
Pursuing Intracellular Pathogens with Hyaluronan. From a ‘Pro-Infection’ Polymer to a Biomaterial for ‘Trojan Horse’ Systems

 ,
,
Abstract
:1. Introduction
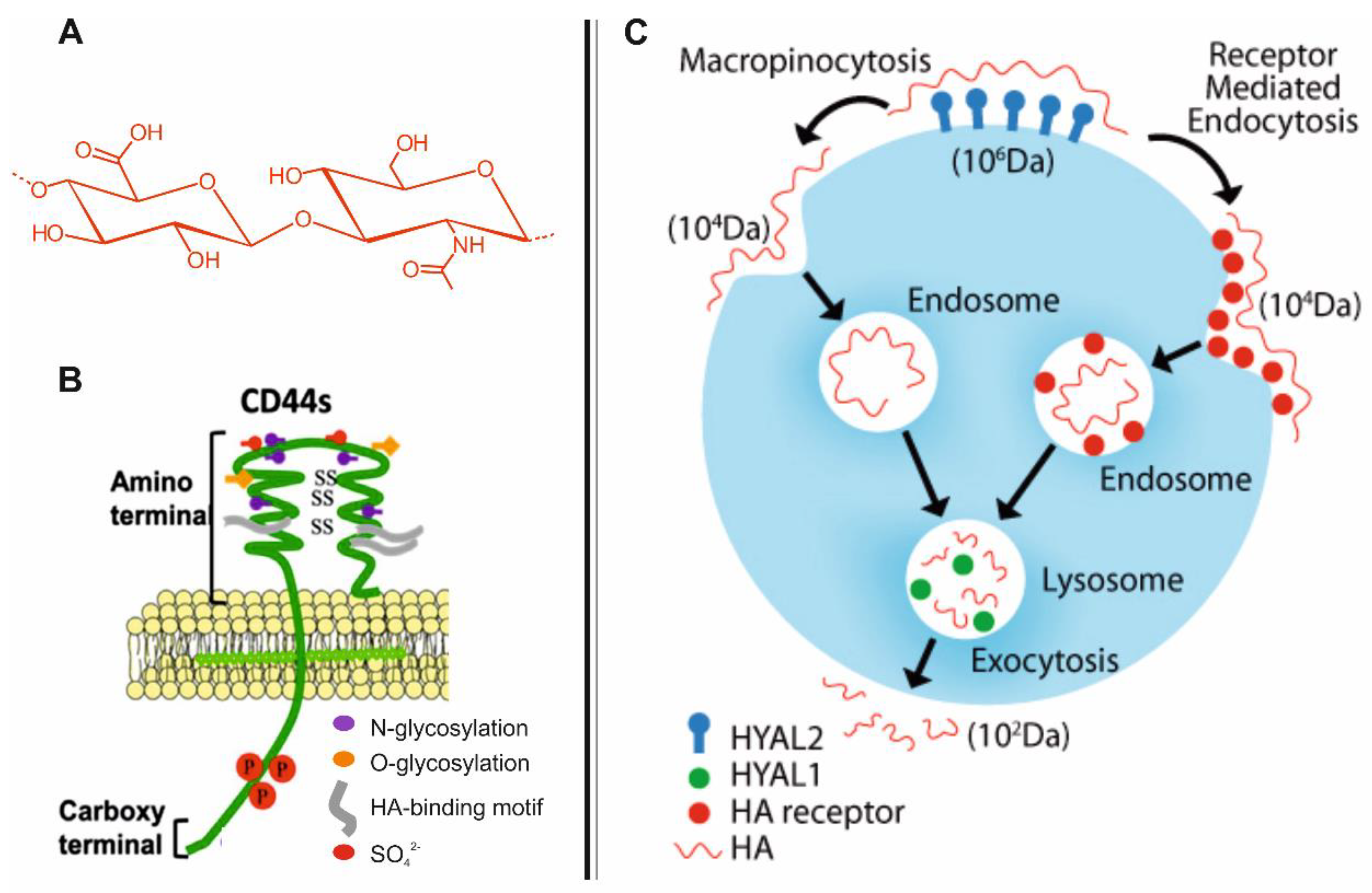
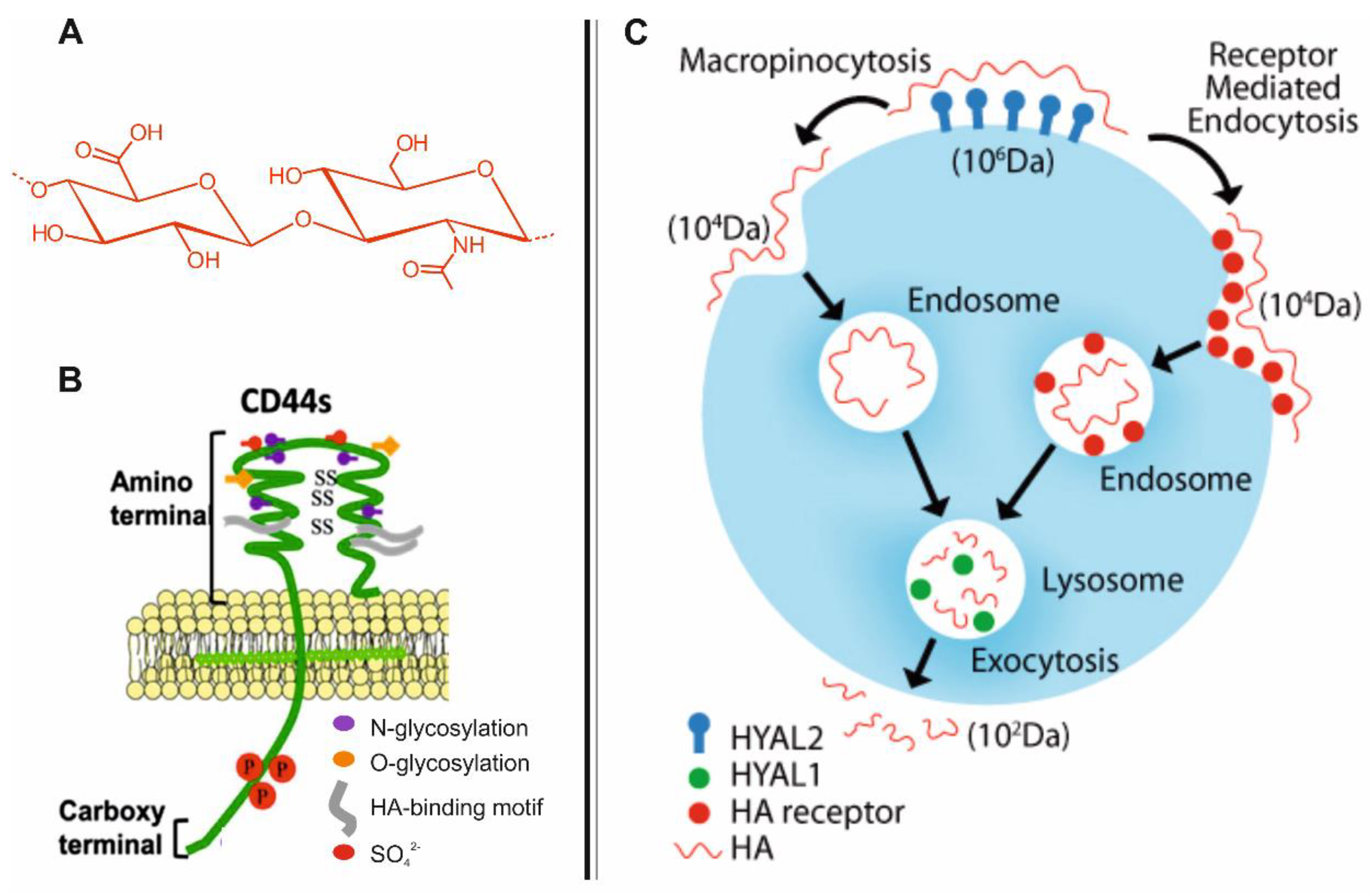
2. Biodistribution and Roles of HA
3. CD44-Mediated Uptake of HA in Host Cells
4. Role of CD44 and HA in the Uptake and Proliferation of Intracellular Pathogens
5. HA-Based Nano-Carriers in Drug Delivery
6. HA-Based Nano-Carriers for Targeting Sub-Cellular Compartments
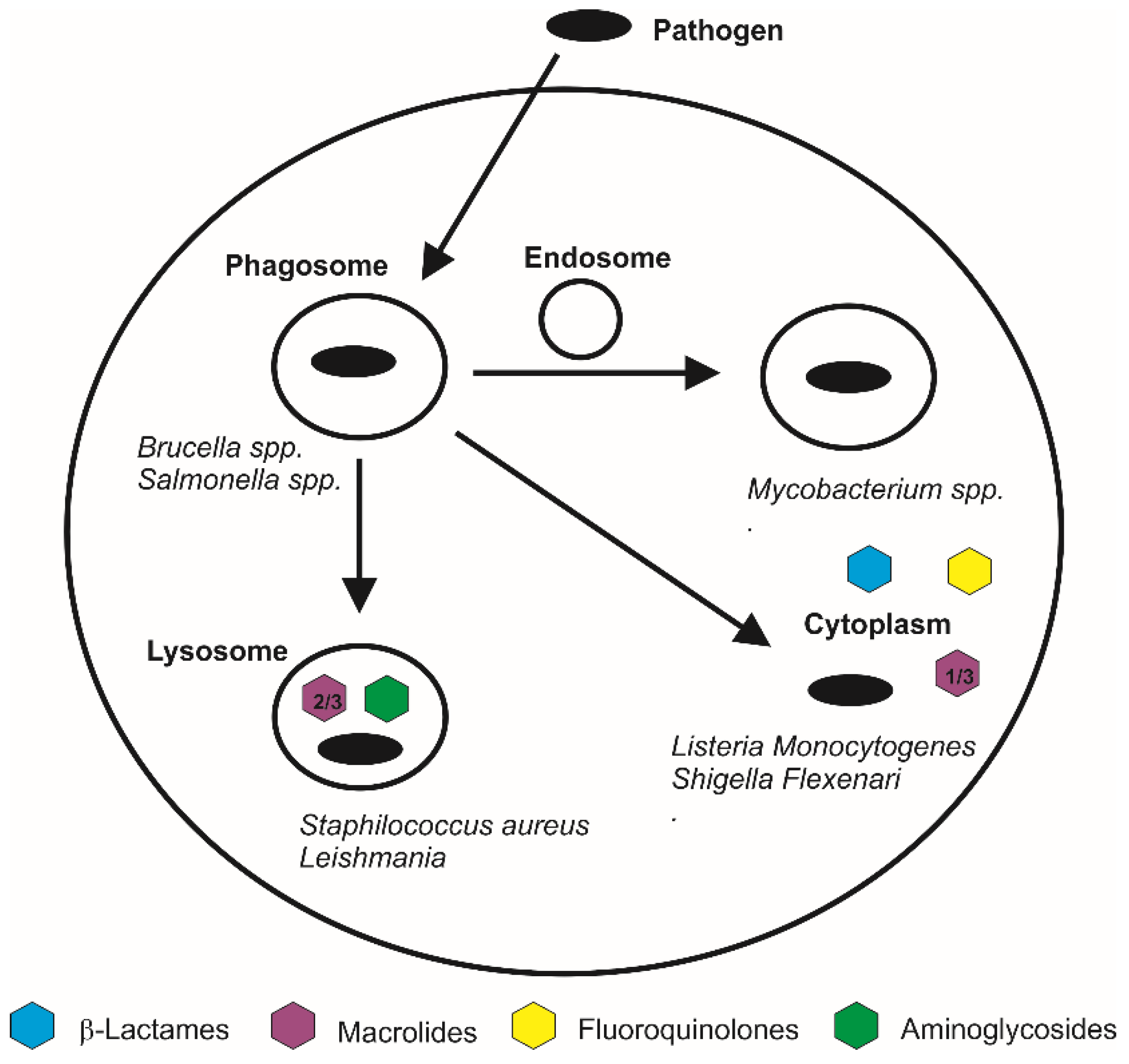
6.1. Lysosomal HA-Based Nano-Carriers
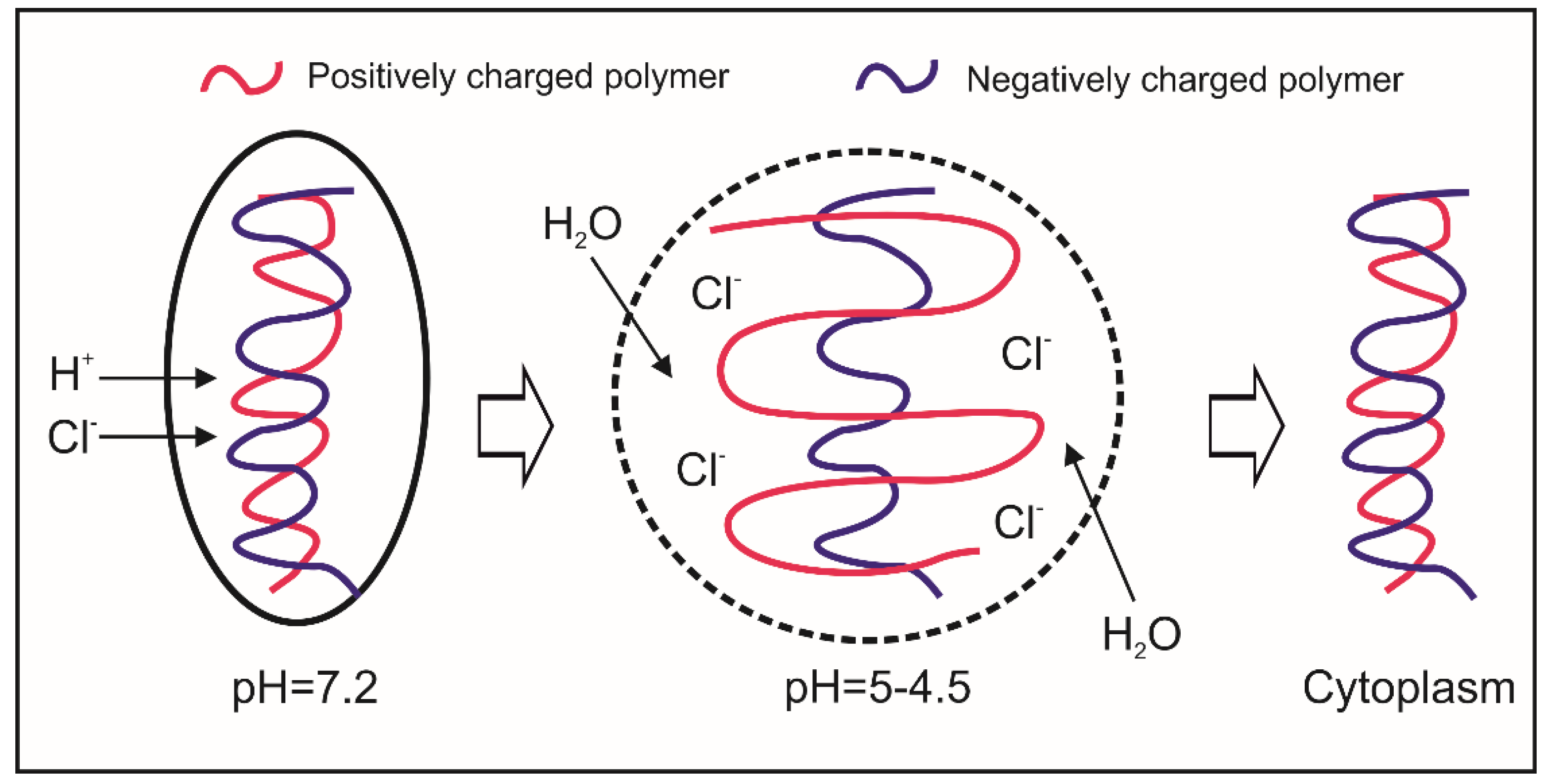
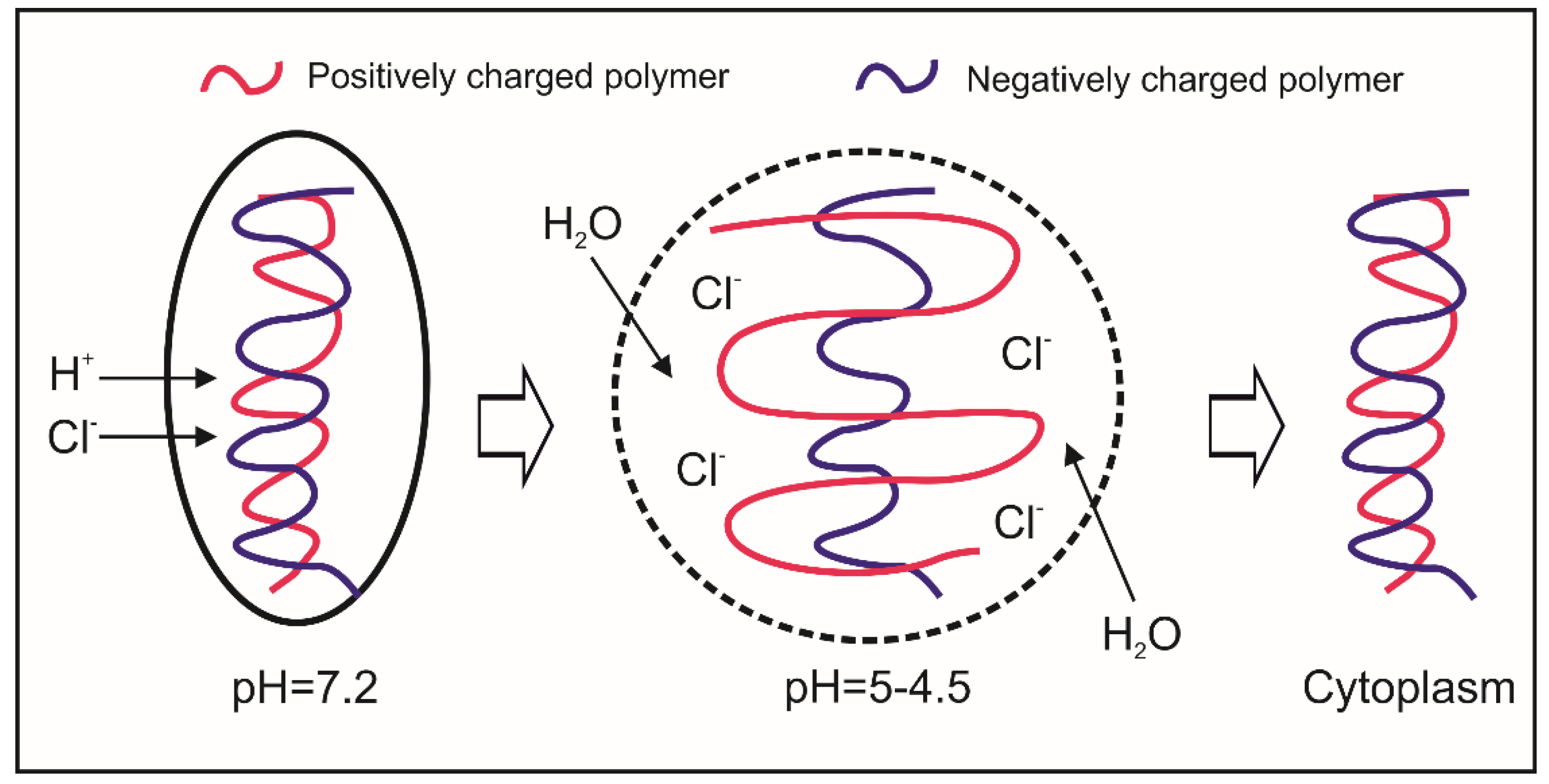
6.2. Cytosolic HA-Based Nano-Carriers
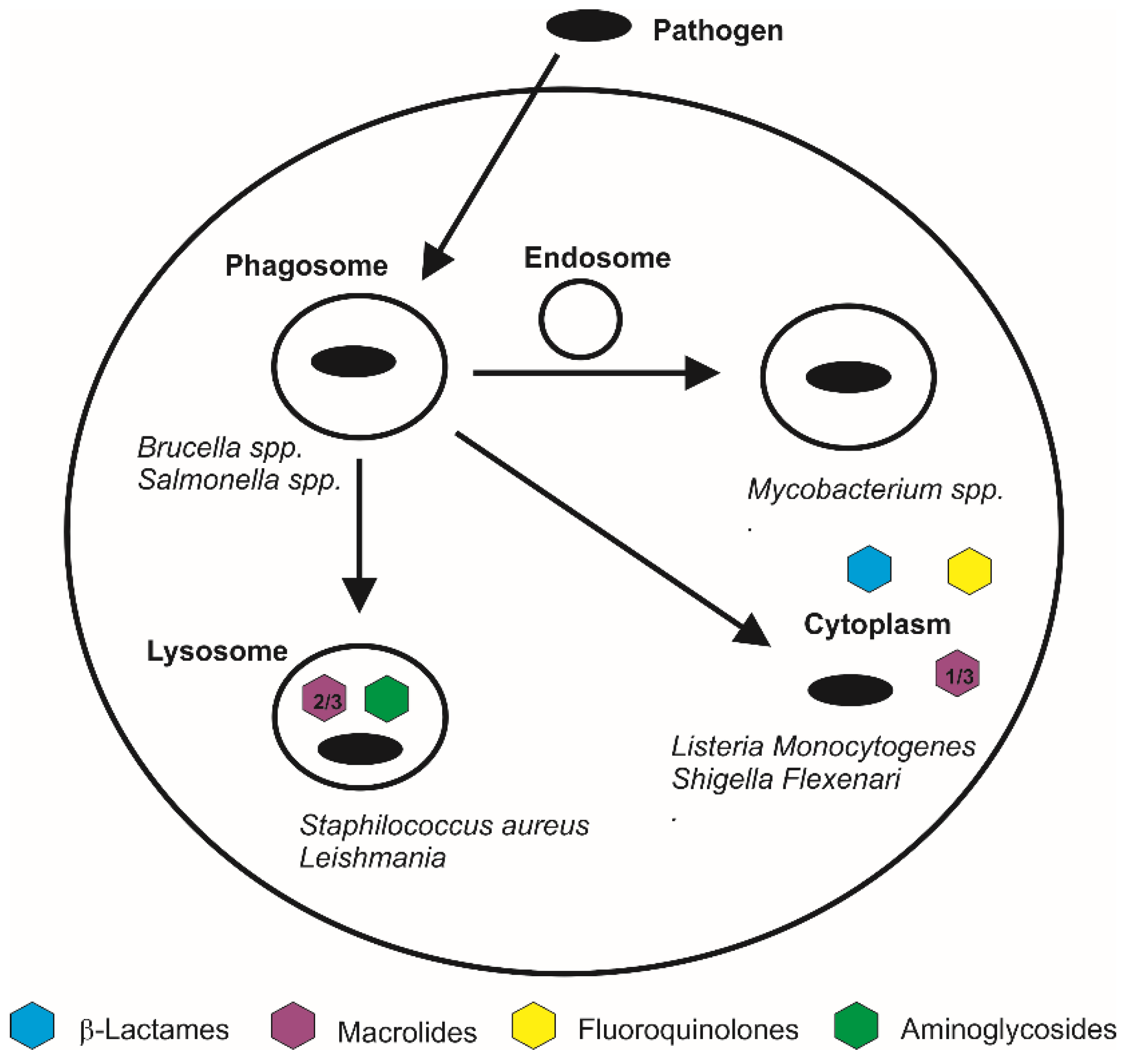
7. The Application of HA-Based Nano-Carriers for the Intracellular Delivery of Antimicrobials
8. Conclusions and Perspectives
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Casadevall, A. Evolution of Intracellular Pathogens. Annu. Rev. Microbiol. 2008, 62, 19–33. [Google Scholar] [CrossRef] [PubMed]
- Pizarro-Cerdá, J.; Cossart, P. Bacterial Adhesion and Entry into Host Cells. Cell 2006, 124, 715–727. [Google Scholar] [CrossRef] [PubMed]
- Aquino, R.S.; Park, P.W. Glycosaminoglycans and infection. Front. Biosci. 2016, 21, 1260–1277. [Google Scholar]
- Schrager, H.M.; Albertì, S. Hyaluronic acid capsule modulates M protein-mediated adherence and acts as a ligand for attachment of group A Streptococcus to CD44 on human keratinocytes. J. Clin. Investig. 1998, 101, 1708–1716. [Google Scholar] [CrossRef] [PubMed]
- Aoki, K.; Matsumoto, S. Extracellular Mycobacterial DNA-binding Protein 1 Participates in Mycobacterium-Lung Epithelial Cell Interaction through Hyaluronic Acid. J. Biol. Chem. 2004, 279, 39798–39806. [Google Scholar] [CrossRef] [PubMed]
- Naderer, T.; Heng, J. Intracellular Survival of Leishmania major Depends on Uptake and Degradation of Extracellular Matrix Glycosaminoglycans by Macrophages. PLoS Pathog. 2015, 11, e1005136. [Google Scholar] [CrossRef] [PubMed]
- Leemans, J.C.; Florquin, S. CD44 is a macrophage binding site for Mycobacterium tuberculosis that mediates macrophage recruitment and protective immunity against tuberculosis. J. Clin. Investig. 2003, 111, 681–689. [Google Scholar] [CrossRef] [PubMed]
- Moffat, F.L.; Han, T. Involvement of CD44 and the cytoskeletal linker protein ankyrin in human neutrophil bacterial phagocytosis. J. Cell. Physiol. 1996, 168, 638–647. [Google Scholar] [CrossRef]
- Eriksson, E.; Dons, L. CD44-Regulated Intracellular Proliferation of Listeria monocytogenes. Infect. Immun. 2003, 71, 4102–4111. [Google Scholar] [CrossRef] [PubMed]
- Meyer, K.; Palmer, J.W. The polysaccharide of the vitreous humor. J. Biol. Chem. 1934, 107, 629–634. [Google Scholar]
- Rapport, M.M.; Weissmann, B. Isolation of a Crystalline Disaccharide, Hyalobiuronic Acid, from Hyaluronic Acid. Nature 1951, 168, 205–211. [Google Scholar] [CrossRef]
- Laurent, U.B.G.; Fraser, J.R.E. Turnover of hyaluronate in the aqueous humour and vitreous body of the rabbit. Exp. Eye Res. 1983, 36, 493–503. [Google Scholar] [CrossRef]
- Balazs, E.A.; Laurent, T.C. Nomenclature of hyaluronic acid. Biochem. J. 1986, 235, 903. [Google Scholar] [CrossRef] [PubMed]
- Fakhari, A.; Berkland, C. Applications and emerging trends of hyaluronic acid in tissue engineering, as a dermal filler and in osteoarthritis treatment. Acta Biomater. 2013, 9, 7081–7092. [Google Scholar] [CrossRef] [PubMed]
- Stern, R.; Kogan, G. The many ways to cleave hyaluronan. Biotechnol. Adv. 2007, 25, 537–557. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Crewe, C. Hyaluronan in adipose tissue: Beyond dermal filler and therapeutic carrier. Sci. Transl. Med. 2016, 8, 323ps4. [Google Scholar] [CrossRef] [PubMed]
