Betaine Attenuates Monocrotaline-Induced Pulmonary Arterial Hypertension in Rats via Inhibiting Inflammatory Response

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Experimental Design
2.2. Assessment of Hemodynamics
2.3. Measurement of RVHI
2.4. H&E and Masson Staining
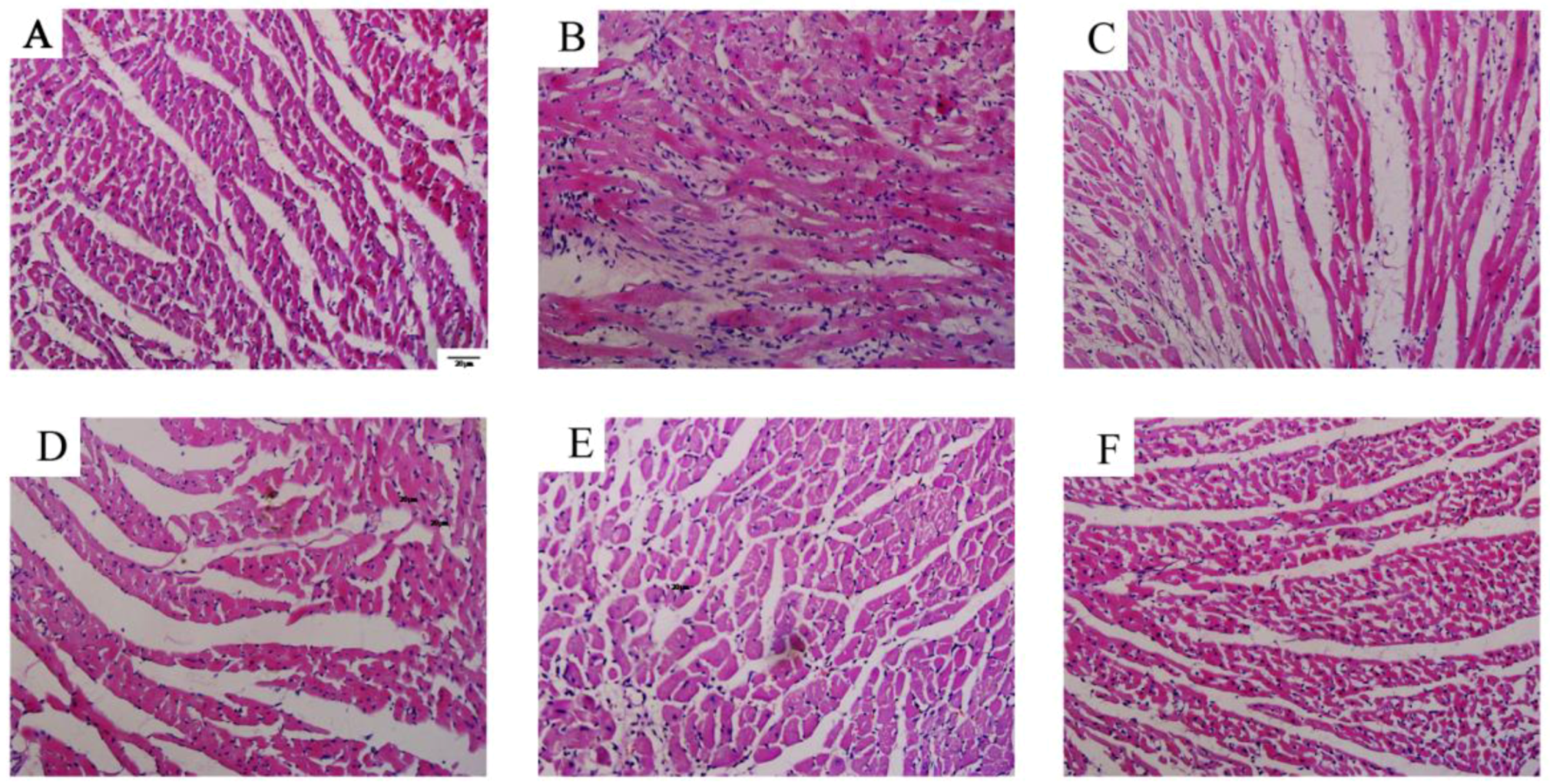
2.5. Evaluation of RV Remodeling
2.6. Immunohistologic Study
2.7. Western Blot
2.8. Data Analysis
3. Results
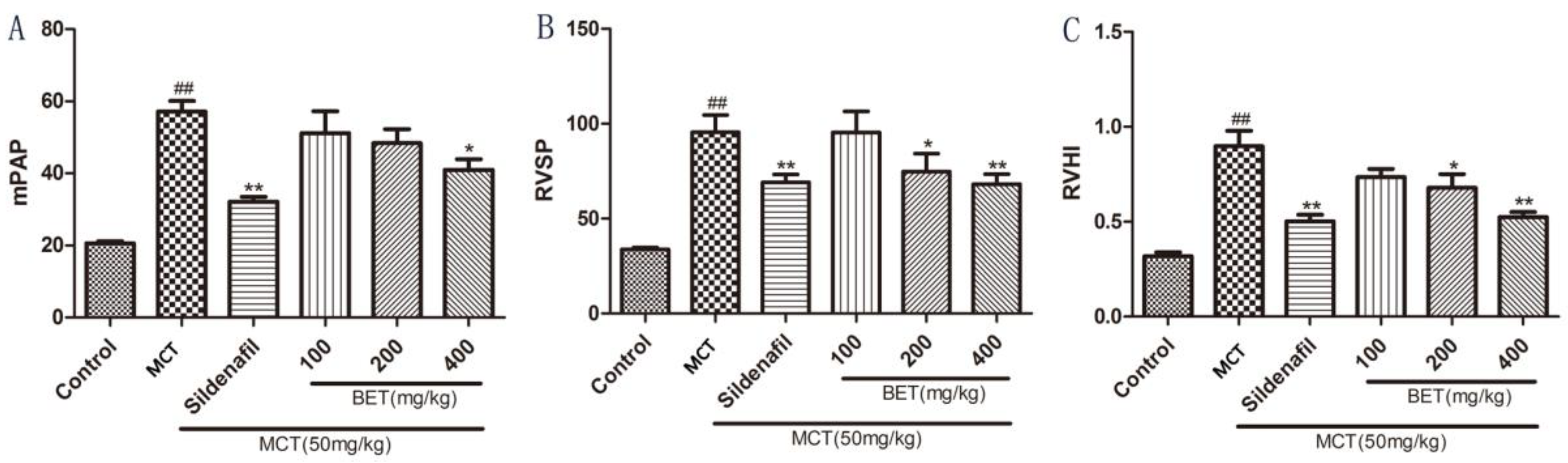
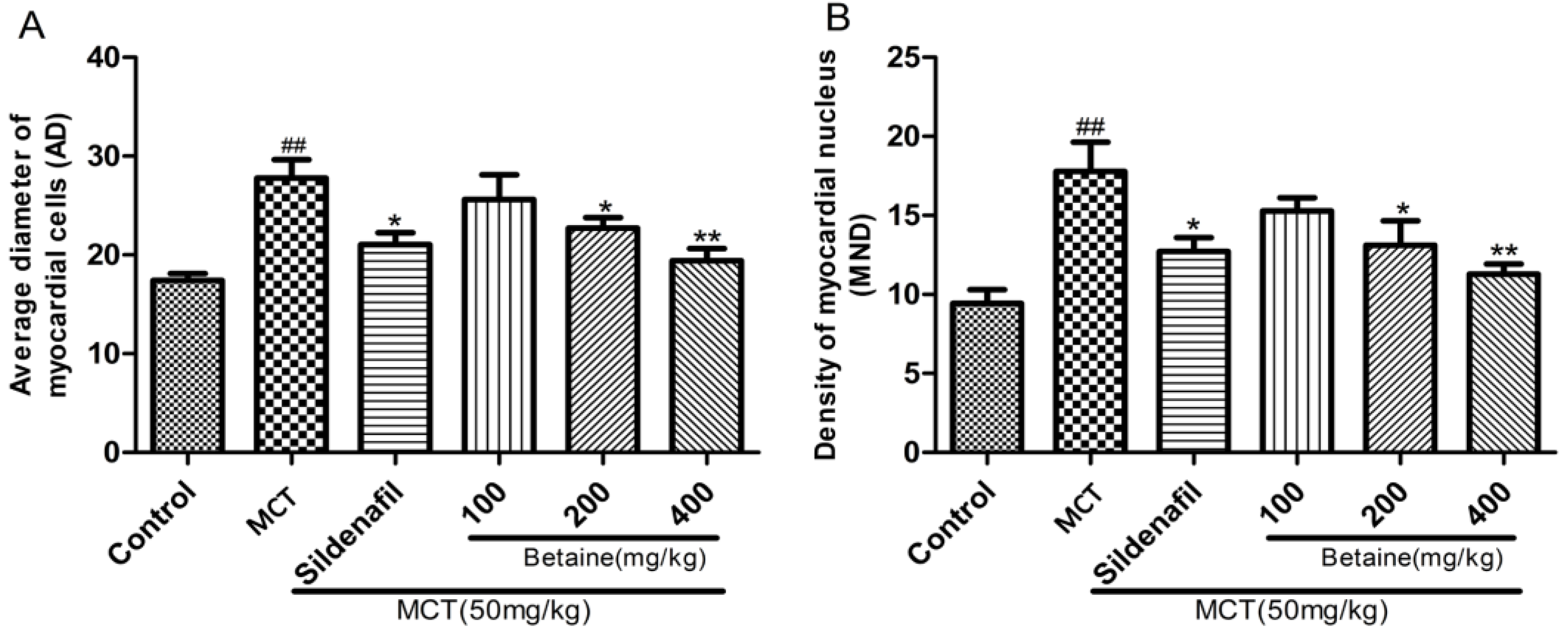
3.1. Betaine Inhibits the Development of PAH and RV Hypertrophy
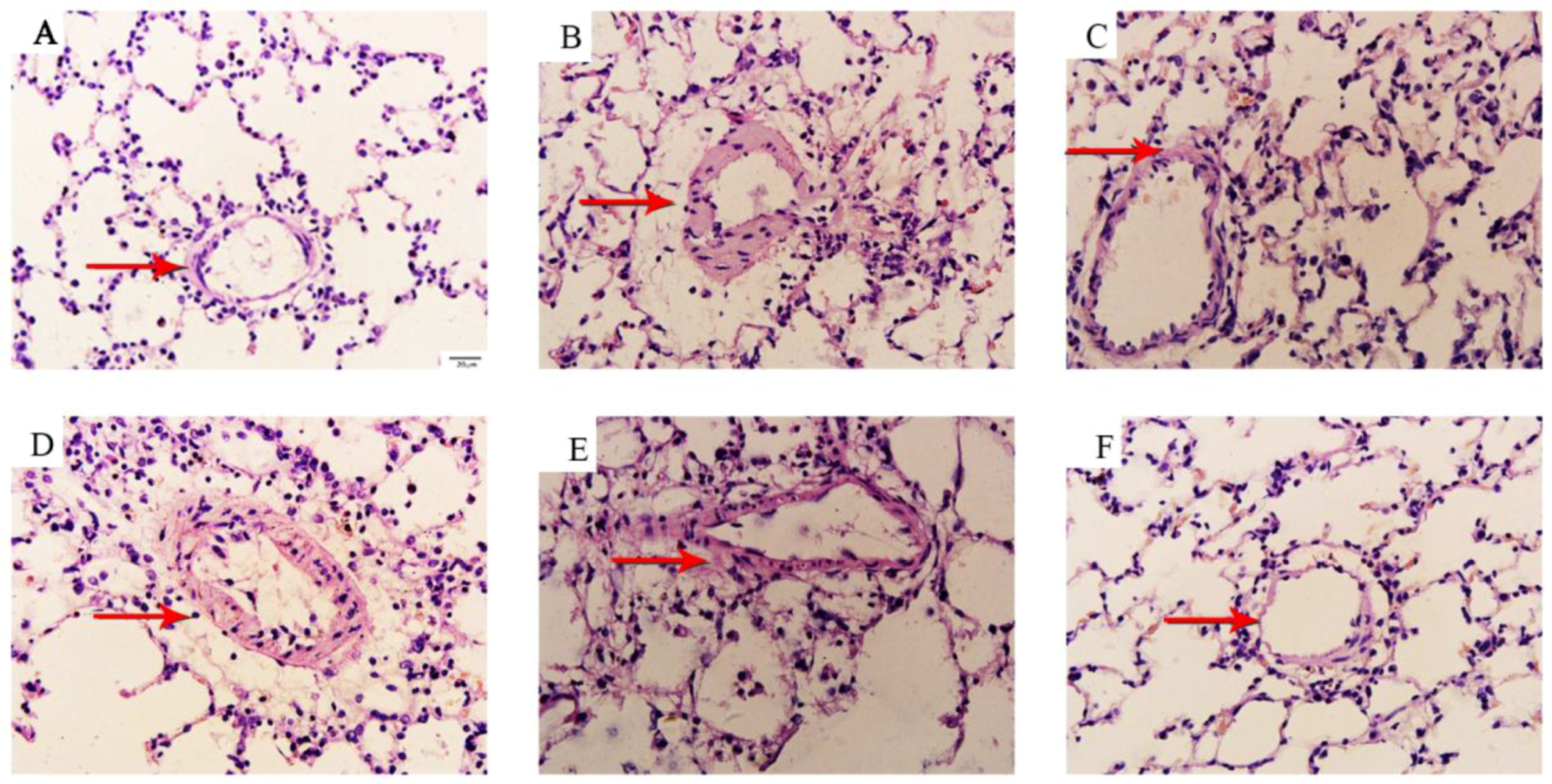
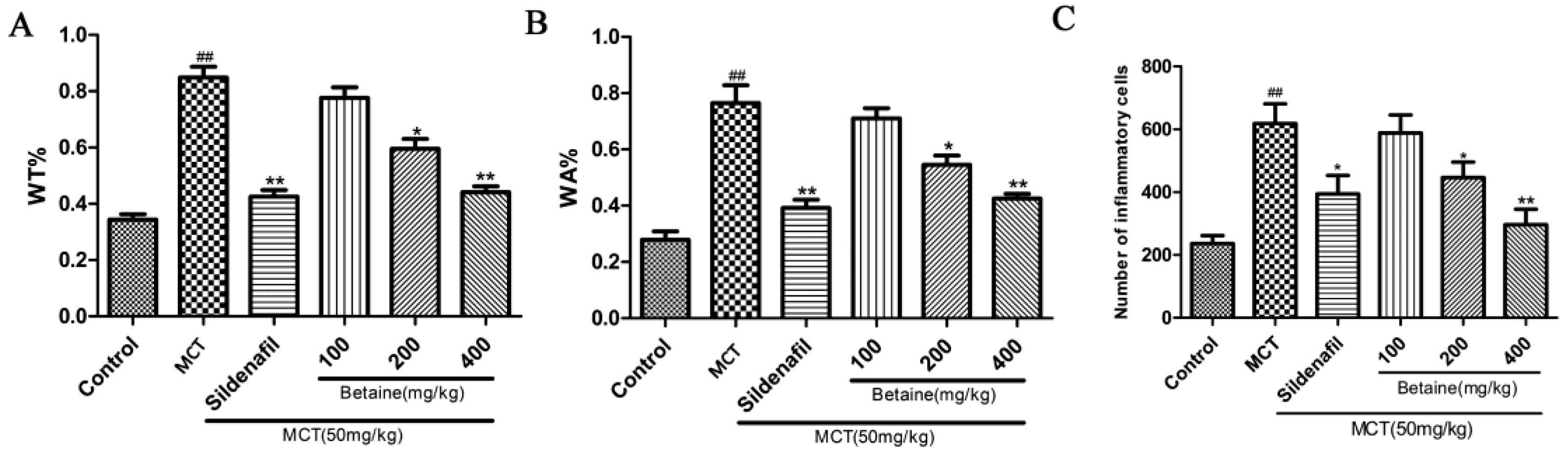
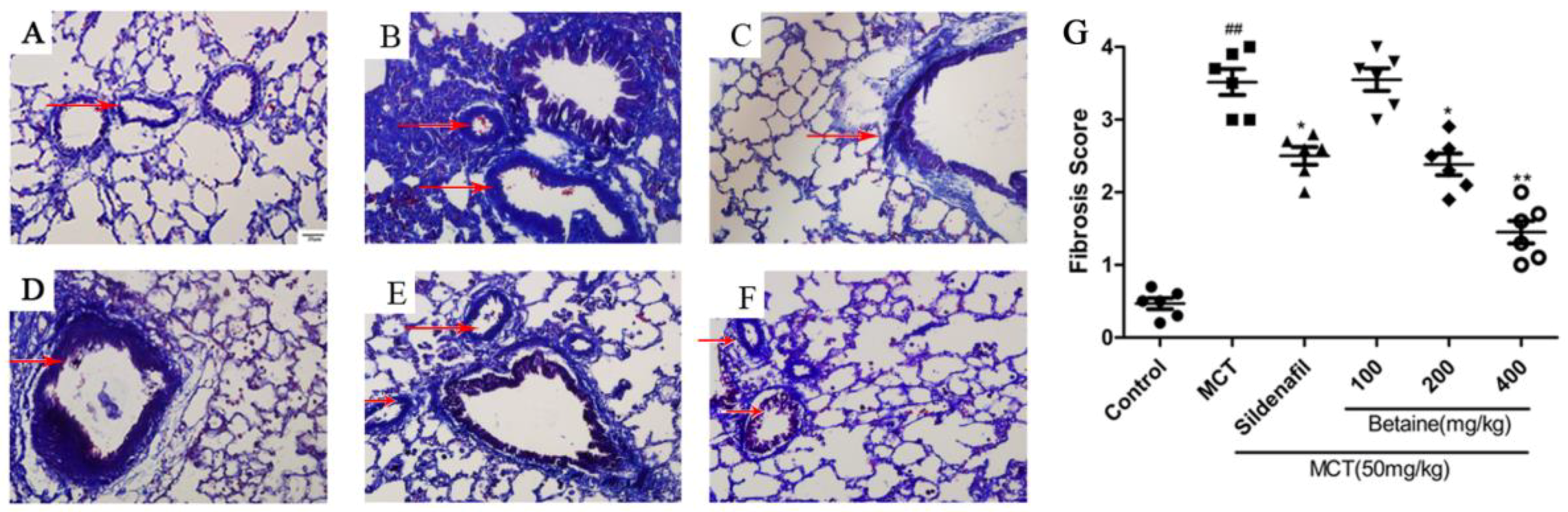
3.2. Betaine Reverses Pulmonary Vascular and RV Remodeling and Inhibits Inflammatory Cells Infiltration
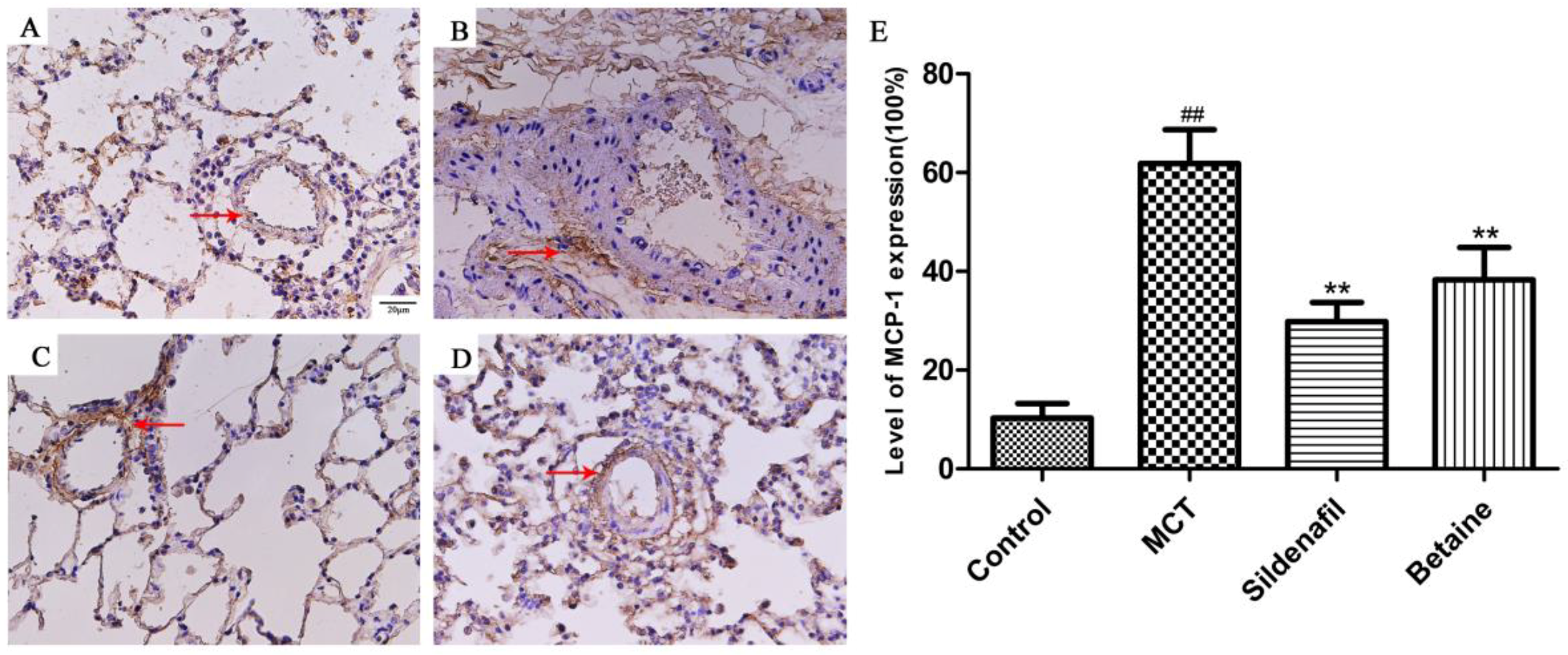
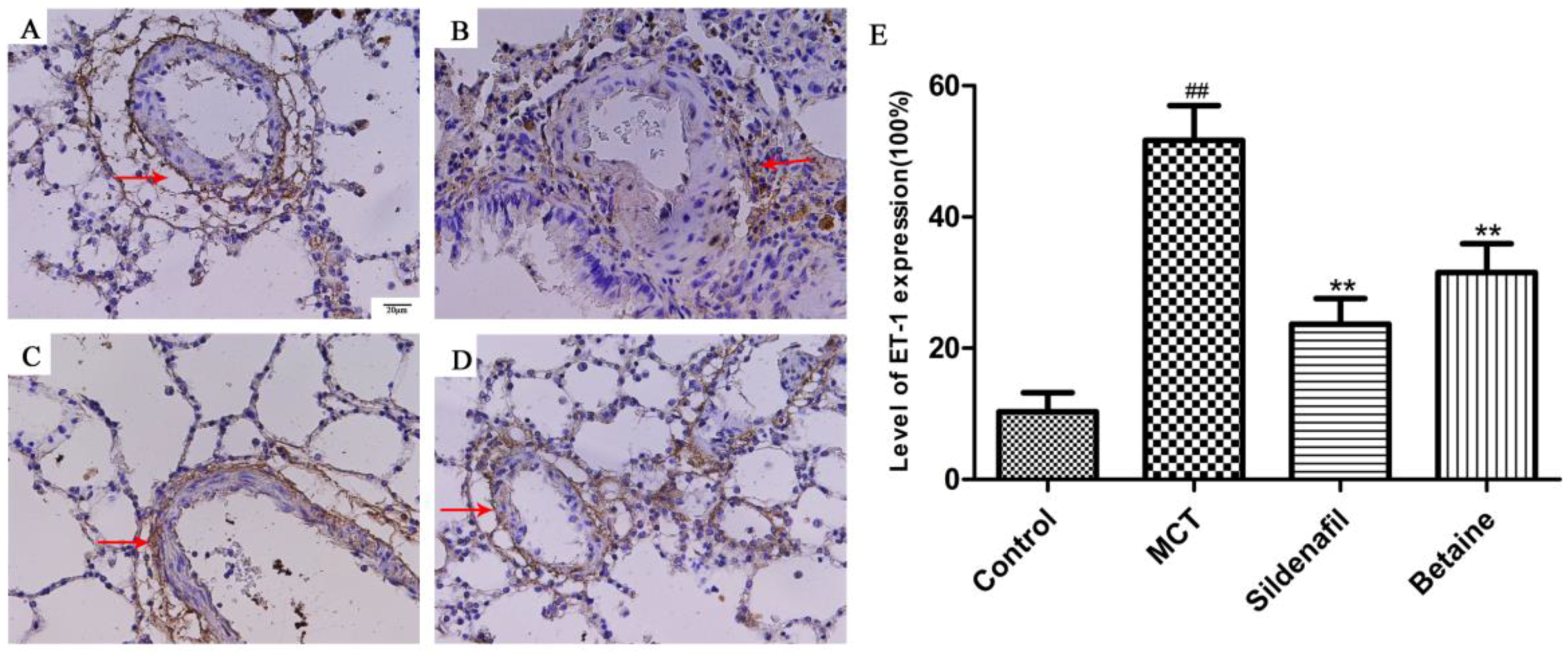
3.3. Immunohistochemical Analyses Revealed Betaine Attenuated MCP-1, ET-1 Expression in Lung
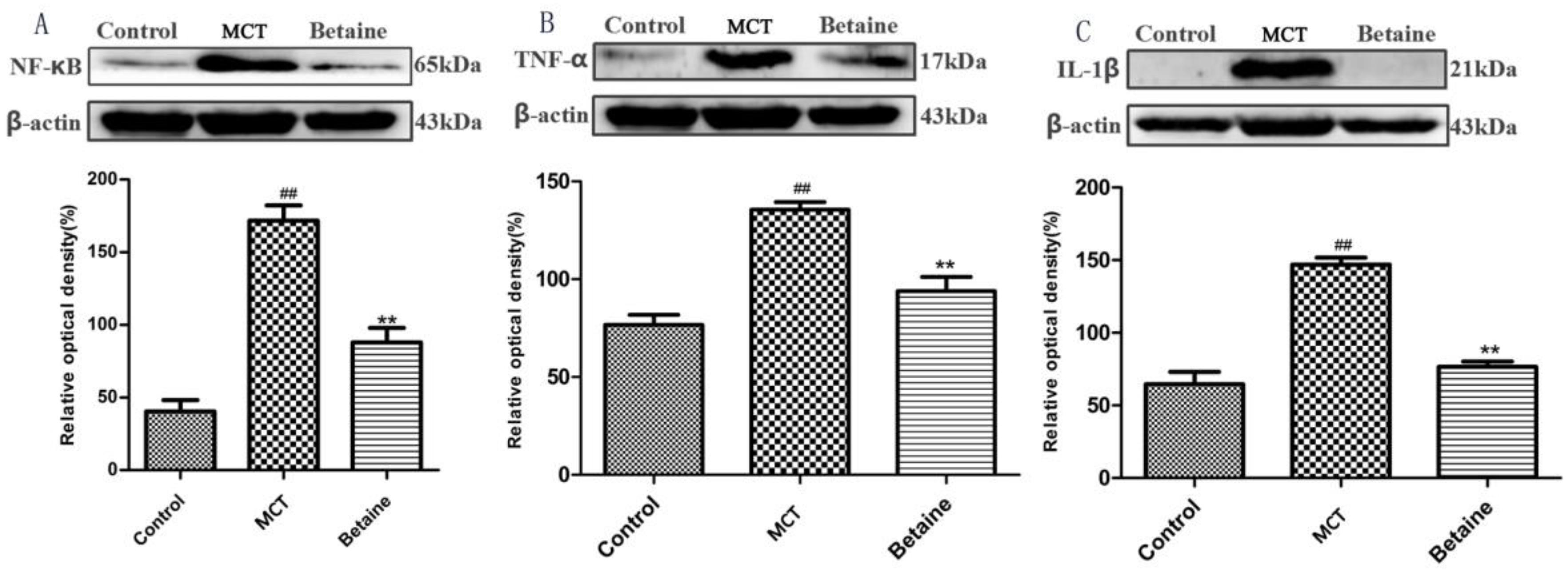
3.4. Betaine Attenuates NF-κB, TNF-α, and IL-1β Protein Expression
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Yung, L.M.; Nikolic, I.; Paskin-Flerlage, S.D.; Pearsall, R.S.; Kumar, R.; Yu, P.B. A Selective Transforming Growth Factor-β Ligand Trap Attenuates Pulmonary Hypertension. Am. J. Respir. Crit. Care Med. 2016, 194, 1140–1151. [Google Scholar] [CrossRef] [PubMed]
- Rhodes, C.J.; Ghataorhe, P.; Wharton, J.; Rue-Albrecht, K.C.; Hadinnapola, C.; Watson, G.; Bleda, M.; Haimel, M.; Coghlan, G.; Corris, P.A. Plasma Metabolomics Implicates Modified Transfer RNAs and Altered Bioenergetics in the Outcomes of Pulmonary Arterial Hypertension. Circulation 2017, 135, 460–475. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, Y.J.; Ibrahim, Y.F.; Shults, N.V. Apoptosis-based therapy to treat pulmonary arterial hypertension. J. Rare Dis. Res. Treat. 2016, 1, 17–24. [Google Scholar] [PubMed]
- Wang, Q.; Zuo, X.R.; Wang, Y.Y.; Xie, W.P.; Wang, H.; Zhang, M. Monocrotaline-induced pulmonary arterial hypertension is attenuated by TNF-alpha antagonists via the suppression of TNF-alpha expression and NF-κB pathway in rats. Vascul. Pharmacol. 2013, 58, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Wong, M.J.; Kantores, C.; Ivanovska, J.; Jain, A.; Jankov, R.P. Simvastatin prevents and reverses chronic pulmonary hypertension in newborn rats via pleiotropic inhibition of RhoA signaling. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2016, 311, L985–L999. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Liang, X.; Zhou, Y.; Zhang, S.; Yang, F.; Guo, H.; Fan, R.; Feng, N.; Jia, M.; Wang, Y.; et al. Role of dynorphin in hypoxic pulmonary hypertension. Eur. J. Pharmacol. 2016, 791, 78–84. [Google Scholar] [CrossRef] [PubMed]
- Huertas, A.; Tu, L.; Thuillet, R.; Le Hiress, M.; Phan, C.; Ricard, N.; Nadaud, S.; Fadel, E.; Humbert, M.; Guignabert, C. Leptin signalling system as a target for pulmonary arterial hypertension therapy. Eur. Respir. J. 2015, 45, 1066–1080. [Google Scholar] [CrossRef] [PubMed]
- Farkas, D.; Alhussaini, A.A.; Kraskauskas, D.; Kraskauskiene, V.; Cool, C.D.; Nicolls, M.R.; Natarajan, R.; Farkas, L. Nuclear factor κB inhibition reduces lung vascular lumen obliteration in severe pulmonary hypertension in rats. Am. J. Respir. Cell Mol. Biol. 2014, 51, 413–425. [Google Scholar] [CrossRef] [PubMed]
- Price, L.C.; Caramori, G.; Perros, F.; Meng, C.; Gambaryan, N.; Dorfmuller, P.; Montani, D.; Casolari, P.; Zhu, J.; Dimopoulos, K. Nuclear factor κ-B is activated in the pulmonary vessels of patients with end-stage idiopathic pulmonary arterial hypertension. PLoS ONE 2013, 8, e75415. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Wei, C.; Kim, I.K.; Janssen-Heininger, Y.; Gupta, S. Inhibition of nuclear factor-κB in the lungs prevents monocrotaline-induced pulmonary hypertension in mice. Hypertension 2014, 63, 1260–1269. [Google Scholar] [CrossRef] [PubMed]
- Voelkel, N.F.; Tuder, R.M.; Bridges, J.; Arend, W.P. Interleukin-1 receptor antagonist treatment reduces pulmonary hypertension generated in rats by monocrotaline. Am. J. Respir. Cell Mol. Biol. 1994, 11, 664–675. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.Q.; Xiao, J.; Fan, H.X.; Yu, Y.; He, R.R.; Feng, X.L.; Kurihara, H.; So, K.F.; Yao, X.S.; Gao, H. Polyphenols from wolfberry and their bioactivities. Food Chem. 2017, 214, 644–654. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.H.; Sung, B.; Kang, Y.J.; Jang, J.Y.; Hwang, S.Y.; Lee, Y.; Kim, M.; Im, E.; Yoon, J.H.; Kim, C.M.; et al. Anti-inflammatory effects of betaine on AOM/DSSinduced colon tumorigenesis in ICR male mice. Int. J. Oncol. 2014, 45, 1250–1256. [Google Scholar] [CrossRef] [PubMed]
