old World Cutaneous Leishmaniasis Caused by Leishmania infantum with Posaconazole. Antimicrob. Agents Chemother. 2011, 55, 1774–1776. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Control of the Leishmaniasis; WHO: Geneva, Switzerland, 2010. [Google Scholar]
- García-Hernández, R.; Manzano, J.I.; Castanys, S.; Gamarro, F. Leishmania donovani develops resistance to drug combinations. PLOS Negl. Trop. Dis. 2012, 6. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Kumar, M.; Singh, R.K. Leishmaniasis: Current status of available drugs and new potential drug targets. Asian Pac. J. Trop. Med. 2012, 5, 485–497. [Google Scholar] [CrossRef]
- Olliaro, P.L. Drug combinations for visceral leishmaniasis. Curr. Opin. Infect. Dis. 2010, 23. [Google Scholar] [CrossRef] [PubMed]
- Kobets, T.; Grekov, I.; Lipoldova, M. Leishmaniasis: Prevention, parasite detection and treatment. Curr. Med. Chem. 2012, 19, 1443–1474. [Google Scholar] [CrossRef]
- Alcântara, L.M.; Ferreira, T.C.S.; Gadelha, F.R.; Miguel, D.C. Challenges in drug discovery targeting TriTryp diseases with an emphasis on leishmaniasis. Int. J. Parasitol. Drugs Drug Resist. 2018, 8, 430–439. [Google Scholar] [CrossRef]
- DNDi. Target Product Profile—Cutaneous Leishmaniasis. Available online: https://www.dndi.org/diseases-projects/leishmaniasis/tpp-cl/ (accessed on 16 October 2018).
- DNDi Target Product Profile—Visceral Leishmaniasis. Available online: https://www.dndi.org/diseases-projects/leishmaniasis/tpp-vl/ (accessed on 16 October 2018).
- Marín, C.; Clares, M.P.; Ramírez-Macías, I.; Blasco, S.; Olmo, F.; Soriano, C.; Verdejo, B.; Rosales, M.J.; Gomez-Herrera, D.; García-España, E.; et al. In vitro activity of scorpiand-like azamacrocycle derivatives in promastigotes and intracellular amastigotes of Leishmania infantum and Leishmania braziliensis. Eur. J. Med. Chem. 2013, 62, 466–477. [Google Scholar] [CrossRef]
- Martín-Montes, A.; Santivañez-Veliz, M.; Moreno-Viguri, E.; Martín-Escolano, R.; Jiménez-Montes, C.; Lopez-Gonzalez, C.; Marín, C.; Sanmartín, C.; Gutiérrez Sánchez, R.; Sánchez-Moreno, M.; et al. In vitro antileishmanial activity and iron superoxide dismutase inhibition of arylamine Mannich base derivatives. Parasitology 2017, 144, 1783–1790. [Google Scholar] [CrossRef]
- Martín-Escolano, R.; Moreno-Viguri, E.; Santivañez-Veliz, M.; Martin-Montes, A.; Medina-Carmona, E.; Paucar, R.; Marín, C.; Azqueta, A.; Cirauqui, N.; Pey, A.L.; et al. Second Generation of Mannich Base-Type Derivatives with in Vivo Activity against Trypanosoma cruzi. J. Med. Chem. 2018, 61, 5643–5663. [Google Scholar] [CrossRef] [PubMed]
- Mehlotra, R.K. Antioxidant defense mechanisms in parasitic protozoa. Crit. Rev. Microbiol. 1996, 22, 295–314. [Google Scholar] [CrossRef] [PubMed]
- González-Bártulos, M.; Aceves-Luquero, C.; Qualai, J.; Cussó, O.; Martínez, M.A.; Fernández de Mattos, S.; Menéndez, J.A.; Villalonga, P.; Costas, M.; Ribas, X.; et al. Pro-Oxidant Activity of Amine-Pyridine-Based Iron Complexes Efficiently Kills Cancer and Cancer Stem-Like Cells. PLoS ONE 2015, 10. [Google Scholar] [CrossRef] [PubMed]
- Nwaka, S.; Hudson, A. Innovative lead discovery strategies for tropical diseases. Nat. Rev. 2006, 5, 941–955. [Google Scholar] [CrossRef] [PubMed]