- Endre, A.B. Hyaluronan as an Ophthalmic Viscoelastic Device. Curr. Pharm. Biotechnol. 2008, 9, 236–238. [Google Scholar]
- Ponta, H.; Sherman, L. CD44: From adhesion molecules to signalling regulators. Nat. Rev. Mol. Cell Biol. 2003, 4, 33–45. [Google Scholar] [CrossRef] [PubMed]
- Racine, R.; Mummert, M.E. Hyaluronan Endocytosis: Mechanisms of Uptake and Biological Functions. In Molecular Regulation of Endocytosis; InTech: London, UK, 2012; ISBN 978-953-51-0662-3. [Google Scholar]
- Tammi, R.; Rilla, K. Hyaluronan Enters Keratinocytes by a Novel Endocytic Route for Catabolism. J. Biol. Chem. 2001, 276, 35111–35122. [Google Scholar] [CrossRef] [PubMed]
- Underhill, C.B.; Nguyen, H.A. CD44 Positive Macrophages Take up Hyaluronan during Lung Development. Dev. Biol. 1993, 155, 324–336. [Google Scholar] [CrossRef] [PubMed]
- Bertolami, C.N.; Berg, S. Binding and Internalization of Hyaluronate by Human Cutaneous Fibroblasts. Matrix 1992, 12, 11–21. [Google Scholar] [CrossRef]
- Aguiar, D.J.; Knudson, W. Internalization of the Hyaluronan Receptor CD44 by Chondrocytes. Exp. Cell Res. 1999, 252, 292–302. [Google Scholar] [CrossRef] [PubMed]
- Culty, M.; Shizari, M. Binding and degradation of hyaluronan by human breast cancer cell lines expressing different forms of CD44: Correlation with invasive potential. J. Cell. Physiol. 1994, 160, 275–286. [Google Scholar] [CrossRef] [PubMed]
- Fraser, J.R.E.; Laurent, T.C. Hyaluronan: Its nature, distribution, functions and turnover. J. Intern. Med. 1997, 242, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Toole, B.P. Hyaluronan: From extracellular glue to pericellular cue. Nat. Rev. Cancer 2004, 4, 528–539. [Google Scholar] [CrossRef] [PubMed]
- Weigel, P.H.; Hascall, V.C. Hyaluronan Synthases. J. Biol. Chem. 1997, 272, 13997–14000. [Google Scholar] [CrossRef] [PubMed]
- Itano, N.; Kimata, K. Molecular Cloning of Human Hyaluronan Synthase. Biochem. Biophys. Res. Commun. 1996, 222, 816–820. [Google Scholar] [CrossRef] [PubMed]
- Itano, N.; Sawai, T. Three Isoforms of Mammalian Hyaluronan Synthases Have Distinct Enzymatic Properties. J. Biol. Chem. 1999, 274, 25085–25092. [Google Scholar] [CrossRef] [PubMed]
- Stern, R. Hyaluronan catabolism: A new metabolic pathway. Eur. J. Cell Biol. 2004, 83, 317–325. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Chang, M.C.Y. The Hyaluronan Receptor RHAMM Regulates Extracellular-regulated Kinase. J. Biol. Chem. 1998, 273, 11342–11348. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, M.; Assmann, V. Problems with RHAMM. Cell 1998, 95, 591–592. [Google Scholar] [CrossRef]
- Stern, R. Devising a pathway for hyaluronan catabolism: Are we there yet? Glycobiology 2003, 13, 105R–115R. [Google Scholar] [CrossRef] [PubMed]
- Rice, K.G. The Chemistry, Biology, and Medical Applications of Hyaluronan and Its Derivatives. J. Med. Chem. 1998, 41, 5336. [Google Scholar] [CrossRef]
- Zhou, B.; Weigel, J.A. Identification of the Hyaluronan Receptor for Endocytosis (HARE). J. Biol. Chem. 2000, 275, 37733–37741. [Google Scholar] [CrossRef] [PubMed]
- Banerji, S.; Ni, J. LYVE-1, a New Homologue of the CD44 Glycoprotein, Is a Lymph-specific Receptor for Hyaluronan. J. Cell Biol. 1999, 144, 789–801. [Google Scholar] [CrossRef] [PubMed]
- Stern, R.; Asari, A.A. Hyaluronan fragments: An information-rich system. Eur. J. Cell Biol. 2006, 85, 699–715. [Google Scholar] [CrossRef] [PubMed]
- Day, A.J.; Prestwich, G.D. Hyaluronan-binding proteins: Tying up the giant. J. Biol. Chem. 2002, 277, 4585–4588. [Google Scholar] [CrossRef] [PubMed]
