Synthesis, Biological Evaluation, and Docking Studies of Novel Bisquaternary Aldoxime Reactivators on Acetylcholinesterase and Butyrylcholinesterase Inhibited by Paraoxon

 , , ,
, , ,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
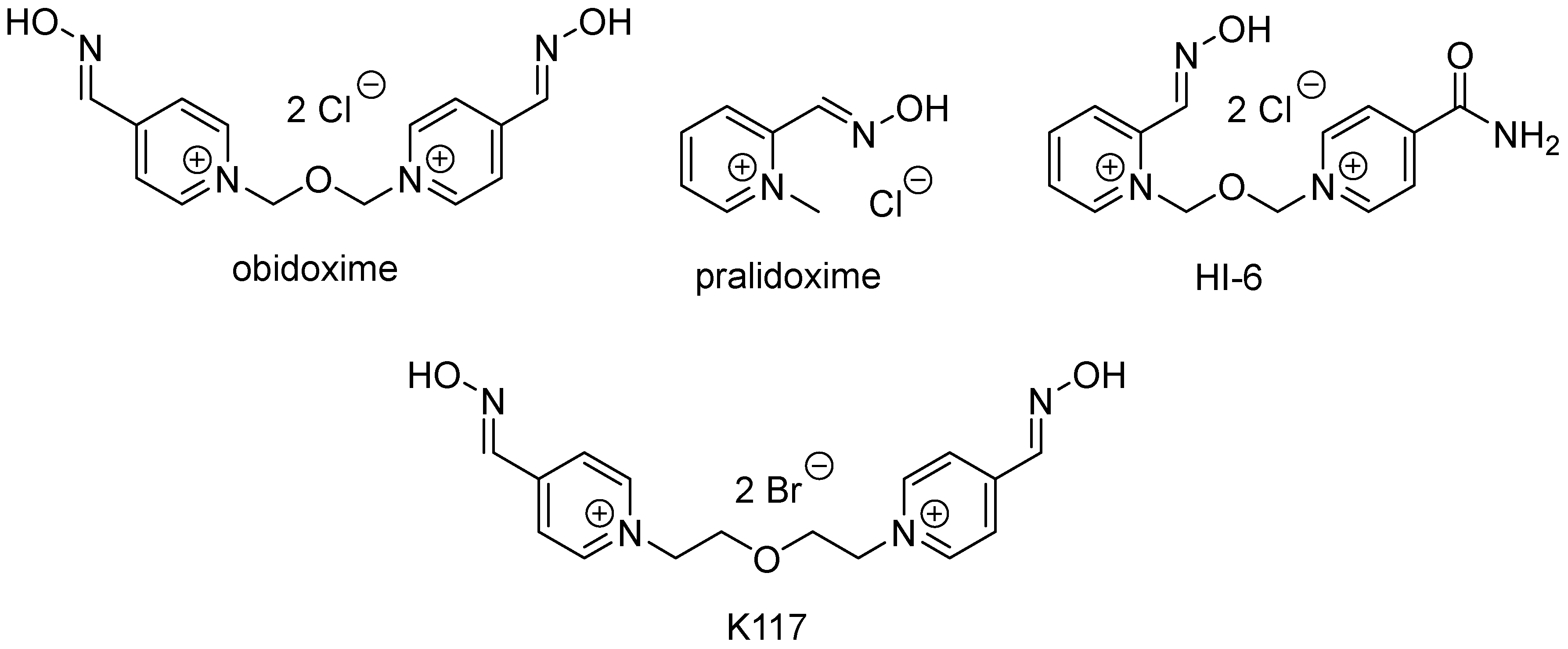
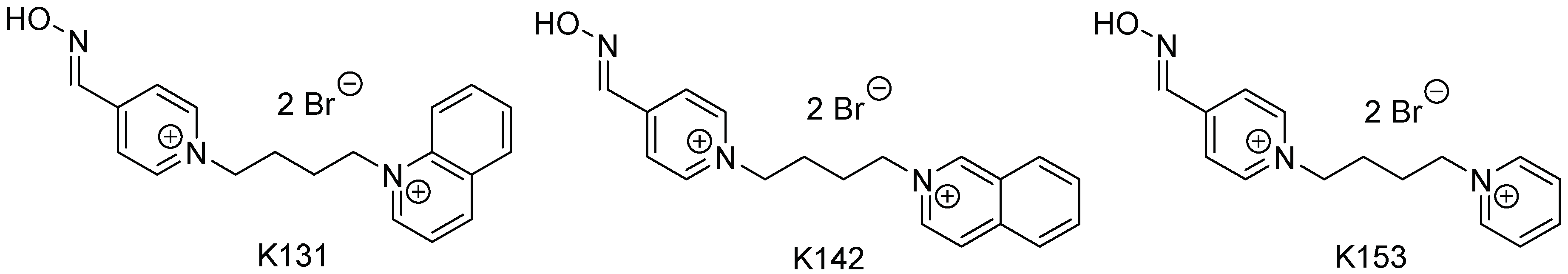
2.1. Synthesis and Characterization of the New Oximes
2.2. Biochemical Experiments
2.3. Molecular Modeling Studies
3. Results and Discussion
3.1. Characterization Data of the New Oximes
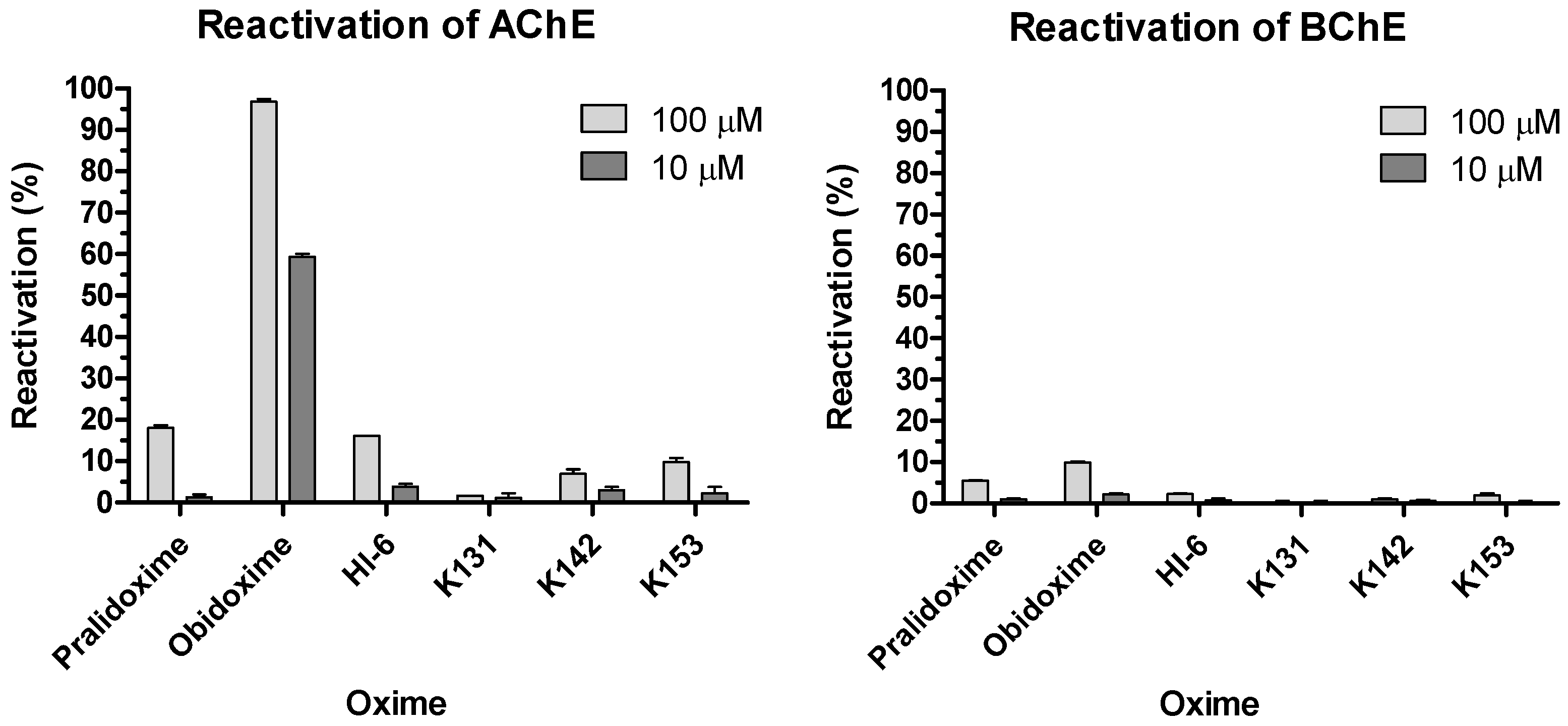
3.2. Biochemical Experiments
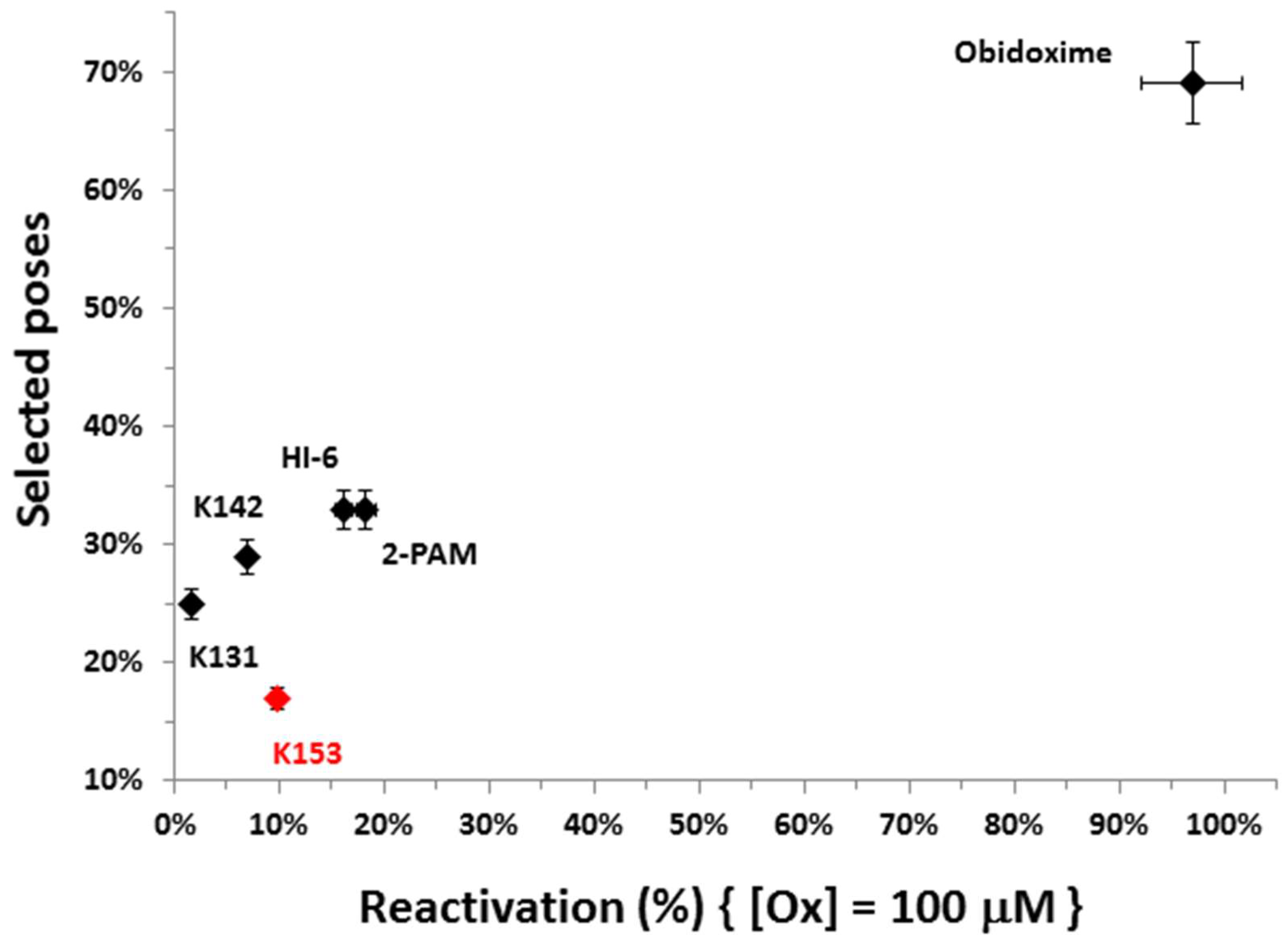
3.3. Docking Studies
4. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Newmark, J. Therapy for Nerve Agent Poisoning. Arch. Neurol. 2004, 61, 649–652. [Google Scholar] [CrossRef] [PubMed]
- Gorecki, L.; Korabecny, J.; Musilek, K.; Malinak, D.; Nepovimova, E.; Dolezal, R.; Jun, D.; Soukup, O.; Kuca, K. SAR study to find optimal cholinesterase reactivator against organophosphorous nerve agents and pesticides. Arch. Toxicol. 2016, 90, 2831–2859. [Google Scholar] [CrossRef] [PubMed]
- Carletti, E.; Aurbek, N.; Gillon, E.; Loiodice, M.; Nicolet, Y.; Fontecilla-Camps, J.-C.; Masson, P.; Thiermann, H.; Nachon, F.; Worek, F. Structure-Activity Analysis of Aging and Reactivation of Human Butyrylcholinesterase Inhibited by Analogues of Tabun. Biochem. J. 2009, 412, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Musilova, L.; Jun, D.; Kuca, K.; Pohanka, M.; Katalinic, M.; Kovarik, Z. Development of New Antidotes of Organophosphate Intoxications: Oxime-Assisted Reactivation of Dimethoxy- and Diethoxy-Phosphorylated Human Butyrylcholinesterase for Construction of “pseudo Catalytic” Bioscavengers. Toxicol. Lett. 2009, 189, S216. [Google Scholar] [CrossRef]
