Molecular Sex Identification in Dioecious Hippophae rhamnoides L. via RAPD and SCAR Markers

 ,
,
Abstract
:1. Introduction
2. Results
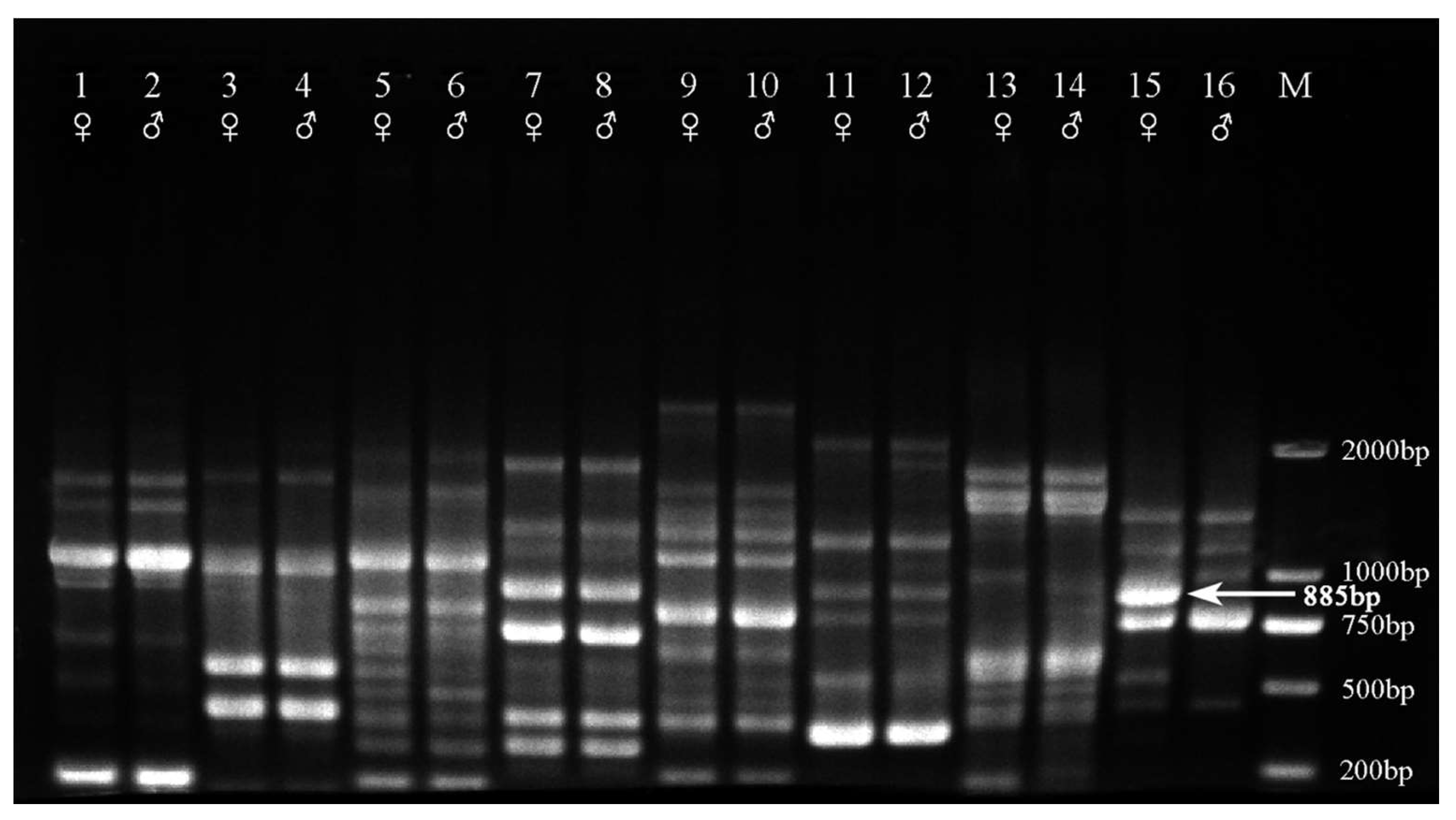
2.1. Sex-Linked RAPD Marker
2.2. DNA Sequence of Female-Specific Fragments
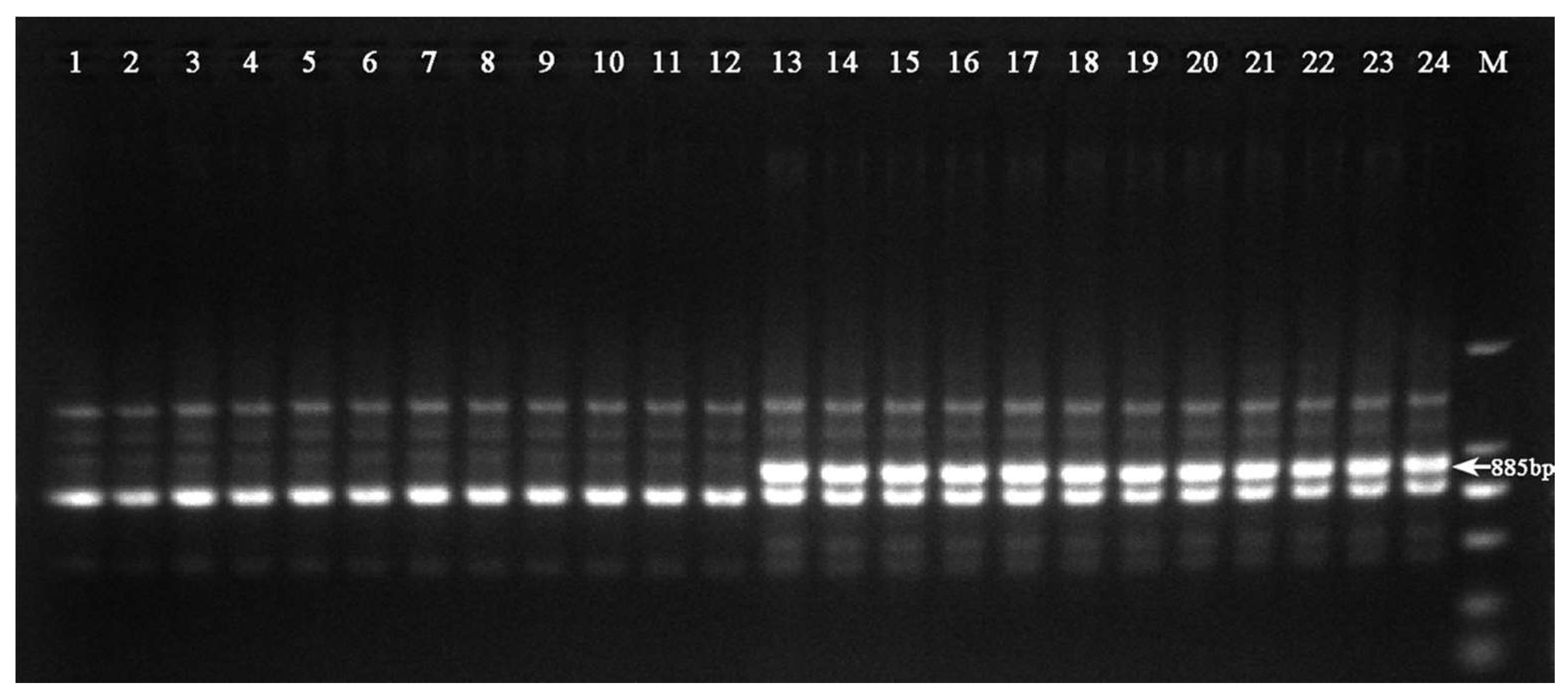
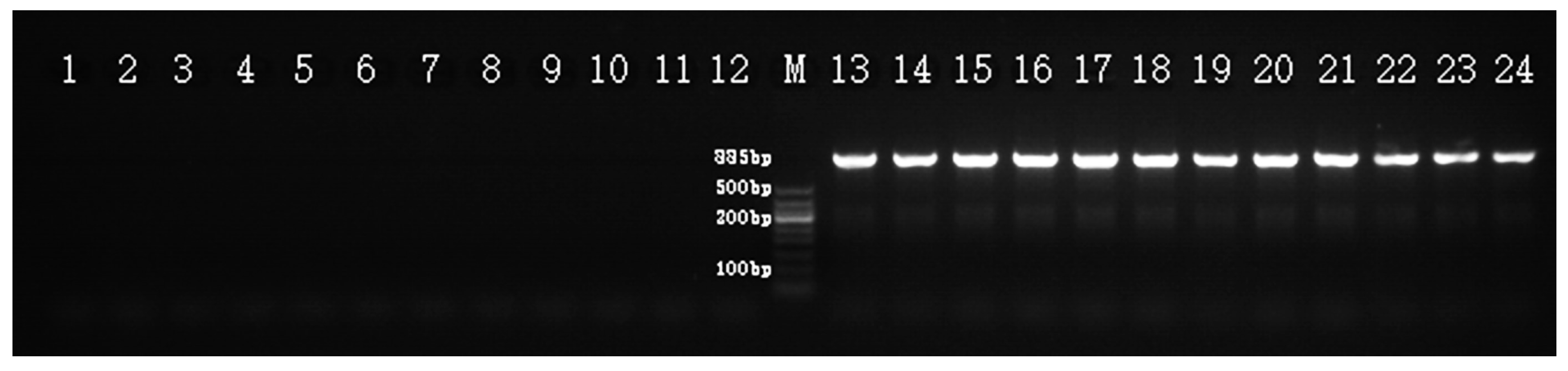
2.3. SCAR Marker Development and Examination
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. DNA Extraction
4.3. RAPD Amplification
4.4. Cloning of the RAPD Amplicon
4.5. SCAR Marker Development and Examination
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Ruan, C.J.; Xie, Q.L. Preliminary report on seabuckthorn variety introduction at beach in Vancheng City. Hippophae 2001, 14, 15–19. [Google Scholar]
- Fu, L.; Su, H.; Li, R.; Cui, Y. Harvesting technologies for sea buckthorn fruit. Engineering in Agriculture. Environ. Food 2014, 7, 64–69. [Google Scholar]
- Renner, S.S.; Ricklefs, R.E. Dioecy and its correlations in the flowering plants. Am. J. Bot. 1995, 82, 596–606. [Google Scholar] [CrossRef]
- Charlesworth, D. Plant sex chromosomes. Annu. Rev. Plant Biol. 2016, 67, 397–420. [Google Scholar] [CrossRef] [PubMed]
- Ming, R.; Bendahmane, A.; Renner, S.S. Sex chromosomes in land plants. Annu. Rev. Plant Biol. 2011, 62, 485–514. [Google Scholar] [CrossRef] [PubMed]
- Vyskot, B.; Hobza, R. The genomics of plant sex chromosomes. Plant Sci. 2015, 236, 126–135. [Google Scholar] [CrossRef] [PubMed]
- Westergaard, M. The mechanism of sex determination in flowering plants. Adv. Genet. 1958, 9, 217–281. [Google Scholar] [PubMed]
- Liu, Z.; Moore, P.H.; Ma, H.; Ackerman, C.M.; Ragiba, M.; Yu, Q.; Pearl, H.M.; Kim, M.S.; Charlton, J.W.; Stiles, J.I.; et al. A primitive Y chromosome in papaya marks incipient sex chromosome evolution. Nature 2004, 427, 348–352. [Google Scholar] [CrossRef] [PubMed]
- Kejnovsky, E.; Vyskot, B. Silene latifolia: The classical model to study heteromorphic sex chromosomes. Cytogenet. Genome Res. 2010, 129, 250–262. [Google Scholar] [CrossRef] [PubMed]