- Tamer, T.M. Hyaluronan and synovial joint: Function, distribution and healing. Interdiscip. Toxicol. 2013, 6, 111–125. [Google Scholar] [CrossRef] [PubMed]
- Moskowitz, R.W. Hyaluronic acid supplementation. Curr. Rheumatol. Rep. 2000, 2, 466–471. [Google Scholar] [CrossRef] [PubMed]
- Feinberg, R.; Beebe, D. Hyaluronate in vasculogenesis. Science 1983, 220, 1177–1179. [Google Scholar] [CrossRef] [PubMed]
- Delmage, J.; Powars, D.R. The selective suppression of immunogenicity by hyaluronic acid. Ann. Clin. Lab. Sci. 1986, 16, 303–310. [Google Scholar] [PubMed]
- Powell, J.D.; Horton, M.R. Threat matrix. Immunol. Res. 2005, 31, 207–218. [Google Scholar] [CrossRef]
- Nikitovic, D.; Tzardi, M. Cancer Microenvironment and Inflammation: Role of Hyaluronan. Front. Immunol. 2015, 6, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Hascall, V.C.; Majors, A.K. Intracellular hyaluronan: A new frontier for inflammation? Biochim. Biophys. Acta 2004, 1673, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Termeer, C.; Sleeman, J.P. Hyaluronan-magic glue for the regulation of the immune response? Trends Immunol. 2003, 24, 112–114. [Google Scholar] [CrossRef]
- Abatangelo, G.; Cortivo, R. Cell detachment mediated by hyaluronic acid. Exp. Cell Res. 1982, 137, 73–78. [Google Scholar] [CrossRef]
- Koochekpour, S.; Pilkington, G.J. Hyaluronic acid/CD44H interaction induces cell detachment and stimulates migration and invasion of human glioma cells in vitro. Int. J. Cancer 1995, 63, 450–454. [Google Scholar] [CrossRef] [PubMed]
- Assmann, V.; Fieber, C. CD44 is the Principal Mediator of Hyaluronic-Acid-Induced Melanoma Cell Proliferation. J. Investig. Dermatol. 2001, 116, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Jiang, D.; Liang, J.; Fan, J.; Yu, S.; Chen, S.; Luo, Y.; Prestwich, G.D.; Mascarenhas, M.M.; Garg, H.G.; Quinn, D.A.; et al. Regulation of lung injury and repair by Toll-like receptors and hyaluronan. Nat. Med. 2005, 11, 1173–1179. [Google Scholar] [CrossRef] [PubMed]
- Teder, P.; Vandivier, R.W.; Jiang, D.; Liang, J.; Cohn, L.; Puré, E.; Henson, P.M.; Noble, P.W. Resolution of lung inflammation by CD44. Science 2002, 296, 155–158. [Google Scholar] [CrossRef] [PubMed]
- Zöller, M. CD44: Can a cancer-initiating cell profit from an abundantly expressed molecule? Nat. Rev. Cancer 2011, 11, 254–267. [Google Scholar] [CrossRef] [PubMed]
- Sleeman, J.P.; Kondo, K. Variant Exons v6 and v7 Together Expand the Repertoire of Glycosaminoglycans Bound by CD44. J. Biol. Chem. 1997, 272, 31837–31844. [Google Scholar] [CrossRef] [PubMed]
- Stamenkovic, I.; Amiot, M. A lymphocyte molecule implicated in lymph node homing is a member of the cartilage link protein family. Cell 1989, 56, 1057–1062. [Google Scholar] [CrossRef]
- Idzerda, R.L.; Carter, W.G. Isolation and DNA sequence of a cDNA clone encoding a lymphocyte adhesion receptor for high endothelium. PNAS 1989, 86, 4659–4663. [Google Scholar] [CrossRef] [PubMed]
- Thankamony, S.P.; Knudson, W. Acylation of CD44 and Its Association with Lipid Rafts Are Required for Receptor and Hyaluronan Endocytosis. J. Biol. Chem. 2006, 281, 34601–34609. [Google Scholar] [CrossRef] [PubMed]
- Oliferenko, S.; Paiha, K. Analysis of CD44-Containing Lipid Rafts. Recruitment of Annexin II and Stabilization by the Actin Cytoskeleton. J. Cell Biol. 1999, 146, 843–854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, D.; Sy, M.S. A cysteine residue located in the transmembrane domain of CD44 is important in binding of CD44 to hyaluronic acid. J. Exp. Med. 1996, 183, 1987–1994. [Google Scholar] [CrossRef] [PubMed]
- McGary, C.T.; Raja, R.H. Endocytosis of hyaluronic acid by rat liver endothelial cells. Evidence for receptor recycling. Biochem. J. 1989, 257, 875–884. [Google Scholar] [CrossRef] [PubMed]