- Bingül, İ.; Başaran-Küçükgergin, C.; Aydın, A.F.; Çoban, J.; Doğan-Ekici, I.; Doğru-Abbasoğlu, S.; Uysal, M. Betaine treatment decreased oxidative stress, inflammation, and stellate cell activation in rats with alcoholic liver fibrosis. Environ. Toxicol. Pharmacol. 2016, 45, 170–178. [Google Scholar] [CrossRef] [PubMed]
- Ndisang, J.F. The heme oxygenase system selectively modulates proteins implicated in metabolism, oxidative stress and inflammation in spontaneously hypertensive rats. Curr. Pharm. Des. 2014, 20, 1318–1327. [Google Scholar] [CrossRef] [PubMed]
- Miwa, M.; Tsuboi, M.; Noguchi, Y.; Enokishima, A.; Nabeshima, T.; Hiramatsu, M. Effects of betaine on lipopolysaccharide-induced memory impairment in mice and the involvement of GABA transporter 2. J. Neuroinflamm. 2011, 8, 153. [Google Scholar]
- Bingül, İ.; Aydın, A.F.; Başaran-Küçükgergin, C.; Doğan-Ekici, I.; Çoban, J.; Doğru-Abbasoğlu Sv Uysal, M. High-fat diet plus carbon tetrachloride-induced liver fibrosis is alleviated by betaine treatment in rats. Int. Immunopharmacol. 2016, 39, 199–207. [Google Scholar]
- Zheng, P.; Liu, J.; Mai, S.; Yuan, Y.; Wang, Y.; Dai, G. Regulation of signal transducer and activator of transcription 3 and apoptotic pathways by betaine attenuates isoproterenol-induced acute myocardial injury in rats. Hum. Exp. Toxicol. 2015, 34, 538–547. [Google Scholar] [CrossRef] [PubMed]
- Yi, E.Y.; Kim, Y.J. Betaine inhibits in vitro and in vivo angiogenesis through suppression of the NF-κB and Akt signaling pathways. Int. J. Oncol. 2012, 41, 1879–1885. [Google Scholar] [CrossRef] [PubMed]
- Go, E.K.; Jung, K.J.; Kim, J.Y.; Yu, B.P.; Chung, H.Y. Betaine suppresses proinflammatory signaling during aging: The involvement of nuclear factor-κB via nuclear factor-inducing kinase/IκB kinase and mitogen-activated protein kinases. J. Gerontol. A Biol. Sci. Med. Sci. 2005, 60, 1252–1264. [Google Scholar] [CrossRef] [PubMed]
- Xiao, T.; Xie, L.; Huang, M.; Shen, J. Differential expression of microRNA in the lungs of rats with pulmonary arterial hypertension. Mol. Med. Rep. 2017, 15, 591–596. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Yin, J.; Cai, W.; Liu, J.; Zhang, N.; Yan, S.; Song, L.; Li, X. Coupling Factor 6 Is Upregulated in Monocrotaline-induced Pulmonary Arterial Hypertension in Rats. Am. J. Med. Sci. 2016, 352, 631–636. [Google Scholar] [CrossRef] [PubMed]