- Jun, D.; Musilova, L.; Kuca, K.; Kassa, J.; Bajgar, J. Potency of Several Oximes to Reactivate Human Acetylcholinesterase and Butyrylcholinesterase Inhibited by Paraoxon in Vitro. Chem. Biol. Interact. 2008, 175, 421–424. [Google Scholar] [CrossRef] [PubMed]
- Wiesner, J.; Kriz, Z.; Kuca, K.; Jun, D.; Koca, J. Why Acetylcholinesterase Reactivators Do Not Work in Butyrylcholinesterase. J. Enzyme Inhib. Med. Chem. 2010, 25, 318–322. [Google Scholar] [CrossRef] [PubMed]
- Musilova, L.; Kuca, K.; Jung, Y.-S.; Jun, D. In Vitro Oxime-Assisted Reactivation of Paraoxon-Inhibited Human Acetylcholinesterase and Butyrylcholinesterase. Clin. Toxicol. 2009, 47, 545–550. [Google Scholar] [CrossRef] [PubMed]
- Kuca, K.; Jun, D.; Musilek, K. Structural Requirements of Acetylcholinesterase Reactivators. Mini-Rev. Med. Chem. 2006, 6, 269–277. [Google Scholar] [CrossRef] [PubMed]
- Kuca, K.; Jun, D.; Bajgar, J. Currently Used Cholinesterase Reactivators against Nerve Agent Intoxication: Comparison of Their Effectivity in Vitro. Drug Chem. Toxicol. 2007, 30, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Binder, J.; Paar, M.; Jun, D.; Pohanka, M.; Hrabinova, M.; Opletalova, V.; Kuca, K. New Bisquaternary Isoquinolinium Inhibitors of Brain Cholinesterases-Synthesis and Anticholinesterase Activity. Lett. Drug Des. Discov. 2010, 7, 1–4. [Google Scholar]
- Komloova, M.; Horova, A.; Hrabinova, M.; Jun, D.; Dolezal, R.; Vinsova, J.; Kuca, K.; Musilek, K. Preparation, in Vitro Evaluation and Molecular Modelling of Pyridinium-Quinolinium/isoquinolinium Non-Symmetrical Bisquaternary Cholinesterase Inhibitors. Bioorg. Med. Chem. Lett. 2013, 23, 6663–6666. [Google Scholar] [CrossRef] [PubMed]