- Culty, M.; O’Mara, T.E. Hyaluronan receptor (CD44) expression and function in human peripheral blood monocytes and alveolar macrophages. J. Leukoc. Biol. 1994, 56, 605–611. [Google Scholar] [CrossRef] [PubMed]
- Cywes, C.; Stamenkovic, I. CD44 as a receptor for colonization of the pharynx by group A Streptococcus. J. Clin. Investig. 2000, 106, 995–1002. [Google Scholar] [CrossRef] [PubMed]
- Skoudy, A.; Mounier, J. CD44 binds to the Shigella IpaB protein and participates in bacterial invasion of epithelial cells. Cell. Microbiol. 2000, 2, 19–33. [Google Scholar] [CrossRef] [PubMed]
- Hirayama, Y.; Yoshimura, M. Mycobacteria Exploit Host Hyaluronan for Efficient Extracellular Replication. PLoS Pathog. 2009, 5, e1000643. [Google Scholar] [CrossRef] [PubMed]
- Li, M.W.; Yudin, A.I.; VandeVoort, C.A.; Sabeur, K.; Primakoff, P.; Overstreet, J.W. Inhibition of monkey sperm hyaluronidase activity and heterologous cumulus penetration by flavonoids. Biol. Reprod. 1997, 56, 1383–1389. [Google Scholar] [CrossRef] [PubMed]
- Nagy, N.; Kuipers, H.F.; Frymoyer, A.R.; Ishak, H.D.; Bollyky, J.B.; Wight, T.N.; Bollyky, P.L. 4-Methylumbelliferone treatment and hyaluronan inhibition as a therapeutic strategy in inflammation, autoimmunity, and cancer. Front. Immunol. 2015, 6, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Kakizaki, I.; Kojima, K.; Takagaki, K.; Endo, M.; Kannagi, R.; Ito, M.; Maruo, Y.; Sato, H.; Yasuda, T.; Mita, S.; et al. A novel mechanism for the inhibition of hyaluronan biosynthesis by 4-methylumbelliferone. J. Biol. Chem. 2004, 279, 33281–33289. [Google Scholar] [CrossRef] [PubMed]
- Braun, S.; Botzki, A. Design of benzimidazole- and benzoxazole-2-thione derivatives as inhibitors of bacterial hyaluronan. Eur. J. Med. Chem. 2011, 46, 4419–4429. [Google Scholar] [CrossRef] [PubMed]
- Hynes, W.L.; Walton, S.L. Hyaluronidases of Gram-positive bacteria. FEMS Microbiol. Lett. 2000, 183, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Burdick, J.A.; Prestwich, G.D. Hyaluronic Acid Hydrogels for Biomedical Applications. Adv. Mater. 2011, 23, H41–H56. [Google Scholar] [CrossRef] [PubMed]
- Prestwich, G.D.; Kuo, J.W. Chemically-modified HA for therapy and regenerative medicine. Curr. Pharm. Biotechnol. 2008, 9, 242–245. [Google Scholar] [CrossRef] [PubMed]
- Montanari, E.; Capece, S. Hyaluronic Acid Nanohydrogels as a Useful Tool for BSAO Immobilization in the Treatment of Melanoma Cancer Cells. Macromol. Biosci. 2013, 13, 1185–1194. [Google Scholar] [CrossRef] [PubMed]
- Di Meo, C.; Montanari, E. Highly versatile nanohydrogel platform based on riboflavin-polysaccharide derivatives useful in the development of intrinsically fluorescent and cytocompatible drug carriers. Carbohydr. Polym. 2015, 115, 502–509. [Google Scholar] [CrossRef] [PubMed]
- Choi, K.Y.; Min, K.H. Self-assembled hyaluronic acid nanoparticles as a potential drug carrier for cancer therapy: Synthesis, characterization, and in vivo biodistribution. J. Mater. Chem. 2009, 19, 4102–4107. [Google Scholar] [CrossRef]
- Pitarresi, G.; Craparo, E.F. Composite Nanoparticles Based on Hyaluronic Acid Chemically Cross-Linked with α,β-Polyaspartylhydrazide. Biomacromolecules 2007, 8, 1890–1898. [Google Scholar] [CrossRef] [PubMed]
- Jha, A.K.; Hule, R.A. Structural Analysis and Mechanical Characterization of Hyaluronic Acid-Based Doubly Cross-Linked Networks. Macromolecules 2009, 42, 537–546. [Google Scholar] [CrossRef] [PubMed]
- Park, J.H.; Cho, H.J. Hyaluronic acid derivative-coated nanohybrid liposomes for cancer imaging and drug delivery. J. Control. Release 2014, 174, 98–108. [Google Scholar] [CrossRef] [PubMed]