- Varshney, R. Monocrotaline-Induced Pulmonary Hypertension Involves Downregulation of Antiaging Protein Klotho and eNOS ActivityNovelty and Significance. Hypertension 2016, 68, 1255–1263. [Google Scholar] [CrossRef] [PubMed]
- Shields, K.J.; Verdelis, K.; Passineau, M.J.; Faight, E.M.; Zourelias, L.; Wu, C.; Chong, R.; Benza, R.L. Three-dimensional micro computed tomography analysis of the lung vasculature and differential adipose proteomics in the Sugen/hypoxia rat model of pulmonary arterial hypertension. Pulm. Circ. 2016, 6, 586–596. [Google Scholar] [CrossRef] [PubMed]
- Li, X.Q.; Wang, H.M.; Yang, C.G.; Zhang, X.H.; Han, D.D.; Wang, H.L. Fluoxetine inhibited extracellular matrix of pulmonary artery and inflammation of lungs in monocrotaline-treated rats. Acta Pharmacol. Sin. 2011, 32, 217–222. [Google Scholar] [CrossRef] [PubMed]
- Alirezaei, M.; Khoshdel, Z.; Dezfoulian, O.; Rashidipour, M.; Taghadosi, V. Beneficial antioxidant properties of betaine against oxidative stress mediated by levodopa/benserazide in the brain of rats. J. Physiol. Sci. 2015, 65, 243–252. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Huang, D.; Hu, Q.; Wu, J.; Wang, Y.; Feng, J. Betaine alleviates hepatic lipid accumulation via enhancing hepatic lipid export and fatty acid oxidation in rats fed with a high-fat diet. Br. J. Nutr. 2015, 113, 1835–1843. [Google Scholar] [CrossRef] [PubMed]
- Dardi, F.; Manes, A.; Palazzini, M.; Bachetti, C.; Mazzanti, G.; Rinaldi, A.; Albini, A.; Gotti, E.; Monti, E.; Bacchi Reggiani, M.L. Combining bosentan and sildenafil in pulmonary arterial hypertension patients failing monotherapy: Real-world insights. Eur. Respir. J. 2015, 46, 414–421. [Google Scholar] [CrossRef] [PubMed]
- Nogueira-Ferreira, R.; Vitorino, R.; Ferreira, R.; Henriques-Coelho, T. Exploring the monocrotaline animal model for the study of pulmonary arterial hypertension: A network approach. Pulm. Pharmacol. Ther. 2015, 35, 8–16. [Google Scholar] [CrossRef] [PubMed]
- Polonio, I.B.; Acencio, M.M.; Pazetti, R.; Almeida, F.M.; Silva, B.S.; Pereira, K.A.; Souza, R. Lodenafil treatment in the monocrotaline model of pulmonary hypertension in rats. J. Bras. de Pneumol. 2014, 40, 421–424. [Google Scholar] [CrossRef]
