Modification and Assembly of a Versatile Lactonase for Bacterial Quorum Quenching

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
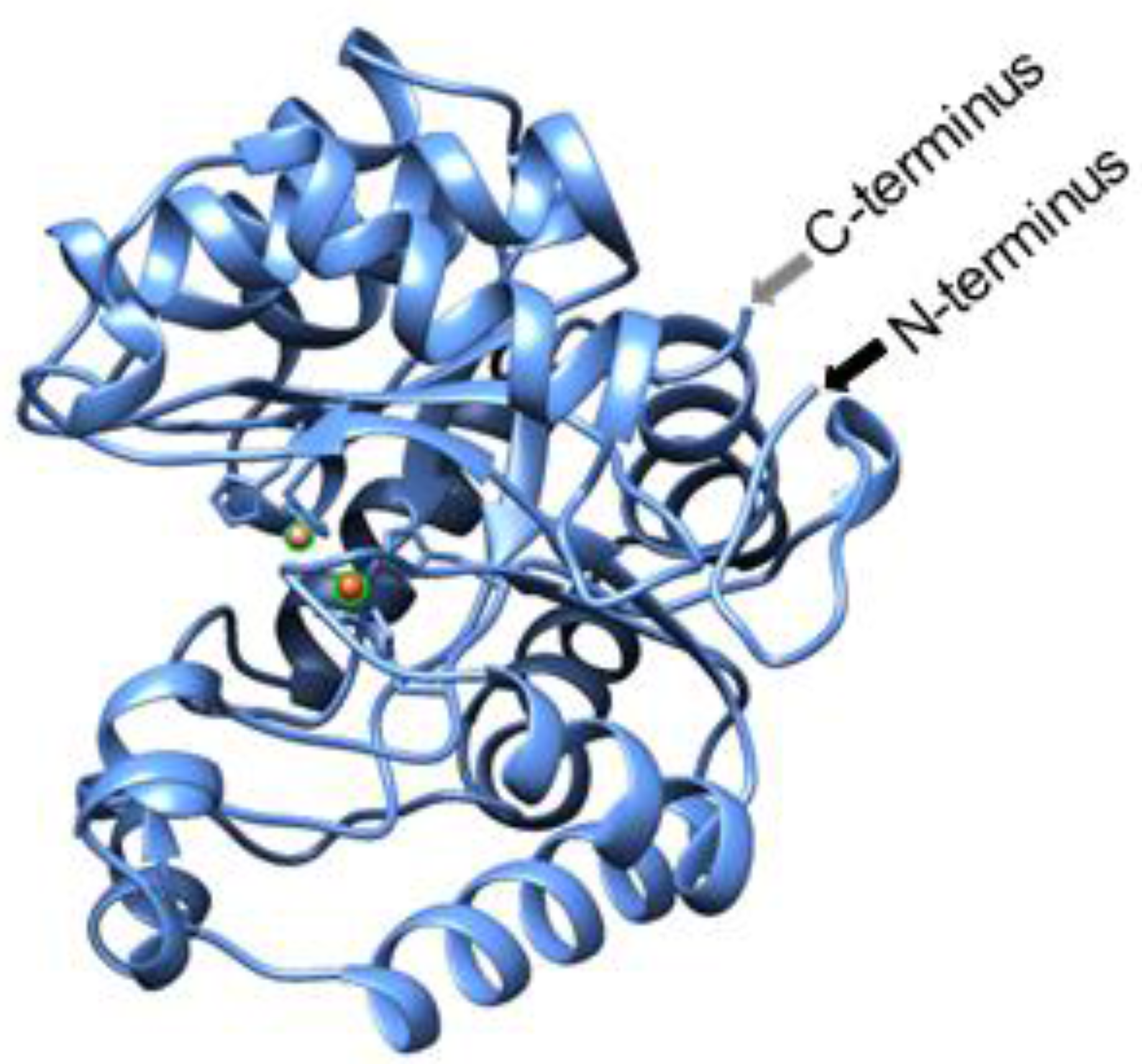
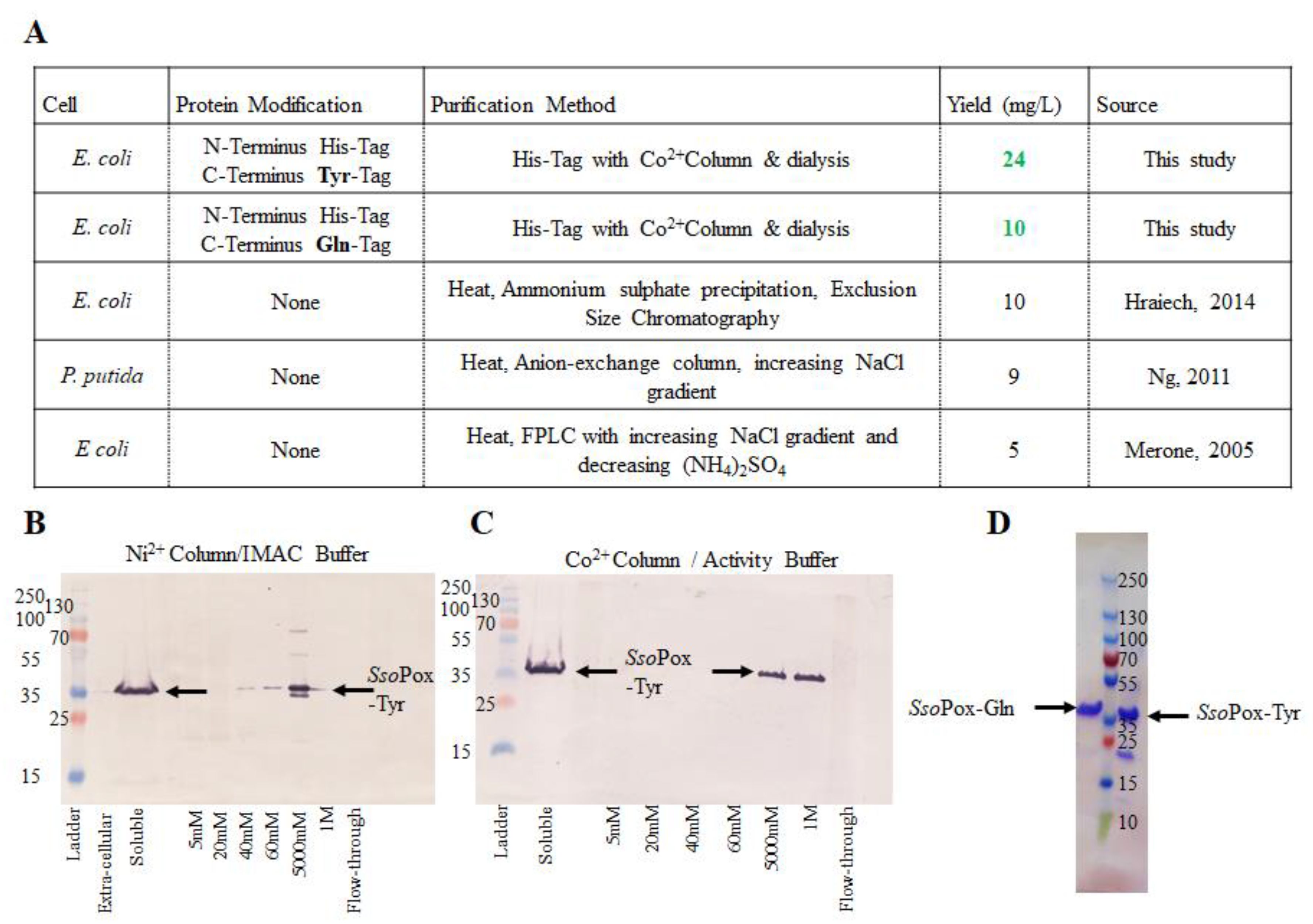
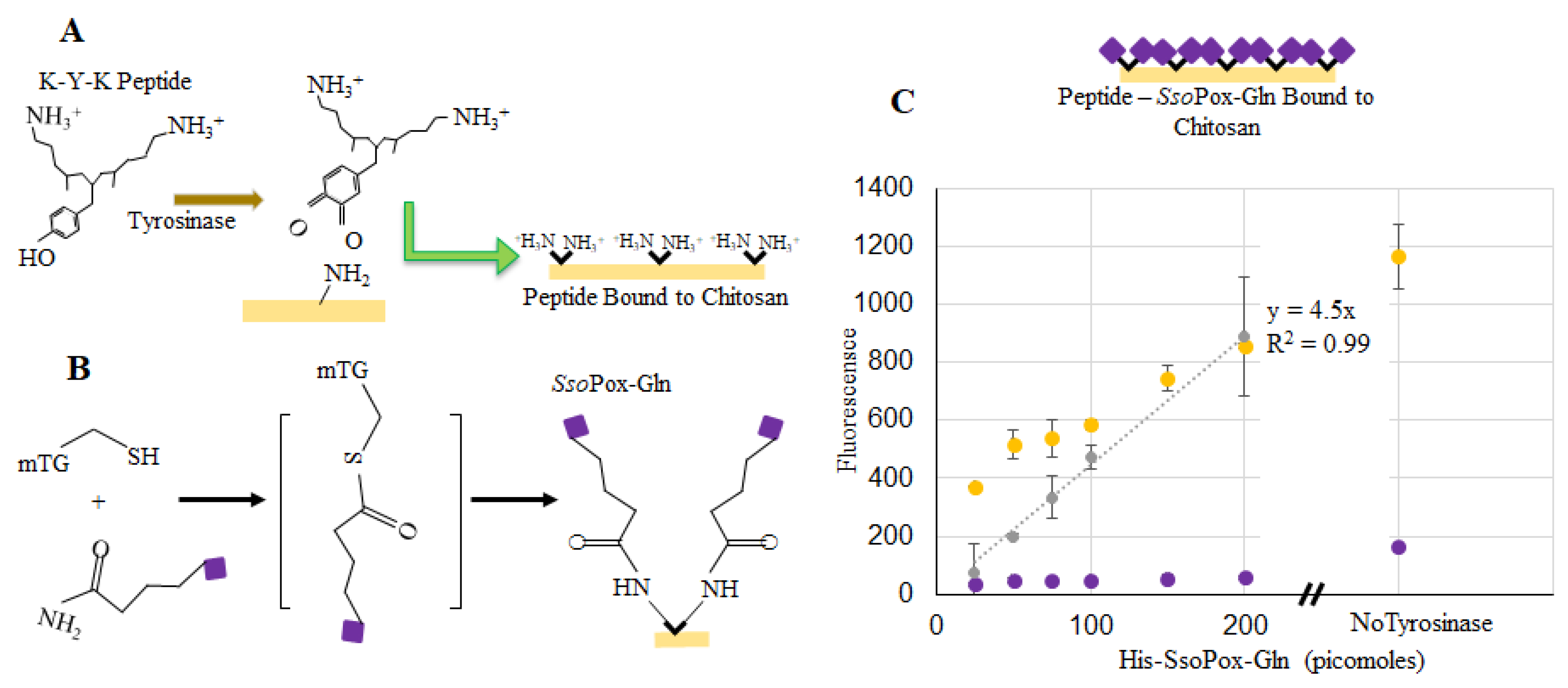