- Mizrahy, S.; Raz, S.R. Hyaluronan-coated nanoparticles: The influence of the molecular weight on CD44-hyaluronan interactions and on the immune response. J. Control. Release 2011, 156, 231–238. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; He, Y. Hyaluronic acid-modified hydrothermally synthesized iron oxide nanoparticles for targeted tumor MR imaging. Biomaterials 2014, 35, 3666–3677. [Google Scholar] [CrossRef] [PubMed]
- Chudobova, D.; Nejdl, L. Complexes of Silver(I) Ions and Silver Phosphate Nanoparticles with Hyaluronic Acid and/or Chitosan as Promising Antimicrobial Agents for Vascular Grafts. Int. J. Mol. Sci. 2013, 14, 13592–13614. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Miller, R.J. Hyaluronic Acid-Based Drug Conjugates: State-of-the-Art and Perspectives. J. Biomed. Nanotechnol. 2014, 10, 4–16. [Google Scholar] [CrossRef] [PubMed]
- Gennari, A.; Gujral, C. Revisiting Boronate/Diol Complexation as a Double Stimulus-Responsive Bioconjugation. Bioconjug. Chem. 2017, 28, 1391–1402. [Google Scholar] [CrossRef] [PubMed]
- Arpicco, S.; Milla, P. Hyaluronic Acid Conjugates as Vectors for the Active Targeting of Drugs, Genes and Nanocomposites in Cancer Treatment. Molecules 2014, 19, 3193–3230. [Google Scholar] [CrossRef] [PubMed]
- Mero, A.; Campisi, M. Hyaluronic Acid as a Protein Polymeric Carrier: An Overview and a Report on Human Growth Hormone. Curr. Drug Targets 2015, 16, 1503–1511. [Google Scholar] [CrossRef] [PubMed]
- Dreaden, E.C.; Morton, S.W. Bimodal Tumor-Targeting from Microenvironment Responsive Hyaluronan Layer-by-Layer (LbL) Nanoparticles. ACS Nano 2014, 8, 8374–8382. [Google Scholar] [CrossRef] [PubMed]
- Montanari, E.; Gennari, A. Hyaluronan/Tannic Acid Nanoparticles via Catechol/Boronate Complexation as a Smart Antibacterial System. Macromol. Biosci. 2016, 16, 1815–1823. [Google Scholar] [CrossRef] [PubMed]
- De la Fuente, M.; Seijo, B.A.; Alonso, M.J. Novel Hyaluronic Acid-Chitosan Nanoparticles for Ocular Gene Therapy. Investig. Ophthalmol. Vis. Sci. 2008, 49, 2016–2024. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Ahn, C.H.; Park, T.G. Poly [lactic-co-(glycolic acid)]-grafted hyaluronic acid copolymer micelle nanoparticles for target-specific delivery of doxorubicin. Macromol. Biosci. 2009, 9, 336–342. [Google Scholar] [CrossRef] [PubMed]
- Yadav, A.K.; Mishra, P.; Jain, S. Preparation and characterization of HA-PEG-PCL intelligent core-corona nanoparticles for delivery of doxorubicin. J. Drug Target 2008, 16, 464–478. [Google Scholar] [CrossRef] [PubMed]
- Montanari, E.; Di Meo, C.; Sennato, S.; Francioso, A.; Marinelli, A.L.; Ranzo, F.; Schippa, S.; Coviello, T.; Bordi, F.; Matricardi, P. Hyaluronan-cholesterol nanohydrogels: Characterisation and effectiveness in carrying alginate lyase. New Biotechnol. 2017, 37, 80–89. [Google Scholar] [CrossRef] [PubMed]
- Ganesh, S.; Iyer, A.K.; Morrissey, D.V.; Amiji, M.M. Hyaluronic acid based self-assembling nanosystems for CD44 target mediated siRNA delivery to solid tumours. Biomaterials 2013, 34, 3489–3502. [Google Scholar] [CrossRef] [PubMed]
- Craparo, G.F.; Cavallaro, G.; Bondì, M.L.; Giammona, G. Preparation of polymeric nanoparticles by photo-crosslinking of an acryloylated polyaspartamide in w/o microemulsion. Macromol. Chem. Phys. 2004, 205, 1955–1964. [Google Scholar] [CrossRef]
- Shu, X.Z.; Liu, Y.; Luo, Y.; Roberts, M.C.; Prestwich, G.D. Disulfide cross-linked hyaluronan hydrogels. Biomacromolecules 2002, 3, 1304–1311. [Google Scholar] [CrossRef] [PubMed]
- Oishi, M.; Nagasaki, Y. Stimuli-responsive smart nanogels for cancer diagnostics and therapy. Nanomedicine 2010, 5, 451–468. [Google Scholar] [CrossRef] [PubMed]