- Papadopulos, A.S.T.; Chester, M.; Ridout, K.; Filatov, D.A. Rapid Y degeneration and dosage compensation in plant sex chromosomes. Proc. Natl. Acad. Sci. USA 2015, 112, 13021–13026. [Google Scholar] [CrossRef] [PubMed]
- Ainsworth, C. Boys and Girls Come Out to Play: The Molecular Biology of Dioecious Plants. Ann. Bot. 2000, 86, 211–221. [Google Scholar] [CrossRef]
- Matsunaga, S.; Kawano, S. Sex Determination by Sex Chromosomes in Dioecious Plants. Plant Biol. 2001, 3, 481–488. [Google Scholar] [CrossRef]
- Charlesworth, D. Plant sex determination and sex chromosomes. Heredity 2002, 88, 94–101. [Google Scholar] [CrossRef] [PubMed]
- Negrutiu, I.; Vyskot, B.; Barbacar, N.; Georgiev, S.; Moneger, F. Dioecious plants. A key to the early events of sex chromosome evolution. Plant Physiol. 2001, 127, 1418–1424. [Google Scholar] [CrossRef] [PubMed]
- Tanurdzic, M.; Banks, J.A. Sex-determining mechanisms in land plants. Plant Cell 2004, 16 (Suppl. 1), S61–S71. [Google Scholar] [CrossRef] [PubMed]
- Irish, E.E.; Nelson, T. Sex Determination in Monoecious and Dioecious Plants. Plant Cell 1989, 1, 737–744. [Google Scholar] [CrossRef] [PubMed]
- Durand, R.; Durand, B.; Michel, J.P.D. Sexual determination and sexual differentiation. Crit. Rev. Plant Sci. 1990, 9, 295–316. [Google Scholar] [CrossRef]
- Anastassopoulos, E. The Origins of Genome Architecture; Sinauer Associates: Sunderland, MA, USA, 2007; pp. 491–493. [Google Scholar]
- Pannell, J. Mixed genetic and environmental sex determination in an androdioecious population of Mercurialis annua. Heredity 1997, 78, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Elena, T.; Capraru, G.; Rosu, C.M.; Zamfirache, M.M.; Olteanu, Z.; Manzu, C. Morphometric pattern of somatic chromosomes in three Romanian seabuckthorn genotypes. Caryologia 2011, 64, 189–196. [Google Scholar] [CrossRef]
- Khadke, G.N.; Hima, B.K.; Ravishankar, K.V. Development of SCAR marker for sex determination in dioecious betelvine (Piper betle L.). Curr. Sci. 2012, 103, 712–716. [Google Scholar]
- Masayuki, M. Development of SCAR markers for sex determination in the dioecious shrub Aucuba japonica (Cornaceae). Genome 2009, 52, 231–237. [Google Scholar]