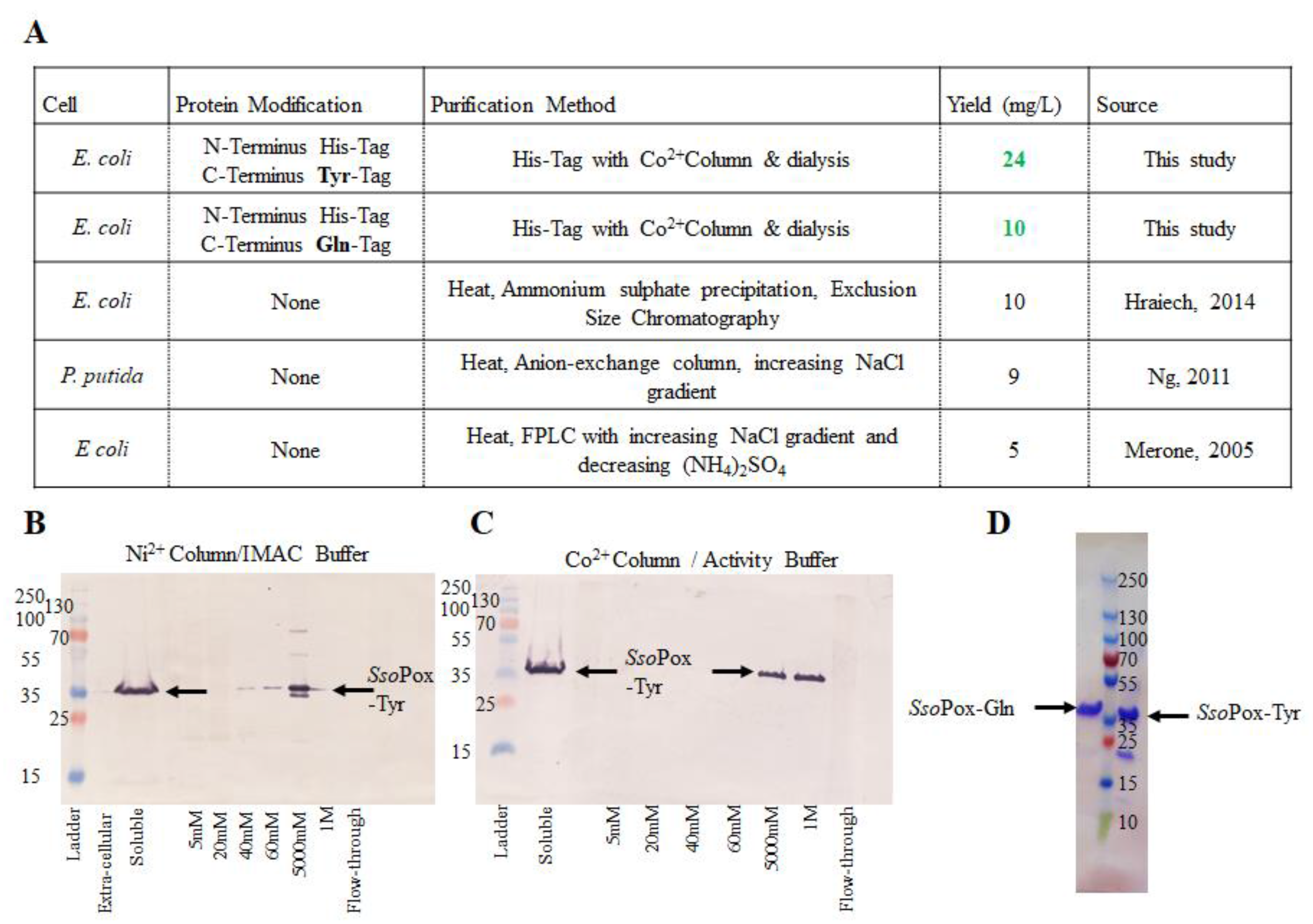
2.1. Purification of Modified SsoPox
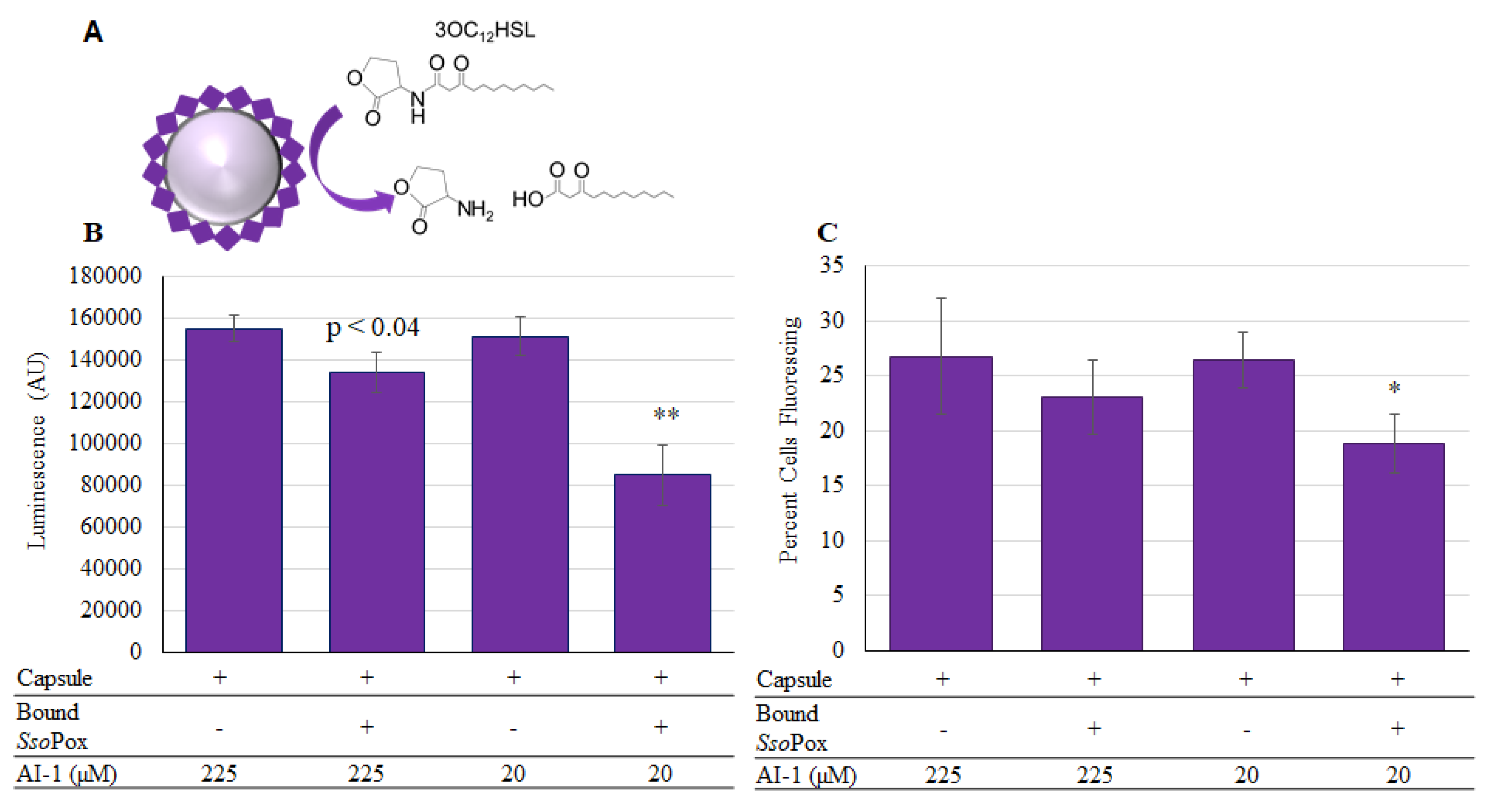