- Ossipov, D.A. Nanostructured hyaluronic acid-based materials for active delivery to cancer. Exp. Opin. Drug Deliv. 2010, 7, 681–703. [Google Scholar] [CrossRef] [PubMed]
- Rios de la Rosa, J.M.; Tirella, A. The CD44-Mediated Uptake of Hyaluronic Acid-Based Carriers in Macrophages. Adv. Healthc. Mater. 2017, 6. [Google Scholar] [CrossRef] [PubMed]
- Silva, J.P.; Gonçalves, C. Delivery of LLKKK18 loaded into self-assembling hyaluronic acid nanogel for tuberculosis treatment. J. Control. Release 2016, 235, 112–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lallana, E.; Rios de la Rosa, J.M.; Tirella, A.; Pelliccia, M.; Gennari, A.; Stratford, I.J.; Puri, S.; Ashford, M.; Tirelli, N. Chitosan/Hyaluronic Acid Nanoparticles: Rational Design Revisited for RNA Delivery. Mol. Pharmaceut. 2017, 14, 2422–2436. [Google Scholar] [CrossRef] [PubMed]
- Choi, K.Y.; Min, K.H.; Yoon, H.Y.; Kim, K.; Park, J.H.; Kwon, I.C.; Choi, K.; Jeong, S.Y. PEGylation of hyaluronic acid nanoparticles improves tumor targetability in vivo. Biomaterials 2011, 32, 1880–1889. [Google Scholar] [CrossRef] [PubMed]
- Almalik, A.; Day, P.J.; Tirelli, N. HA-Coated Chitosan Nanoparticles for CD44-Mediated Nucleic Acid Delivery. Mol Pharm. 2013, 13, 1671–1680. [Google Scholar] [CrossRef] [PubMed]
- Pieters, J. Entry and survival of pathogenic mycobacteriain macrophages. Microbes Infect. 2001, 3, 249–255. [Google Scholar] [CrossRef]
- Richter-Dahlfors, A.; Buchan, A.M.J. Murine Salmonellosis Studied by Confocal Microscopy: Salmonella typhimurium Resides Intracellularly inside Macrophages and Exerts a Cytotoxic Effect on Phagocytes in vivo. J. Exp. Med. 1997, 186, 569–580. [Google Scholar] [CrossRef] [PubMed]
- Köhler, S.; Porte, F. The intramacrophagic environment of Brucella suis and bacterial response. Vet. Microbiol. 2002, 90, 299–309. [Google Scholar] [CrossRef]
- Suzuki, T.; Sasakawa, C. Molecular Basis of the Intracellular Spreading of Shigella. Infect. Immun. 2001, 69, 5959–5966. [Google Scholar] [CrossRef] [PubMed]
- Portnoy, D.A.; Auerbuch, V. The cell biology of Listeria monocytogenes infection. The intersection of bacterial pathogenesis and cell-mediated immunity. J. Cell Biol. 2002, 158, 409–414. [Google Scholar] [CrossRef] [PubMed]
- Van Kirk, L.S.; Hayes, S.F. Ultrastructure of Rickettsia rickettsii Actin Tails and Localization of Cytoskeletal Proteins. Infect. Immun. 2000, 68, 4706–4713. [Google Scholar] [CrossRef] [PubMed]
- Flannagan, R.S.; Heit, B. Intracellular replication of Staphylococcus aureus in mature phagolysosomes in macrophages precedes host cell death, and bacterial escape and dissemination. Cell. Microbiol. 2016, 18, 514–535. [Google Scholar] [CrossRef] [PubMed]
- Haggar, A.; Hussain, M. Extracellular Adherence Protein from Staphylococcus aureus Enhances Internalization into Eukaryotic Cells. Infect. Immun. 2003, 71, 2310–2317. [Google Scholar] [CrossRef] [PubMed]
- Van Bambeke, F. Cellular pharmacodynamics and pharmacokinetics of antibiotics: Current views and perspectives. Curr. Opin. Drug Discov. Dev. 2006, 9, 218–230. [Google Scholar]
- Carryn, S.; Chanteux, H. Intracellular pharmacodynamics of antibiotics. Infect. Dis. Clin. N. Am. 2003, 17, 615–634. [Google Scholar] [CrossRef]
- Maurin, M.; Raoult, D. Use of Aminoglycosides in Treatment of Infections Due to Intracellular Bacteria. Antimicrob. Agents Chemother. 2001, 45, 2977–2986. [Google Scholar] [CrossRef] [PubMed]
- Falzari, K.; Zhu, Z.; Pan, D.; Liu, H.; Hongmanee, P.; Franzblau, S.G. In Vitro and In Vivo Activities of Macrolide Derivatives against Mycobacterium tuberculosis. Antimicrob. Agents Chemother. 2005, 49, 1447–1454. [Google Scholar] [CrossRef] [PubMed]