- Ginger, M.L. Trypanosomatid biology and euglenozoan evolution: New insights and shifting paradigms revealed through genome sequencing. Protist 2005, 156, 337–392. [Google Scholar] [CrossRef] [PubMed]
- Cazzulo, J.J. Aerobic fermentation of glucose by trypanosomatids. FASEB J. 1992, 6, 3153–3161. [Google Scholar] [CrossRef] [PubMed]
- Cussó, O.; Garcia-Bosch, I.; Ribas, X.; Lloret-Fillol, J.; Costas, M. Asymmetric epoxidation with H2O2 by manipulating the electronic properties of non-heme iron catalysts. J. Am. Chem. Soc. 2013, 135, 14871–14878. [Google Scholar] [CrossRef]
- Costas, M.; Que, L., Jr. Ligand topology tuning of iron-catalyzed hydrocarbon oxidations. Angew. Chem. Int. Ed. 2002, 41, 2179–21781. [Google Scholar] [CrossRef]
- Bringaud, F.; Rivière, L.; Coustou, V. Energy metabolism of trypanosomatids: Adaptation to available carbon sources. Mol. Biochem. Parasitol. 2006, 149, 1–9. [Google Scholar] [CrossRef]
- Saunders, E.C.; de Souza, D.P.; Naderer, T.; Sernee, M.F.; Ralton, J.E.; Doyle, M.A.; MacRae, J.I.; Chambers, J.L.; Heng, J.; Nahid, A.; et al. Central carbon metabolism of Leishmania parasites. Parasitol. 2010, 137, 1303–1313. [Google Scholar] [CrossRef]
- Depledge, D.P.; MacLean, L.M.; Hodgkinson, M.R.; Smith, B.A.; Jackson, A.P.; Ma, S.; Uliana, S.R.; Smith, D.F. Leishmania-specific surface antigens show sub-genus sequence variation and immune recognition. PLoS Negl. Trop. Dis. 2010, 4. [Google Scholar] [CrossRef] [PubMed]
- Kirkinezos, I.G.; Moraes, C.T. Reactive oxygen species and mitochondrial diseases. Semin. Cell Dev. Biol. 2001, 12, 449–457. [Google Scholar] [CrossRef] [PubMed]
- Michels, P.A.; Bringaud, F.; Herman, M.; Hannaert, V. Metabolic functions of glycosomes in trypanosomatids. Biochim. Biophys. Acta. 2006, 1763, 1463–1477. [Google Scholar] [CrossRef] [PubMed]
- Company, A.; Gómez, L.; Fontrodona, X.; Ribas, X.; Costas, M. A Novel Platform for Modeling Oxidative Catalysis in Non-Heme Iron Oxygenases with Unprecedented Efficiency. Chem. Eur. J. 2008, 14, 5727–5731. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Bosch, I.; Company, A.; Fontrodona, X.; Ribas, X.; Costas, M. Efficient and Selective Peracetic Acid Epoxidation Catalyzed by a Robust Manganese Catalyst. Org. Lett. 2008, 10, 2095–2098. [Google Scholar] [CrossRef] [PubMed]
- Glerup, J.; Goodson, P.A.; Hazell, A.; Hazell, R.; Hodgson, D.J.; McKenzie, C.J.; Michelsen, K.; Rychlewska, U.; Toftlund, H. Synthesis and Characterization of Bis(.mu.-oxo)dimanganese(III,III), -(III,IV), and -(IV,IV) Complexes with Ligands Related to N,N′-Bis(2-pyridylmethyl)-1,2-ethanediamine (Bispicen). Inorg. Chem. 1994, 33, 4105–4111. [Google Scholar] [CrossRef]
- Christiansen, L.; Hendrickson, D.N.; Toftlund, H.; Wilson, S.R.; Xie, C.L. Synthesis and structure of metal complexes of triaza macrocycles with three pendant pyridylmethyl arms. Inorg. Chem. 1986, 25, 2813–2818. [Google Scholar] [CrossRef]
- Wieghardt, K.; Schoeffmann, E.; Nuber, B.; Weiss, J. Syntheses, properties and electrochemistry of transition-metal complexes of the macrocycle 1,4,7-tris(2-pyridylmethyl)-1,4,7-triazacyclononane (L). Crystal structures of [NiL](ClO4)2, [MnL](ClO4)2, and [PdL](PF6)2 containing a distorted-square-base-pyramidal PdIIN5 core. Inorg. Chem. 1986, 25, 4877–4883. [Google Scholar]