2.2. AI-1 Reporters
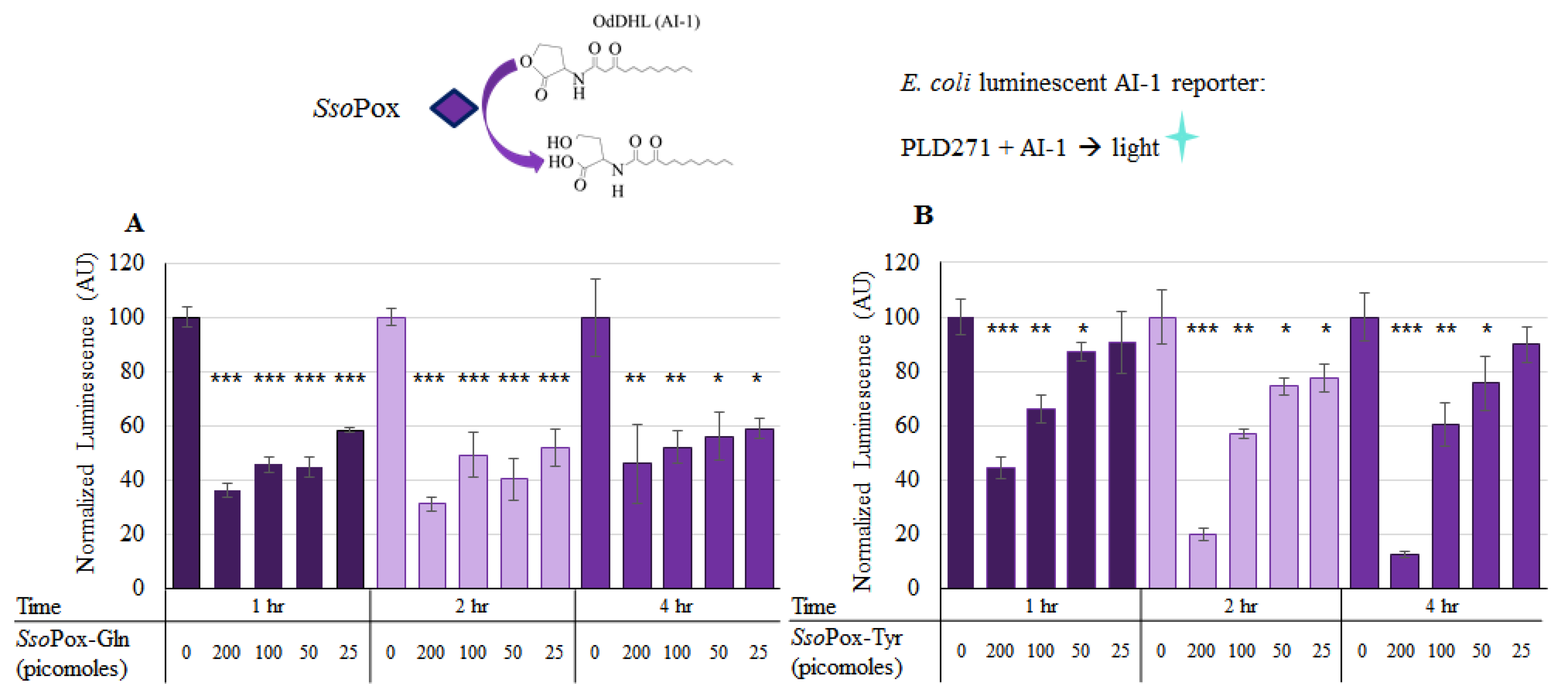
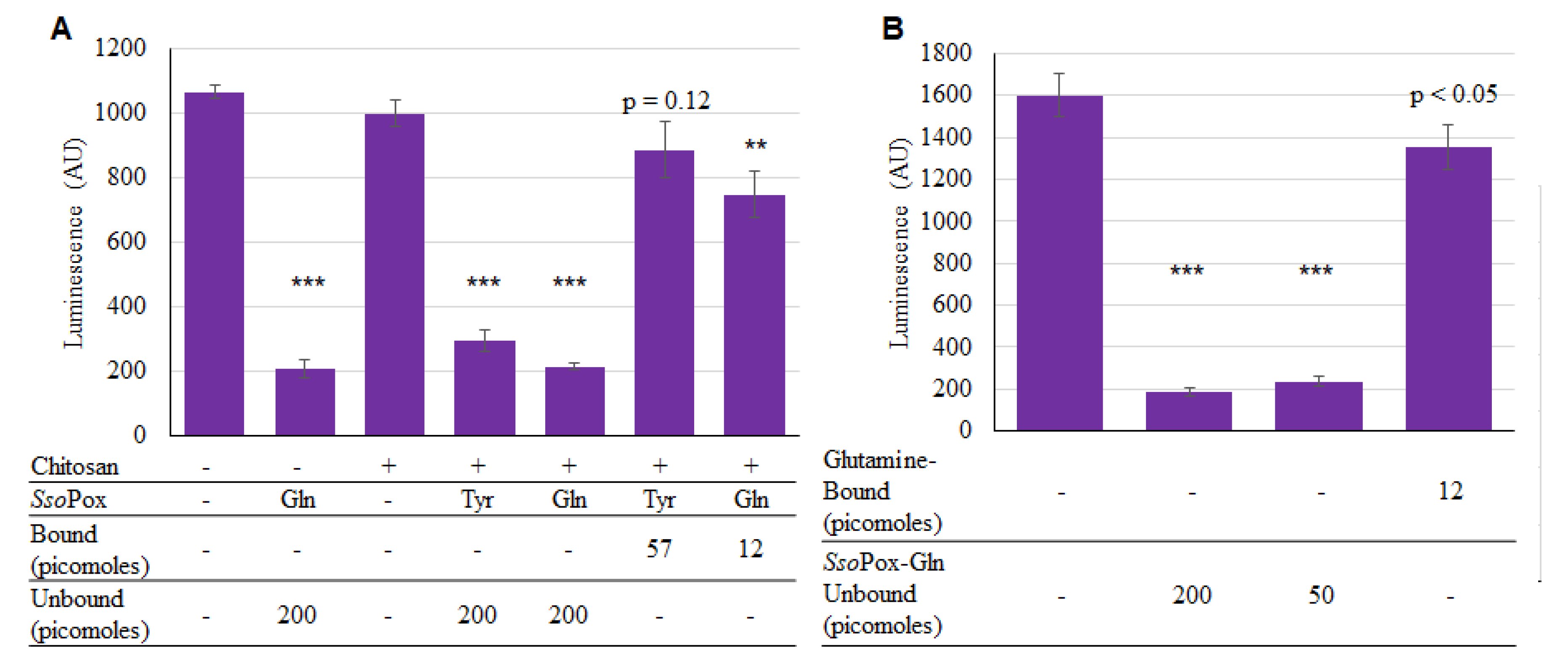