- Van Bambeke, F.; Michot, J.M. Antibiotic efflux pumps in eukaryotic cells: Occurrence and impact on antibiotic cellular pharmacokinetics, pharmacodynamics and toxicodynamics. J. Antimicrob. Chemother. 2003, 51, 1067–1077. [Google Scholar] [CrossRef] [PubMed]
- Montanari, E.; Oates, A.; Di Meo, C.; Meade, J.; Cerrone, R.; Francioso, A.; Devine, D.; Coviello, T.; Mancini, P.; Mosca, L.; et al. Hyaluronan-Based Nanohydrogels for Targeting Intracellular S. aureus in Human Keratinocytes. Adv. Healthc. Mater. 2018, accepted. [Google Scholar]
- Zaki, N.M.; Nasti, A. Nanocarriers for Cytoplasmic Delivery: Cellular Uptake and Intracellular Fate of Chitosan and Hyaluronic Acid-Coated Chitosan Nanoparticles in a Phagocytic Cell Model. Macromol. Biosci. 2011, 11, 1747–1760. [Google Scholar] [CrossRef] [PubMed]
- Platt, V.M.; Szoka, F.C. Jr. Anticancer therapeutics: Targeting macromolecules and nanocarriers to hyaluronan or CD44, a hyaluronan receptor. Mol. Pharm. 2008, 5, 474–486. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Benjaminsen, R.V. Hyaluronic Acid Immobilized Polyacrylamide Nanoparticle Sensors for CD44 Receptor Targeting and pH Measurement in Cells. Bioconjug. Chem. 2012, 23, 2247–2255. [Google Scholar] [CrossRef] [PubMed]
- Palvai, S.; Kuman, M.M. Hyaluronic acid cloaked oleic acid nanoparticles inhibit MAPK signaling with sub-cellular DNA damage in colon cancer cells. J. Mater. Chem. B 2017, 5, 3658–3666. [Google Scholar] [CrossRef]
- Qhattal, H.S.S.; Liu, X. Characterization of CD44-Mediated Cancer Cell Uptake and Intracellular Distribution of Hyaluronan-Grafted Liposomes. Mol. Pharm. 2011, 8, 1233–1246. [Google Scholar] [CrossRef] [PubMed]
- Rajendran, L.; Knölker, H.J. Subcellular targeting strategies for drug design and delivery. Nat. Rev. Drug Discov. 2010, 9, 29–42. [Google Scholar] [CrossRef] [PubMed]
- Yameen, B.; Choi, W.I. Insight into nanoparticle cellular uptake and intracellular targeting. J. Control. Release 2014, 190, 485–499. [Google Scholar] [CrossRef] [PubMed]
- Vasir, J.K.; Labhasetwar, V. Biodegradable nanoparticles for cytosolic delivery of therapeutics. Adv. Drug Deliv. Rev. 2007, 59, 718–728. [Google Scholar] [CrossRef] [PubMed]
- Behr, J.P. The Proton Sponge: A Trick to Enter Cells the Viruses Did Not Exploit. Chim. Int. J. Chem. 1997, 51, 34–36. [Google Scholar]
- Guo, S.; Huang, L. Nanoparticles escaping RES and endosome: Challenges for siRNA delivery for cancer therapy. J. Nanomater. 2011, 2011, 1–12. [Google Scholar] [CrossRef]
- Contreras-Ruiz, L.; De la Fuente, M. Intracellular trafficking of hyaluronic acid-chitosan oligomer-based nanoparticles in cultured human ocular surface cells. Mol. Vis. 2011, 17, 279–290. [Google Scholar] [PubMed]
- An, T.; Zhang, C. Hyaluronic acid-coated poly([small beta]-amino) ester nanoparticles as carrier of doxorubicin for overcoming drug resistance in breast cancer cells. RSC Adv. 2016, 6, 38624–38636. [Google Scholar] [CrossRef]
- Qiu, L.; Li, Z. Self-assembled pH-responsive hyaluronic acid–g-poly(l-histidine) copolymer micelles for targeted intracellular delivery of doxorubicin. Acta Biomater. 2014, 10, 2024–2035. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.S.; Na, K. Photochemically Triggered Cytosolic Drug Delivery Using pH-Responsive Hyaluronic Acid Nanoparticles for Light-Induced Cancer Therapy. Biomacromolecules 2014, 15, 4228–4238. [Google Scholar] [CrossRef] [PubMed]
- Wagner, E. Application of membrane-active peptides for nonviral gene delivery. Adv. Drug Deliv. Rev. 1999, 38, 279–289. [Google Scholar] [CrossRef]