- Nepovimova, E.; Korabecny, J.; Dolezal, R.; Nguyen, T.D.; Jun, D.; Soukup, O.; Pasdiorova, M.; Jost, P.; Muckova, L.; Malinak, D.; et al. A 7-methoxytacrine–4-pyridinealdoxime hybrid as a novel prophylactic agent with reactivation properties in organophosphate intoxication. Toxicol. Res. 2016, 5, 1012–1016. [Google Scholar] [CrossRef]
- Jun, D.; Kuca, K.; Stodulka, P.; Koleckar, V.; Dolezal, R.; Simon, P.; Veverka, M. HPLC Analysis of HI-6 Dichloride and Dimethanesulfonate-Antidotes against Nerve Agents and Organophosphorus Pesticides. Anal. Lett. 2007, 40, 2783–2787. [Google Scholar] [CrossRef]
- Jun, D.; Stodulka, P.; Kuca, K.; Koleckar, V.; Dolezal, B.; Simon, P.; Veverka, M. TLC Analysis of Intermediates Arising during the Preparation of Oxime HI-6 Dimethanesulfonate. J. Chromatogr. Sci. 2008, 46, 316–319. [Google Scholar] [CrossRef] [PubMed]
- Jun, D.; Musilova, L.; Musilek, K.; Kuca, K. In Vitro Ability of Currently Available Oximes to Reactivate Organophosphate Pesticide-Inhibited Human Acetylcholinesterase and Butyrylcholinesterase. Int. J. Mol. Sci. 2011, 12, 2077–2087. [Google Scholar] [CrossRef] [PubMed]
- Ellman, G.; Courtney, K.; Andres, V.; Featherstone, R. A New and Rapid Colorimetric Determination of Acetylcholinesterase Activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef]
- Deppmeier, B.J.; Driessen, A.J.; Hehre, W.J.; Johnson, J.A.; Klunzinger, P.E.; Watanabe, M.; Yu, J. PC Spartan Pro, Version 1.0.5; Wavefunction Inc.: Irvine, CA, 2000. [Google Scholar]
- Dewar, M.J.S.; Zoebisch, E.G.; Healy, E.F.; Stewart, J.J.P. AM1: A New General Purpose Quantum Mechanical Molecular Model. J. Am. Chem. Soc. 1985, 107, 3902–3909. [Google Scholar] [CrossRef]
- Swain, M. Chemicalize.org. J. Chem. Inf. Mod. 2012, 52, 613–615. [Google Scholar] [CrossRef]