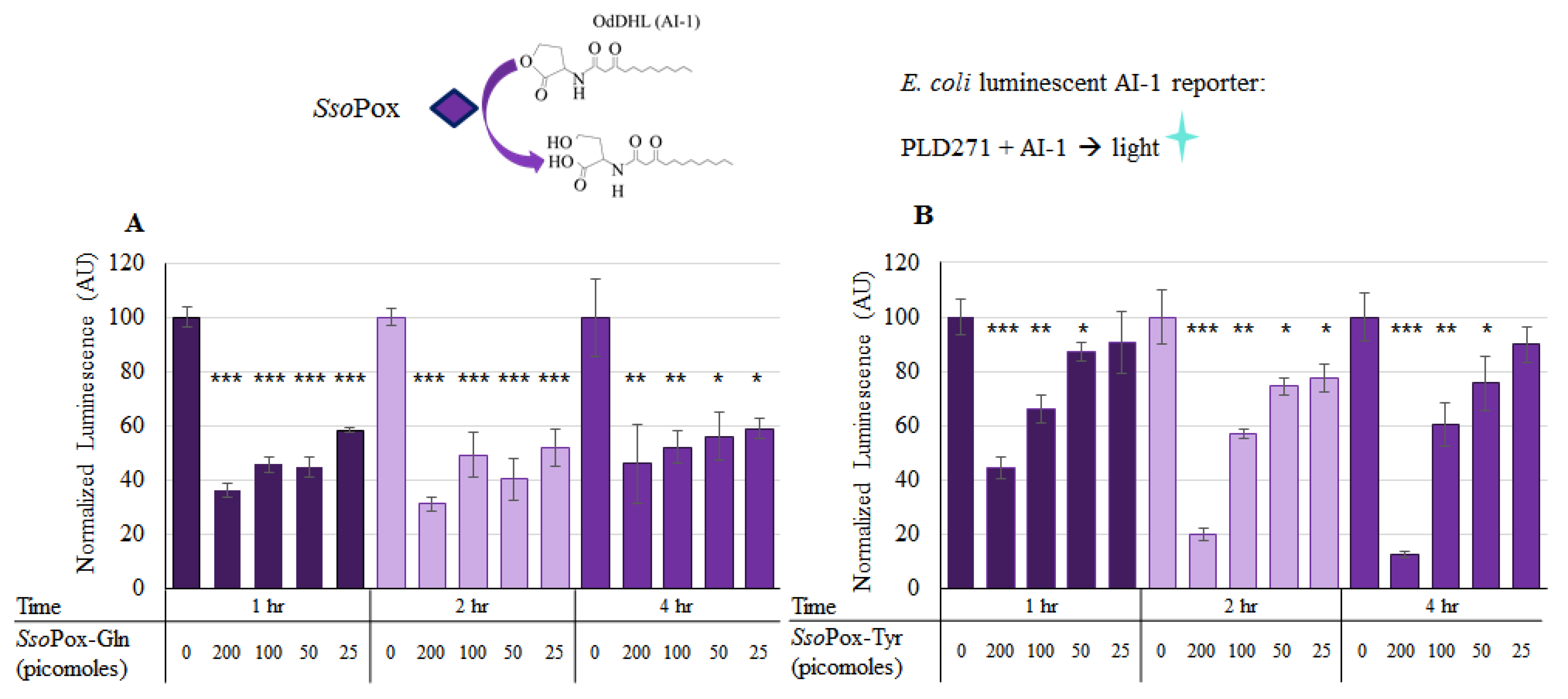
2.3. SsoPox Quorum Quenching
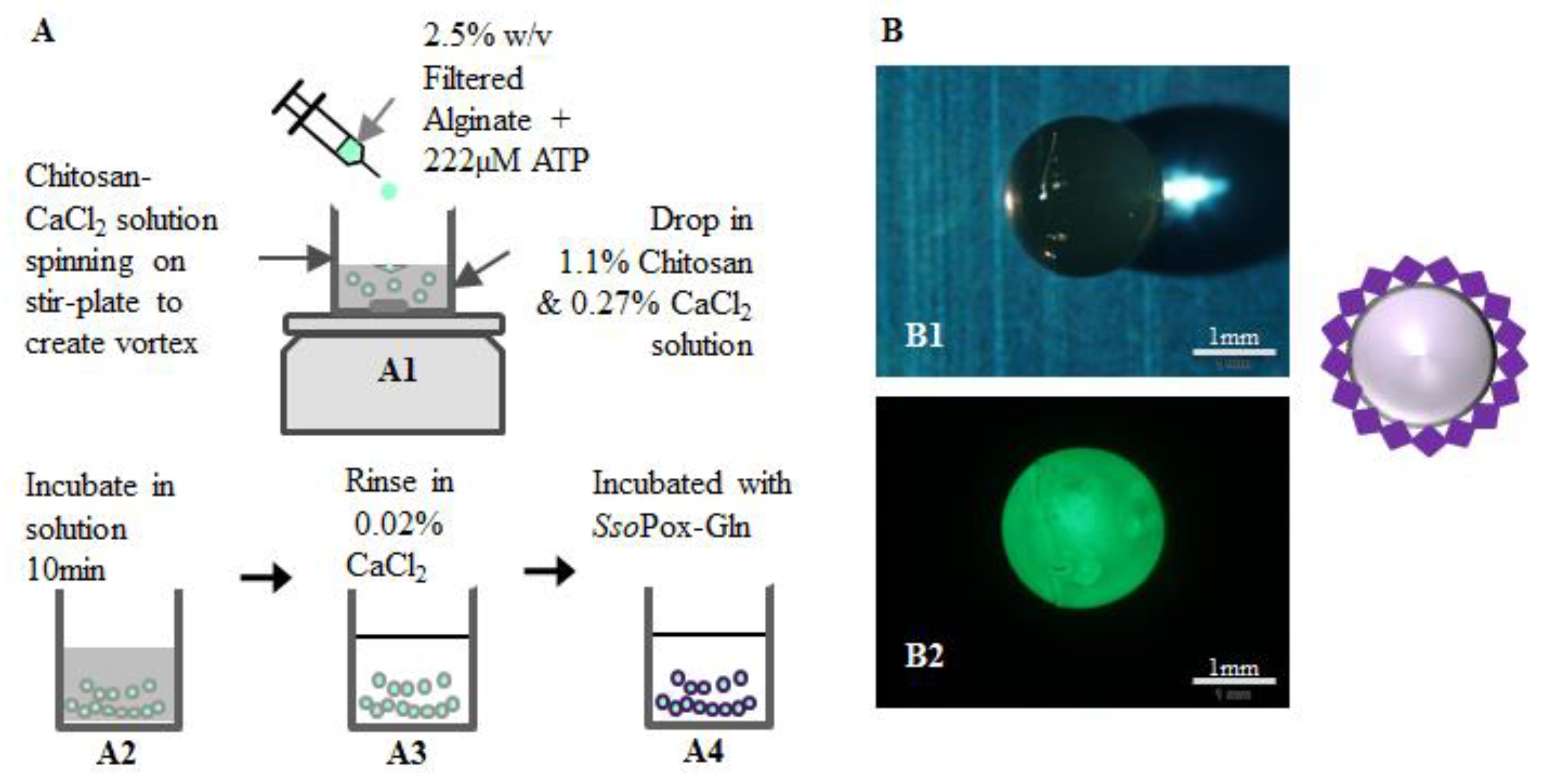