- González, P.; Marín, C.; Rodríguez-González, I.; Hitos, A.B.; Rosales, M.J.; Reina, M.; Díaz, J.G.; González-Coloma, A.; Sánchez-Moreno, M. In vitro activity of C20-diterpenoid alkaloid derivatives in promastigotes and intracellular amastigotes of Leishmania infantum. Int. J. Antimicrob. Agents 2005, 25, 136–141. [Google Scholar] [CrossRef]
- Marín, C.; Ramírez-Macías, I.; López-Céspedes, A.; Olmo, F.; Villegas, N.; Díaz, J.G.; Rosales, M.J.; Gutiérrez-Sánchez, R.; Sánchez-Moreno, M. In vitro and in vivo trypanocidal activity of flavonoids from Delphinium staphisagria against Chagas disease. J. Nat. Prod. 2011, 74, 744–750. [Google Scholar] [CrossRef]
- Sánchez-Moreno, M.; Gómez-Contreras, F.; Navarro, P.; Marín, C.; Ramírez-Macías, I.; Olmo, F.; Sanz, A.M.; Campayo, L.; Cano, C.; Yunta, M.J.R. In vitro leishmanicidal activity of imidazole- or pyrazole-based benzo[g]phthalazine derivatives against Leishmania infantum and Leishmania braziliensis species. J. Antimicrob. Chemother. 2012, 67, 387–397. [Google Scholar] [CrossRef] [PubMed]
- Bourguignon, S.C.; Mello, C.B.; Santos, D.O.; Gonzalez, M.S.; Souto-Padron, T. Biological aspects of the Trypanosoma cruzi (Dm28c clone) intermediate form, between epimastigote and trypomastigote, obtained in modified liver infusion tryptose (LIT) medium. Act. Trop. 2006, 98, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Becerra, C.; Sánchez-Moreno, M.; Osuna, A.; Opperdoes, F.R. Comparative aspects of energy metabolism in plant trypanosomatids. J. Eukar. Microbiol. 1997, 44, 523–529. [Google Scholar] [CrossRef]
- López-Céspedes, A.; Longoni, S.S.; Sauri-Arceo, C.H.; Sánchez-Moreno, M.; Rodríguez-Vivas, R.I.; Escobedo-Ortegón, F.J.; Barrera-Pérez, M.A.; Bolio-González, M.E.; Marín, C. Leishmania spp. epidemiology of canine leishmaniasis in the Yucatan Peninsula. ScientificWorldJournal 2012, 2012. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Beyer, W.F.; Fridovich, I. Assaying for superoxide dismutase activity: Some large consequences of minor changes in conditions. Anal. Biochem. 1987, 161, 559–566. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are available from the authors. |



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | IC50 µM a | Toxicity IC50 Macrophage (µM) | |||||
|---|---|---|---|---|---|---|---|
| Leishmania infantum | Leishmania braziliensis | Leishmania donovani | |||||
| Promastigote Forms | Amastigote Forms | Promastigote Forms | Amastigote Forms | Promastigote Forms | Amastigote Forms | ||
| Glucantime® | 18.0 ± 3.1 | 24.2 ± 2.6 | 25.6 ± 1.6 | 30.4 ± 6.1 | 27.3 ± 4.3 | 33.3 ± 3.7 | 15.20 ± 1.3 |
| 1 | 18.5 ± 1.6 | 20.5 ± 1.8 | 22.3 ± 0.6 | 21.3 ± 0.9 | 28.6 ± 5.8 | 12.5 ± 2.7 | 37.9 ± 4.2 |
| 2 | 52.6 ± 3.1 | 57.5 ± 2.6 | 37.2 ± 1.9 | 12.5 ± 1.5 | 19.7 ± 1.7 | 16.3 ± 2.5 | 105.6 ± 8.8 |
| 3 | 8.5 ± 0.4 | 3.7 ± 1.6 | 9.3 ± 0.3 | 8.7 ± 0.6 | 28.6 ± 3.5 | 11.7 ± 1.3 | 98.5 ± 6.3 |
| 4 | 27.4 ± 0.7 | 24.7 ± 1.2 | 26.4 ± 0.7 | 19.4 ± 1.7 | 60.2 ± 7.1 | 29.9 ± 1.6 | 60.3 ± 3.3 |
| 5 | 1.3 ± 0.1 | 2.3 ± 0.8 | 3.5 ± 0.6 | 8.8 ± 1.3 | 6.8 ± 0.2 | 10.7 ± 0.8 | 97.2 ± 3.8 |