- Discovery Studio Modeling Environment; Release 4.5; BIOVIA, Dassault Systèmes: San Diego, CA, USA, 2015.
- Almeida, J.S.F.A.; Guizado, T.R.C.; Guimaraes, A.P.; Ramalho, T.C.; Gonçalves, A.S.; Koning, M.C.; França, T.C.C. Docking and molecular dynamics studies of peripheral site ligand-oximes as reactivators of sarin-inhibited human acetylcholinesterase. J. Biomol. Struc. Dyn. 2016, 34, 2632–2642. [Google Scholar] [CrossRef] [PubMed]
- Thomsen, R.; Christensen, M.H. MolDock: A new technique for high-accuracy molecular docking. J. Med. Chem. 2006, 49, 3315–3321. [Google Scholar] [CrossRef] [PubMed]
- Kovarik, Z.; Vrdoljak, A.L.; Berend, S.; Katalinić, M.; Kuca, K.; Musilek, K.; Radić, B. Evaluation of Oxime k203 as Antidote in Tabun Poisoning. Arh. Hig. Rada Toksikol. 2009, 60, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Musilova, L.; Jun, D.; Palecek, J.; Cirkva, V.; Musilek, K.; Paar, M.; Hrabinova, M.; Pohanka, M.; Kuca, K. Novel Nucleophilic Compounds with Oxime Group as Reactivators of Paraoxon-Inhibited Cholinesterases. Lett. Drug Des. Discov. 2010, 7, 260–264. [Google Scholar] [CrossRef]
- Kontoyianni, M.; McClellan, L.M.; Sokol, G.S. Evaluation of docking performance: Comparative data on docking algorithms. J. Med. Chem. 2004, 47, 558–565. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of all compounds are available from the authors. |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reactivator | Reactivation (%) | |||||||
|---|---|---|---|---|---|---|---|---|
| HssAChE | HssBChE | |||||||
| 100 µM | 10 µM | 100 µM | 10 µM | |||||
| Mean | SD | Mean | SD | Mean | SD | Mean | SD | |
| pralidoxime | 18.2 | 0.7 | 1.3 | 0.7 | 5.5 | 0.1 | 1.0 | 0.2 |