- Cullis, P.R.; Hope, M.J. Lipid polymorphism and the roles of lipids in membranes. Chem. Phys. Lipids 1986, 40, 127–144. [Google Scholar] [CrossRef]
- Hafez, I.M.; Maurer, N. On the mechanism whereby cationic lipids promote intracellular delivery of polynucleic acids. Gene Ther. 2001, 8, 1188–1196. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.; Hou, Y. Hyaluronic acid conjugation facilitates clearance of intracellular bacterial infections by streptomycin with neglectable nephrotoxicity. Glycobiology 2017, 27, 861–867. [Google Scholar] [CrossRef] [PubMed]
- Mu, H.; Niu, H. Chitosan conjugation enables intracellular bacteria susceptible to aminoglycoside antibiotic. Glycobiology 2016, 26, 1190–1197. [Google Scholar] [CrossRef] [PubMed]
- Montanari, E.; D’Arrigo, G. Chasing bacteria within the cells using levofloxacin-loaded hyaluronic acid nanohydrogels. Eur. J. Pharm. Biopharm. 2014, 87, 518–523. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Qiu, Y. Synthesis of hyaluronan-amikacin conjugate and its bactericidal activity against intracellular bacteria in vitro and in vivo. Carbohydr. Polym. 2018, 181, 132–140. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
| Pathogen | Cell Line | CD44 Role | HA Role |
|---|---|---|---|
| S. pyogenes [4] | Human keratinocytes | CD44 represents the main receptor for cell attachment. | HA-based capsules are synthesised for promoting the cell invasion. |
| S. pyogenes [61] | Murine epithelial keratinocytes | CD44 is found to be widely expressed in the site of infection, acting as a major cellular receptor for the cellular entry. | HA-based capsules are synthesised for promoting the cell invasion. |
| M. tuberculosis [5] | Human lung epithelial cells | Employment of extracellular DNA-binding proteins to attach host cells through HA. | |
| M. tuberculosis [7] | Murine macrophages | CD44 involvement in the binding and subsequent cellular internalisation. | |
| S. aureus [8] | Human neutrophils | CD44 influences the pathogen phagocytosis through its structural and functional linkage to the cytoskeletal microfilaments. | |
| Shigella spp. [62] | Human epithelial cells | The IpaB-CD44 interaction leads to the transduction of signals that participate in the cytoskeletal rearrangements and the subsequent internalisation of the pathogen within the cells. | |
| Listeria spp. [9] | Murine macrophages and fibroblasts | CD44 facilitates the intracellular growth of the pathogen intracellularly. | |
| M. tuberculosis [63] | Human lung epithelial cells | Short HA chains are utilised as a carbon source for proliferation. | |
| M. bovis bacillus Calmette-Guerin [63] | Human lung epithelial cells | Short HA chains are utilised as a carbon source for proliferation. | |
| Leishmania [6] | Murine macrophages | HA acts as endogenous essential nutrient for the growth and virulence. |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Montanari, E.; Di Meo, C.; Oates, A.; Coviello, T.; Matricardi, P. Pursuing Intracellular Pathogens with Hyaluronan. From a ‘Pro-Infection’ Polymer to a Biomaterial for ‘Trojan Horse’ Systems. Molecules 2018, 23, 939. https://doi.org/10.3390/molecules23040939
Montanari E, Di Meo C, Oates A, Coviello T, Matricardi P. Pursuing Intracellular Pathogens with Hyaluronan. From a ‘Pro-Infection’ Polymer to a Biomaterial for ‘Trojan Horse’ Systems. Molecules. 2018; 23(4):939. https://doi.org/10.3390/molecules23040939
Chicago/Turabian StyleMontanari, Elita, Chiara Di Meo, Angela Oates, Tommasina Coviello, and Pietro Matricardi. 2018. "Pursuing Intracellular Pathogens with Hyaluronan. From a ‘Pro-Infection’ Polymer to a Biomaterial for ‘Trojan Horse’ Systems" Molecules 23, no. 4: 939. https://doi.org/10.3390/molecules23040939