- Medoff, B.D.; Okamoto, Y.; Leyton, P.; Weng, M.; Sandall, B.P.; Raher, M.J.; Kihara, S.; Bloch, K.D.; Libby, P.; Luster, A.D. Adiponectin deficiency increases allergic airway inflammation and pulmonary vascular remodeling. Am. J. Respir. Cell Mol. Biol. 2009, 41, 397–406. [Google Scholar] [CrossRef] [PubMed]
- Fuji, S.; Matsushita, S.; Hyodo, K.; Osaka, M.; Sakamoto, H.; Tanioka, K.; Miyakawa, K.; Kubota, M.; Hiramatsu, Y.; Tokunaga, C. Association between endothelial function and micro-vascular remodeling measured by synchrotron radiation pulmonary micro-angiography in pulmonary arterial hypertension. Gen. Thorac. Cardiovasc. Surg. 2016, 64, 597–603. [Google Scholar] [CrossRef] [PubMed]
- Abe, S.; Ishida, K.; Masuda, M.; Ueda, H.; Kohno, H.; Matsuura, K.; Tamura, Y.; Watanabe, M.; Matsumiya, G. A prospective, randomized study of inhaled prostacyclin versus nitric oxide in patients with residual pulmonary hypertension after pulmonary endarterectomy. Gen. Thorac. Cardiovasc. Surg. 2017, 65, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Cheng, Y.; Zong, L.; Huang, L.; Qiao, C.; Li, W.; Gong, B.; Hu, J.; Liu, H.; Wang, X.; et al. Aspirin attenuates monocrotaline-induced pulmonary arterial hypertension in rats by suppressing the ERK/MAPK pathway. Clin. Exp. Hypertens. 2017, 39, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Liu, Z.; Luo, Q.; Zhao, Z.; Zhao, Q.; Zheng, Y.; Xi, Q.; Tang, Y. Glycoprotein 130 Inhibitor Ameliorates Monocrotaline-Induced Pulmonary Hypertension in Rats. Can. J. Cardiol. 2016, 32, 1356.e1–1356.e10. [Google Scholar] [CrossRef] [PubMed]
- Feng, S.; Chen, S.; Yu, W.; Zhang, D.; Zhang, C.; Tang, C.; Du, J.; Jin, H. H2S inhibits pulmonary arterial endothelial cell inflammation in rats with monocrotaline-induced pulmonary hypertension. Lab. Investig. 2017, 97, 268–278. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Park, B.K.; Oh, K.S.; Yi, K.Y.; Lim, C.J.; Seo, H.W.; Lee, B.H. A urotensin II receptor antagonist, KR36676, decreases vascular remodeling and inflammation in experimental pulmonary hypertension. Int. Immunopharmacol. 2016, 40, 196–202. [Google Scholar] [CrossRef] [PubMed]
- Karin, M.; Lin, A. NF-κB at the crossroads of life and death. Nat. Immunol. 2002, 3, 221–227. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.Y.; Cai, Z.Y.; Pu, J.; Li, J.; Shen, J.Y.; Yang, C.D.; He, B. 5-Aminosalicylic Acid Attenuates Monocrotaline-Induced Pulmonary Arterial Hypertension in Rats by Increasing the Expression of Nur77. Inflammation 2017, 40, 806–817. [Google Scholar] [CrossRef] [PubMed]