| 6 | 7.3 ± 0.1 | 10.2 ± 1.5 | 10.1 ± 3.1 | 11.7 ± 1.1 | 4.0 ± 0.1 | 1.3 ± 0.4 | 137.6 ± 7.4 |
| 7 | 12.3 ± 2.2 | 9.0 ± 0.4 | 11.7 ± 0.8 | 8.5 ± 0.3 | 8.1 ± 3.9 | 1.1 ± 0.2 | 172.1 ± 11.3 |
| 8 | 83.0 ± 9.9 | 22.5 ± 3.1 | 73.8 ± 3.6 | 21.7 ± 1.4 | 31.7 ± 2.4 | 1.9 ± 0.3 | 268.3 ± 16.2 |
| 9 | 71.7 ± 10.0 | 33.4 ± 3.2 | 53.2 ± 3.6 | 30.7 ± 2.5 | 29.5 ± 3.6 | 14.5 ± 2.7 | 100.5 ± 4.8 |
| 10 | 2.6 ± 0.7 | 10.4 ± 0.1 | 5.5 ± 0.8 | 11.9 ± 0.8 | 18.6 ± 0.9 | 11.6 ± 3.3 | 126.1 ± 7.1 |
| 11 | 87.5 ± 5.1 | 34.6 ± 2.5 | 60.6 ± 3.8 | 30.4 ± 1.3 | 29.5 ± 4.1 | 19.8 ± 2.6 | 75.1 ± 4.7 |
| SI b | |||||||
| Leishmania infantum | Leishmania braziliensis | Leishmania donovani | |||||
| Promastigote Forms | Amastigote Forms | Promastigote Forms | Amastigote Forms | Promastigote Forms | Amastigote Forms | ||
| Glucantime® | 0.8 | 0.6 | 0.6 | 0.5 | 0.6 | 0.5 | |
| 1 | 2.1 (3) | 1.8 (3) | 1.7 (3) | 1.8 (4) | 1.3 (2) | 3.0 (6) | |
| 2 | 2.0 (2) | 1.8 (3) | 2.8 (5) | 8.4 (17) | 5.4 (9) | 6.5 (13) | |
| 3 | 11.6 (14) | 26.6 (44) | 10.6 (18) | 11.3 (23) | 3.4 (6) | 8.4 (17) | |
| 4 | 2.2 (3) | 2.4 (4) | 2.3 (4) | 3.1 (6) | 1.0 (2) | 2.0 (4) | |
| 5 | 74.8 (93) | 42.3 (70) | 27.8 (46) | 11.0 (22) | 14.3 (24) | 9.1 (18) | |
| 6 | 18.8 (24) | 13.5 (22) | 13.6 (23) | 11.8 (24) | 34.4 (57) | 105.8 (212) | |
| 7 | 14.0 (17) | 19.1 (32) | 14.7 (25) | 20.2 (40) | 21.2 (35) | 156.5 (313) | |
| 8 | 3.2 (4) | 11.9 (20) | 3.6 (6) | 12.4 (25) | 8.5 (14) | 141.2 (282) | |
| 9 | 1.4 (2) | 3.0 (5) | 1.9 (3) | 3.3 (7) | 3.4 (6) | 6.9 (14) | |
| 10 | 48.5 (61) | 12.1 (20) | 22.9 (38) | 10.6 (21) | 6.8 (11) | 10.9 (22) | |
| 11 | 0.9 (1) | 2.2 (4) | 1.2 (2) | (5) | 2.5 (4) | 3.8 (8) | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Urbanová, K.; Ramírez-Macías, I.; Martín-Escolano, R.; Rosales, M.J.; Cussó, O.; Serrano, J.; Company, A.; Sánchez-Moreno, M.; Costas, M.; Ribas, X.; et al. Effective Tetradentate Compound Complexes against Leishmania spp. that Act on Critical Enzymatic Pathways of These Parasites. Molecules 2019, 24, 134. https://doi.org/10.3390/molecules24010134
Urbanová K, Ramírez-Macías I, Martín-Escolano R, Rosales MJ, Cussó O, Serrano J, Company A, Sánchez-Moreno M, Costas M, Ribas X, et al. Effective Tetradentate Compound Complexes against Leishmania spp. that Act on Critical Enzymatic Pathways of These Parasites. Molecules. 2019; 24(1):134. https://doi.org/10.3390/molecules24010134
Chicago/Turabian StyleUrbanová, Kristína, Inmaculada Ramírez-Macías, Rubén Martín-Escolano, María José Rosales, Olaf Cussó, Joan Serrano, Anna Company, Manuel Sánchez-Moreno, Miquel Costas, Xavi Ribas, and et al. 2019. "Effective Tetradentate Compound Complexes against Leishmania spp. that Act on Critical Enzymatic Pathways of These Parasites" Molecules 24, no. 1: 134. https://doi.org/10.3390/molecules24010134
APA StyleUrbanová, K., Ramírez-Macías, I., Martín-Escolano, R., Rosales, M. J., Cussó, O., Serrano, J., Company, A., Sánchez-Moreno, M., Costas, M., Ribas, X., & Marín, C. (2019). Effective Tetradentate Compound Complexes against Leishmania spp. that Act on Critical Enzymatic Pathways of These Parasites. Molecules, 24(1), 134. https://doi.org/10.3390/molecules24010134