| obidoxime | 96.9 | 0.7 | 59.4 | 0.7 | 9.9 | 0.3 | 2.2 | 0.3 |
| HI-6 | 16.1 | 0.0 | 3.9 | 0.7 | 2.3 | 0.2 | 0.8 | 0.4 |
| K131 | 1.7 | 0.0 | 1.1 | 1.1 | 0.3 | 0.3 | 0.3 | 0.3 |
| K142 | 7.0 | 1.0 | 3.0 | 0.8 | 1.1 | 0.2 | 0.6 | 0.3 |
| K153 | 9.8 | 1.0 | 2.3 | 1.5 | 2.0 | 0.5 | 0.3 | 0.3 |
| Oxime | The Best Pose (↓dOP*/↑θOPO*) | |||||
|---|---|---|---|---|---|---|
| dOP (Å) | Angle OPO | Energy of Interaction (kcal/mol) | Energy of H-bond (kcal/mol) | Interactions: H-bond | Interactions: Hydrophobic (π–π) | |
| pralidoxime | 8.644 | 153.35° | −73.444 | −5.153 | Tyr124, Val294/Phe295, Phe295/Arg296 | Tyr72, Tyr124, Trp286,Phe295, Phe297, Tyr337, Phe338, Tyr341 |
| obidoxime | 4.260 | 155.00° | −105.013 | −4.941 | Tyr124, Tyr337 Ser203–POX | Tyr72, Trp86, Tyr124, Trp286, Phe295, Phe297, Tyr337, Phe338, Tyr341, His447 |
| HI-6 | 7.102 | 147.64° | −135.261 | −6.397 | Tyr124, Val294/Phe295, Val282/Asn283 | Tyr72, Trp86, Tyr124, His284, Trp286, Phe295, Phe297, Tyr337, Phe338, Tyr341 |
| K131 | 4.111 | 144.47° | −136.854 | −2.500 | Ser203–POX | Tyr72, Tyr124, His284, Trp286, His287, Phe295, Phe297, Tyr337, Phe338, Tyr341, His447 |
| K142 | 3.864 | 141.70° | −129.978 | −3.586 | Tyr72,Ser203–POX | Tyr72, Tyr124, His284, Trp286, His287, Phe295, Phe297, Tyr337, Phe338, Tyr341 |
| K153 | 6.777 | 147.81° | −119.168 | −2.445 | Val294/Phe295 | Tyr72, Tyr124, His284, Trp286, His287, Phe295, Phe297, Phe338, Tyr341 |
| Oxime | The Best Pose (↓dOP/↑θOPO) | |||||
|---|---|---|---|---|---|---|
| dOP (Å) | θOPO | Energy of Interaction (kcal/mol) | Energy of H-bond (kcal/mol) | Interactions: H-bond | Interactions: Hydrophobic (π–π) | |
| pralidoxime | 5.600 | 148.21° | −57.770 | −2.370 | Thr120 | Phe118, Phe329, Tyr332 |