- Soon, E.; Crosby, A.; Southwood, M.; Yang, P.; Tajsic, T.; Toshner, M.; Appleby, S.; Shanahan, C.M.; Bloch, K.D.; Pepke-Zaba, J.; et al. Bone morphogenetic protein receptor type II deficiency and increased inflammatory cytokine production. A gateway to pulmonary arterial hypertension. Am. J. Respir. Crit. Care Med. 2015, 192, 859–872. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Zhu, T.; Xiong, A.; Ge, X.; Xu, R.; Lu, S.; Hu, C. Calpain mediated pulmonary vascular remodeling in hypoxia induced pulmonary hypertension. Zhong Nan Da Xue Xue Bao Yi Xue Ban = J. Cent. South Univ. Med. Sci. 2016, 41, 929–936. [Google Scholar]
- Dai, Y.P.; Bongalon, S.; Hatton, W.J.; Hume, J.R.; Yamboliev, I.A. ClC-3 chloride channel is upregulated by hypertrophy and inflammation in rat and canine pulmonary artery. Br. J. Pharmacol. 2005, 145, 5–14. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Fang, C.; Cao, G.; Liu, K.; Wang, B.; Wan, Z.; Li, S.; Wu, S. Ethyl pyruvate ameliorates monocrotaline-induced pulmonary arterial hypertension in rats. J. Cardiovasc. Pharmacol. 2014, 64, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.; Zhang, Z.; Liu, T.; Zhang, W.; Liu, J.; Wang, W.; Wang, J. Salusin-beta contributes to vascular inflammation associated with pulmonary arterial hypertension in rats. J. Thorac. Cardiovasc. Surg. 2016, 152, 1177–1187. [Google Scholar] [CrossRef] [PubMed]
- Zimmer, A.; Teixeira, R.B.; Bonetto, J.H.; Siqueira, R.; Carraro, C.C.; Donatti, L.M.; Hickmann, A.; Litvin, I.E.; Godoy, A.E.; Araujo, A.S. Effects of aerobic exercise training on metabolism of nitric oxide and endothelin-1 in lung parenchyma of rats with pulmonary arterial hypertension. Mol. Cell. Biochem. 2017, 429, 73–89. [Google Scholar] [CrossRef] [PubMed]
- Rabinovitch, M.; Guignabert, C.; Humbert, M.; Nicolls, M.R. Inflammation and immunity in the pathogenesis of pulmonary arterial hypertension. Circ. Res. 2014, 115, 165–175. [Google Scholar] [CrossRef] [PubMed]
- Gruber, H.E.; Hoelscher, G.L.; Ingram, J.A.; Bethea, S.; Cox, M.; Hanley, E.N. Proinflammatory cytokines modulate the chemokine CCL2 (MCP-1) in human annulus cells in vitro: CCL2 expression and production. Exp. Mol. Pathol. 2015, 98, 102–105. [Google Scholar] [CrossRef] [PubMed]