- Liao, L.; Liu, J.; Dai, Y.; Li, Q.; Xie, M.; Chen, Q.; Yin, H.; Qiu, G.; Liu, X. Development and application of SCAR markers for sex identification in the dioecious species Ginkgo biloba L. Euphytica 2009, 169, 49–55. [Google Scholar] [CrossRef]
- Ruan, C.J.; Xie, Q.L.; Li, D.Q. Function and benefits of seabuckthorn improving eco environment of loess plateaus. Environ. Prot. 2000, 5, 30–31. [Google Scholar]
- Goel, H.C.; Prasad, J.; Singh, S.; Sagar, R.K.; Kumar, I.P.; Sinha, A.K. Radioprotection by a herbal preparation of Hippophae rhamnoides, RH-3, against whole body lethal irradiation in mice. Phytomedicine 2001, 9, 15–25. [Google Scholar] [CrossRef]
- Teng, B.S.; Lu, Y.H.; Wang, Z.T.; Tao, X.Y.; Wei, D.Z. In vitro anti-tumor activity of isorhamnetin isolated from Hippophae rhamnoides L. against BEL-7402 cells. Pharmacol. Res. 2006, 54, 186–194. [Google Scholar] [CrossRef] [PubMed]
- Mishra, K.P.; Chanda, S.; Karan, D.; Ganju, L.; Sawhney, R.C. Effect of Seabuckthorn (Hippophae rhamnoides) flavone on immune system: An in-vitro approach. Phytother. Res. PTR 2008, 22, 1490–1495. [Google Scholar] [CrossRef] [PubMed]
- Hwang, I.S.; Kim, J.E.; Choi, S.I.; Lee, H.R.; Lee, Y.J.; Jang, M.J. UV radiation-induced skin aging in hairless mice is effectively prevented, by oral intake of sea buckthorn (Hippophae rhamnoides L.) fruit blend for 6 weeks, through MMP suppression and increase of sod activity. Int. J. Mol. Med. 2012, 30, 392–400. [Google Scholar] [CrossRef] [PubMed]
- Shinwari, Z.K.; Gilani, S.S. Sustainable harvest of medicinal plants at Bulashbar Nullah, Astore (northern Pakistan). J. Ethnopharmacol. 2003, 84, 289–298. [Google Scholar] [CrossRef]
- Walf, D.; Wegart, F. Experience gained in the harvesting and utilization of seabuckthorn. In Cultivation and Utilization of Wild Fruit Crops; Bernhard Thalacker Veriage Gmbh & Co.: Braunschweig, Germany, 1993; pp. 22–29. [Google Scholar]
- Persson, A.H.; Nybom, H. Genetic Sex Determination and RAPD Marker Segregation in the Dioecious Species Sea Buckthorn (Hippophae Rhamnoides L.). Hereditas 1998, 129, 45–51. [Google Scholar] [CrossRef]