| obidoxime | 5.623 | 168.15° | −135.823 | −9.600 | Gly115/Gly116, Tyr128, Tyr332 | Phe73, Trp82, Tyr114, Tyr128, Phe329, Tyr332, Trp430, Tyr440, His438 |
| HI-6 | 5.469 | 140.81° | −126.712 | −4.851 | Thr120, His438/Gly439 | Trp82, Phe118, Phe329, Tyr332, Trp430, Tyr440, His438 |
| K131 | 5.587 | 146.97° | −118.023 | −2.859 | Thr120 | Trp82, Phe329, Tyr332, His438 |
| K142 | 5.172 | 144.04° | −115.435 | −2.431 | Thr120 | Trp82, Phe118, Phe329, Tyr332, His438 |
| K153 | 5.414 | 133.13° | −109.310 | −3.860 | Thr120 | Trp82, Phe118, Phe329, Tyr332 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kuca, K.; Jun, D.; Junova, L.; Musilek, K.; Hrabinova, M.; Da Silva, J.A.V.; Ramalho, T.C.; Valko, M.; Wu, Q.; Nepovimova, E.; et al. Synthesis, Biological Evaluation, and Docking Studies of Novel Bisquaternary Aldoxime Reactivators on Acetylcholinesterase and Butyrylcholinesterase Inhibited by Paraoxon. Molecules 2018, 23, 1103. https://doi.org/10.3390/molecules23051103
Kuca K, Jun D, Junova L, Musilek K, Hrabinova M, Da Silva JAV, Ramalho TC, Valko M, Wu Q, Nepovimova E, et al. Synthesis, Biological Evaluation, and Docking Studies of Novel Bisquaternary Aldoxime Reactivators on Acetylcholinesterase and Butyrylcholinesterase Inhibited by Paraoxon. Molecules. 2018; 23(5):1103. https://doi.org/10.3390/molecules23051103
Chicago/Turabian StyleKuca, Kamil, Daniel Jun, Lucie Junova, Kamil Musilek, Martina Hrabinova, Jorge Alberto Valle Da Silva, Teodorico Castro Ramalho, Marian Valko, Qinghua Wu, Eugenie Nepovimova, and et al. 2018. "Synthesis, Biological Evaluation, and Docking Studies of Novel Bisquaternary Aldoxime Reactivators on Acetylcholinesterase and Butyrylcholinesterase Inhibited by Paraoxon" Molecules 23, no. 5: 1103. https://doi.org/10.3390/molecules23051103