- Bohm, F.; Pernow, J. The importance of endothelin-1 for vascular dysfunction in cardiovascular disease. Cardiovasc. Res. 2007, 76, 8–18. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.Y.; Tsai, K.B.; Hsu, J.H.; Shin, S.J.; Wu, J.R.; Yeh, J.L. Liraglutide prevents and reverses monocrotaline-induced pulmonary arterial hypertension by suppressing ET-1 and enhancing eNOS/sGC/PKG pathways. Sci. Rep. 2016, 6, 31788. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.J.; Chen, X.L.; Che, C.; Ai, J.; Li, J.; Han, X.J. Alterations in pulmonary arterial reactivity during pulmonary arterial hypertension at the early-stage of pulmonary fibrosis in rats. Zhongguo Ying Yong Sheng Li Xue Za Zhi = Chin. J. Appl. Physiol. 2011, 27, 110–114. [Google Scholar]
- Yeager, M.E.; Belchenko, D.D.; Nguyen, C.M.; Colvin, K.L.; Ivy, D.D.; Stenmark, K.R. Endothelin-1, the unfolded protein response, and persistent inflammation: Role of pulmonary artery smooth muscle cells. Am. J. Respir. Cell Mol. Biol. 2012, 46, 14–22. [Google Scholar] [CrossRef] [PubMed]
- Li, X.W.; Li, X.H.; Du, J.; Li, D.; Li, Y.J.; Hu, C.P. Calcitonin gene-related peptide down-regulates bleomycin-induced pulmonary fibrosis. Can. J. Physiol. Pharmacol. 2016, 94, 1315–1324. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Yuan, W.; Lv, C.; Li, N.; Liu, T.; Wang, L.; Sun, Y.; Qiu, X.; Fu, Q. Dihydroartemisinin supresses inflammation and fibrosis in bleomycine-induced pulmonary fibrosis in rats. Int. J. Clin. Exp. Pathol. 2015, 8, 1270–1281. [Google Scholar] [PubMed]
- Xiong, S.; Guo, R.; Yang, Z.; Xu, L.; Du, L.; Li, R.; Xiao, F.; Wang, Q.; Zhu, M.; Pan, X. Treg depletion attenuates irradiation-induced pulmonary fibrosis by reducing fibrocyte accumulation, inducing Th17 response, and shifting IFN-gamma, IL-12/IL-4, IL-5 balance. Immunobiology 2015, 220, 1284–1291. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.; Zhou, X.; Zhang, J.; Huang, X.; Zhai, Y.; Zhang, X.; Chu, L. Hydrogen sulfide protects against bleomycin-induced pulmonary fibrosis in rats by inhibiting NF-κB expression and regulating Th1/Th2 balance. Toxicol. Lett. 2014, 224, 387–394. [Google Scholar] [CrossRef] [PubMed]