- Satender, R.; Poonam, S.; Yadav, M.C. A female sex associated randomly amplified polymorphic DNA marker in dioecious Hippophae salicifolia. Genes Genomes Genom. 2009, 3, 96–101. [Google Scholar]
- Sharma, A.; Zinta, G.; Rana, S.; Shirko, P. Molecular identification of sex in Hippophae rhamnoides L. using isozyme and RAPD markers. For. Ecosyst. 2010, 12, 62–66. [Google Scholar] [CrossRef]
- Korekar, G.; Sharma, R.K.; Kumar, R.; Meenu; Bisht, N.C.; Srivastava, R.B.; Ahuja, P.S.; Stobdan, T. Identification and validation of sex-linked SCAR markers in dioecious Hippophae rhamnoides L. (Elaeagnaceae). Biotechnol. Lett. 2012, 34, 973–978. [Google Scholar] [CrossRef] [PubMed]
- Hormaza, J.I.; Dollo, L.; Polito, V.S. Identification of a RAPD marker linked to sex determination in Pistacia vera using bulk segregant analysis. Theor. Appl. Genet. 1994, 89, 9–13. [Google Scholar] [CrossRef] [PubMed]
- Polley, A.; Seigner, E.; Ganal, M.W. Identification of sex in hop (Humulus lupulus) using molecular markers. Genome 1997, 40, 357–361. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.; Sink, K.C. RAPD and SCAR markers linked to the sex expression locus M in asparagus. Euphytica 1997, 94, 329–333. [Google Scholar] [CrossRef]
- Kalia, R.K.; Singh, R.; Rai, M.K.; Mishra, G.P.; Singh, S.R.; Dhawan, A.K. Biotechnological interventions in sea buckthorn (Hippophae L.): Current status and future prospects. Trees 2011, 25, 559–575. [Google Scholar] [CrossRef]
- Williams, J.G.; Kubelik, A.R.; Livak, K.J.; Rafalski, J.A.; Tingey, S.V. DNA polymorphisms amplified by arbitrary primers are useful as genetic markers. Nucleic Acids Res. 1990, 18, 6531–6535. [Google Scholar] [CrossRef] [PubMed]
- Robles, J.; Doers, M. pGEM®-T Vector Systems troubleshooting guide. Promega Notes 1994, 45, 19–20. [Google Scholar]
Sample Availability: Samples of the compounds are available from the authors. |





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Region | Elevation (m) | Longitude | Latitude |
|---|---|---|---|
| Mayigou reservoir, Huangzhong County | 2644 | E101°36′26.39′′ | N36°29'34.76′′ |
| Dahua Village, Huangyuan County | 2745 | E101°1058.77′′ | N36°41′20.60′′ |