- Cicko, S.; Grimm, M.; Ayata, K.; Beckert, J.; Meyer, A.; Hossfeld, M.; Zissel, G.; Idzko, M.; Müller, T. Uridine supplementation exerts anti-inflammatory and anti-fibrotic effects in an animal model of pulmonary fibrosis. Respir. Res. 2015, 16, 105. [Google Scholar] [CrossRef] [PubMed]
- Adegunsoye, A.; Balachandran, J. Inflammatory response mechanisms exacerbating hypoxemia in coexistent pulmonary fibrosis and sleep apnea. Mediators Inflamm. 2015, 2015, 510105. [Google Scholar] [CrossRef] [PubMed]
- Vizza, C.D.; Sastry, B.K.; Safdar, Z.; Harnisch, L.; Gao, X.; Zhang, M.; Lamba, M.; Jing, Z.C. Efficacy of 1, 5, and 20 mg oral sildenafil in the treatment of adults with pulmonary arterial hypertension: A randomized, double-blind study with open-label extension. BMC Pulm. Med. 2017, 17, 44. [Google Scholar] [CrossRef] [PubMed]
- Barst, R.J.; Beghetti, M.; Pulido, T.; Layton, G.; Konourina, I.; Zhang, M.; Ivy, D.D. STARTS-2: Long-term survival with oral sildenafil monotherapy in treatment-naive pediatric pulmonary arterial hypertension. Circulation 2014, 129, 1914–1923. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.C.; Jiang, F.M.; Zheng, Q.L.; Li, C.T.; Peng, X.Y.; He, C.Y.; Luo, J.; Liang, Z.A. Efficacy and safety of sildenafil treatment in pulmonary arterial hypertension: A systematic review. Respir. Med. 2014, 108, 531–537. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors. |









© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, J.-m.; Zhou, R.; Zhang, M.; Tan, H.-r.; Yu, J.-q. Betaine Attenuates Monocrotaline-Induced Pulmonary Arterial Hypertension in Rats via Inhibiting Inflammatory Response. Molecules 2018, 23, 1274. https://doi.org/10.3390/molecules23061274
Yang J-m, Zhou R, Zhang M, Tan H-r, Yu J-q. Betaine Attenuates Monocrotaline-Induced Pulmonary Arterial Hypertension in Rats via Inhibiting Inflammatory Response. Molecules. 2018; 23(6):1274. https://doi.org/10.3390/molecules23061274
Chicago/Turabian StyleYang, Jia-mei, Ru Zhou, Min Zhang, Huan-ran Tan, and Jian-qiang Yu. 2018. "Betaine Attenuates Monocrotaline-Induced Pulmonary Arterial Hypertension in Rats via Inhibiting Inflammatory Response" Molecules 23, no. 6: 1274. https://doi.org/10.3390/molecules23061274
APA StyleYang, J.-m., Zhou, R., Zhang, M., Tan, H.-r., & Yu, J.-q. (2018). Betaine Attenuates Monocrotaline-Induced Pulmonary Arterial Hypertension in Rats via Inhibiting Inflammatory Response. Molecules, 23(6), 1274. https://doi.org/10.3390/molecules23061274