| Chuchuer village, Pingan County | 2875 | E101°51′14.50′′ | N36°23′33.24′′ |
| Xiangyi village, Longhua County | 2933 | E102°02′40.21′′ | N36°12′11.84′′ |
| Tangfang village, Xunhua County | 2544 | E102°22′34.12′′ | N35°49′41.65′′ |
| Tiegeleng village, Xunhua County | 2840 | E102°37′27.01′′ | N35°42′06.18′′ |
| LangTang Village, Minhe County | 2636 | E102°44′10.64′′ | N36°02′59.05′′ |
| Xiaoshuiquan village, Pingan County | 2556 | E101°57′57.64′′ | N36°29′29.05′′ |
| Pandao village, Huangzhong County | 2816 | E101°21′03.40′′ | N36°34′49.34′′ |
| Youning temple, Minhe County | 3007 | E102°11′36.45′′ | N36°45′10.16′′ |
| Bianmagou Village, Datong County | 3010 | E101°50′40.05′′ | N36°57′45.36′′ |
| Double-tree village, Huzhu County | 2436 | E101°55′01.91′′ | N36°46′47.96′′ |
| Chengguan nursery, Datong County | 2566 | E101°33′12.67′′ | N37°01′55.57′′ |
| BaoKu Township, Datong County | 2683 | E101°33′59.65′′ | N37°06′09.15′′ |
| Botanical Garden, Xining City | 2314 | E101°44′38.09′′ | N36°37'29.17′′ |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, W.; Wang, Y.; Zhang, G.; Luan, G.; Chen, S.; Meng, J.; Wang, H.; Hu, N.; Suo, Y. Molecular Sex Identification in Dioecious Hippophae rhamnoides L. via RAPD and SCAR Markers. Molecules 2018, 23, 1048. https://doi.org/10.3390/molecules23051048
Zhou W, Wang Y, Zhang G, Luan G, Chen S, Meng J, Wang H, Hu N, Suo Y. Molecular Sex Identification in Dioecious Hippophae rhamnoides L. via RAPD and SCAR Markers. Molecules. 2018; 23(5):1048. https://doi.org/10.3390/molecules23051048
Chicago/Turabian StyleZhou, Wu, Yuwei Wang, Gong Zhang, Guangxiang Luan, Shasha Chen, Jing Meng, Honglun Wang, Na Hu, and Yourui Suo. 2018. "Molecular Sex Identification in Dioecious Hippophae rhamnoides L. via RAPD and SCAR Markers" Molecules 23, no. 5: 1048. https://doi.org/10.3390/molecules23051048
APA StyleZhou, W., Wang, Y., Zhang, G., Luan, G., Chen, S., Meng, J., Wang, H., Hu, N., & Suo, Y. (2018). Molecular Sex Identification in Dioecious Hippophae rhamnoides L. via RAPD and SCAR Markers. Molecules, 23(5), 1048. https://doi.org/10.3390/molecules23051048