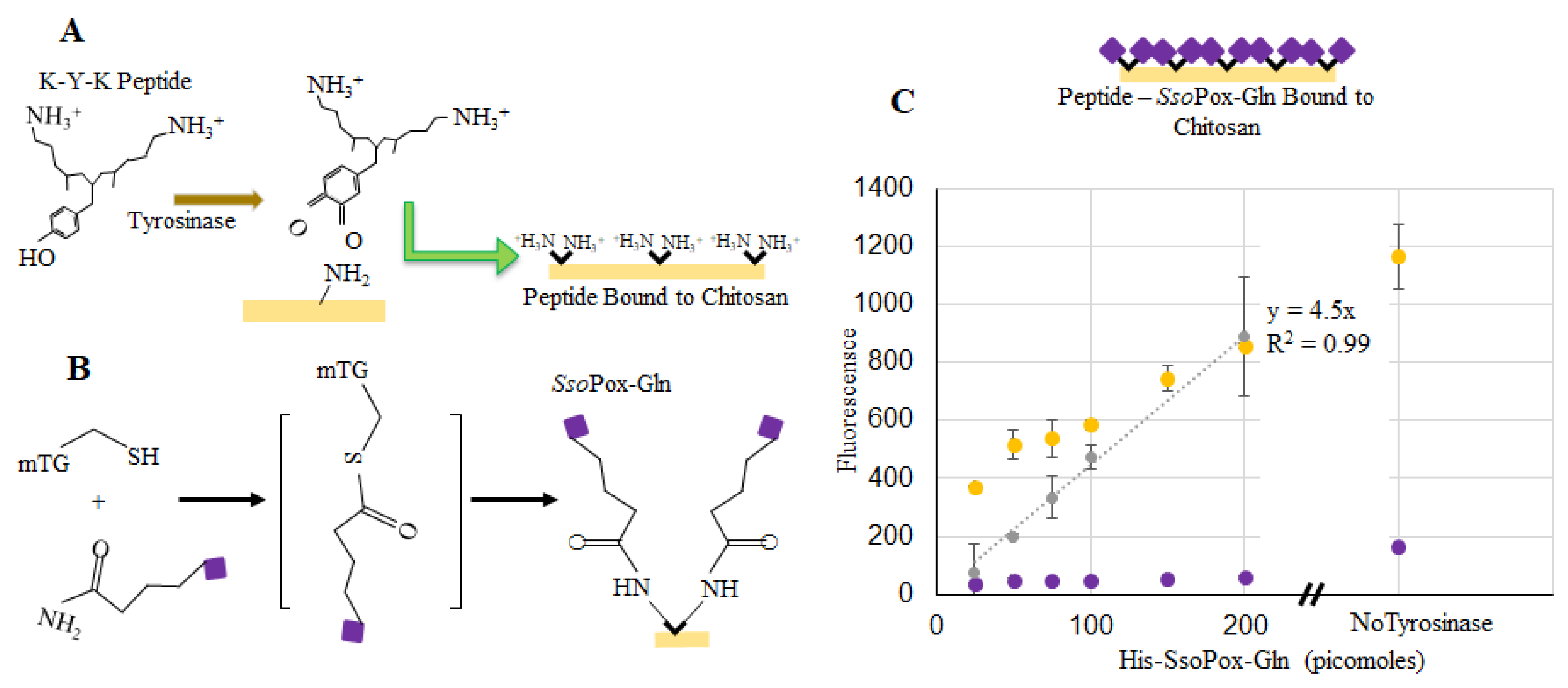
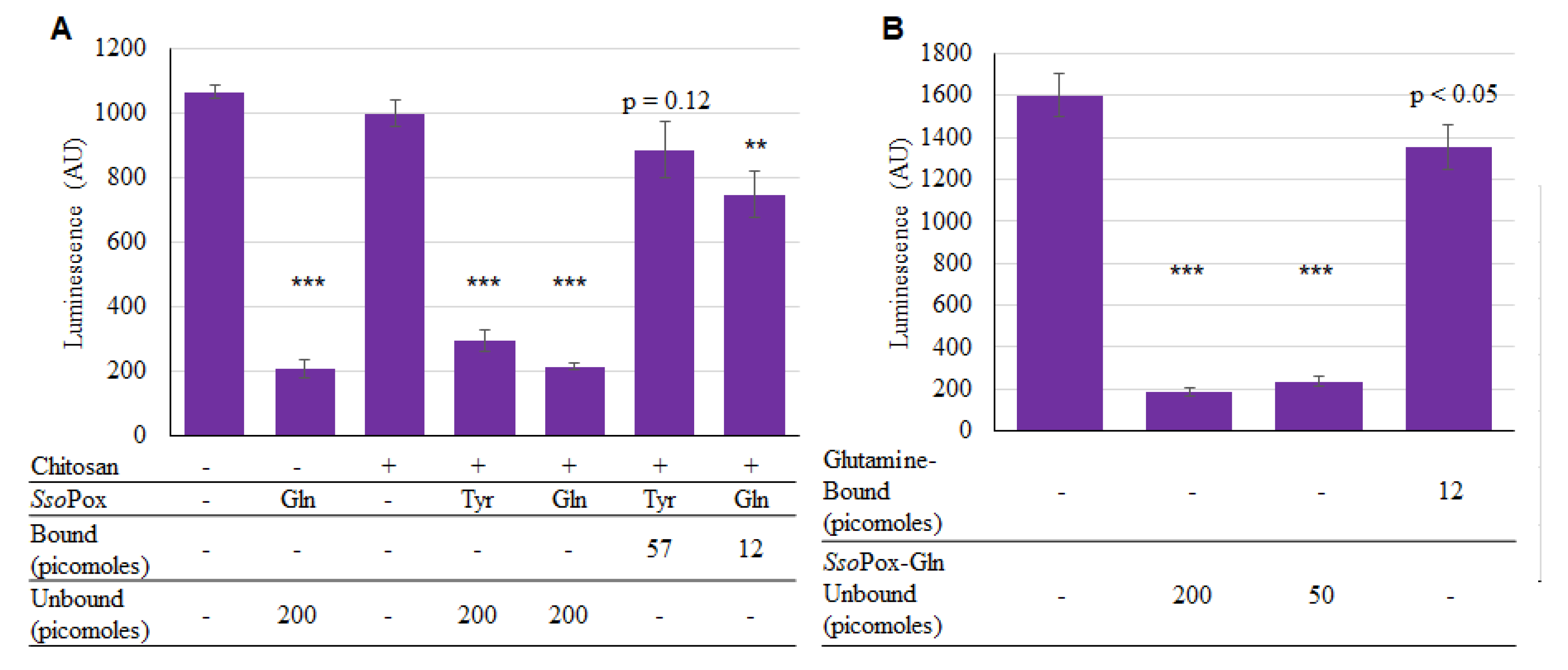
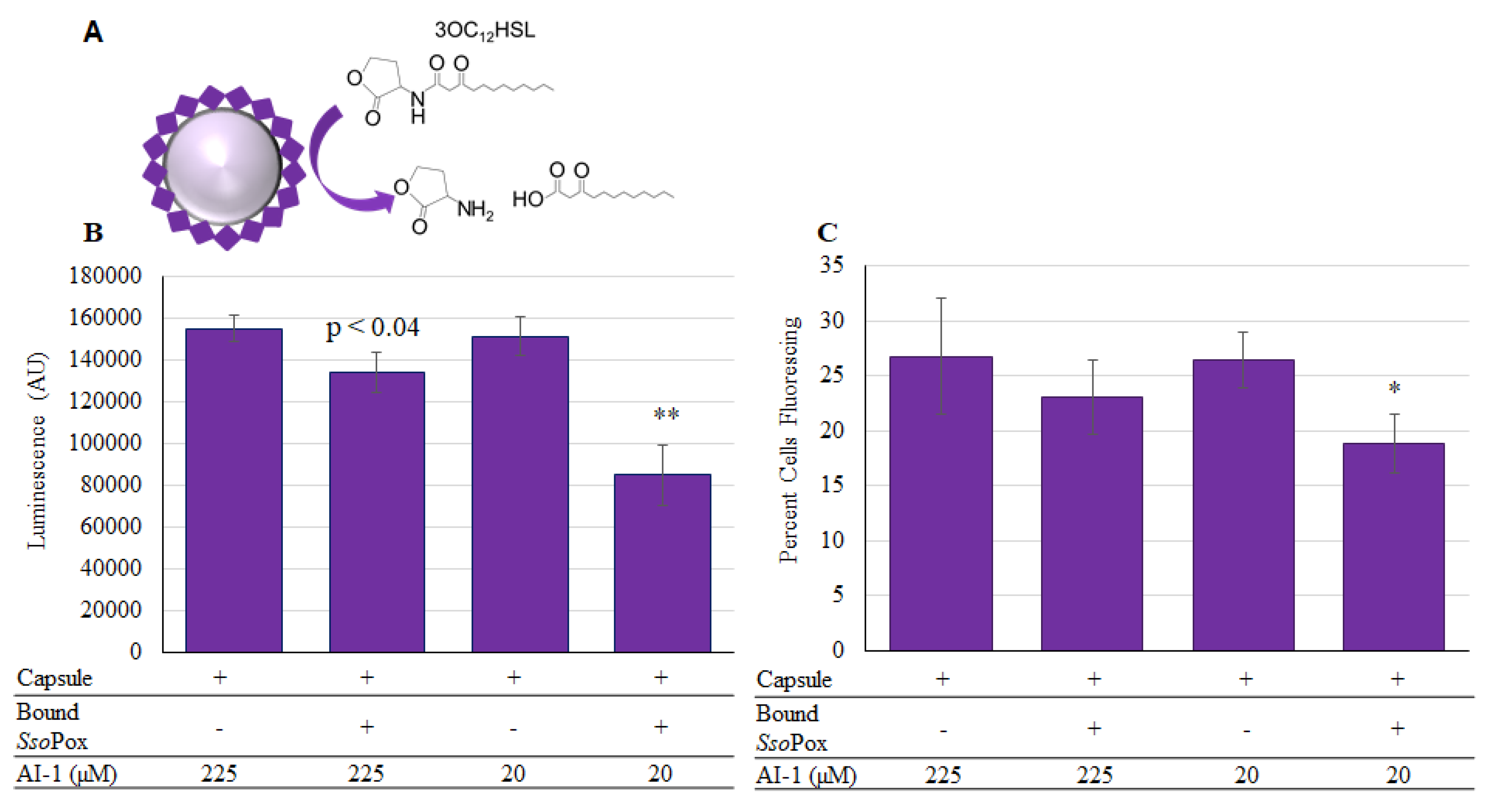
2.4. SsoPox-Gln Capsules for Quorum Quenching
3. Discussion
4. Materials and Methods
4.1. SsoPox Expression Plasmids
4.2. Overexpression and Purification of Modified SsoPox
4.3. AI-1 Reporter Construction
4.4. Determining Lactonase Activity of Modified SsoPox
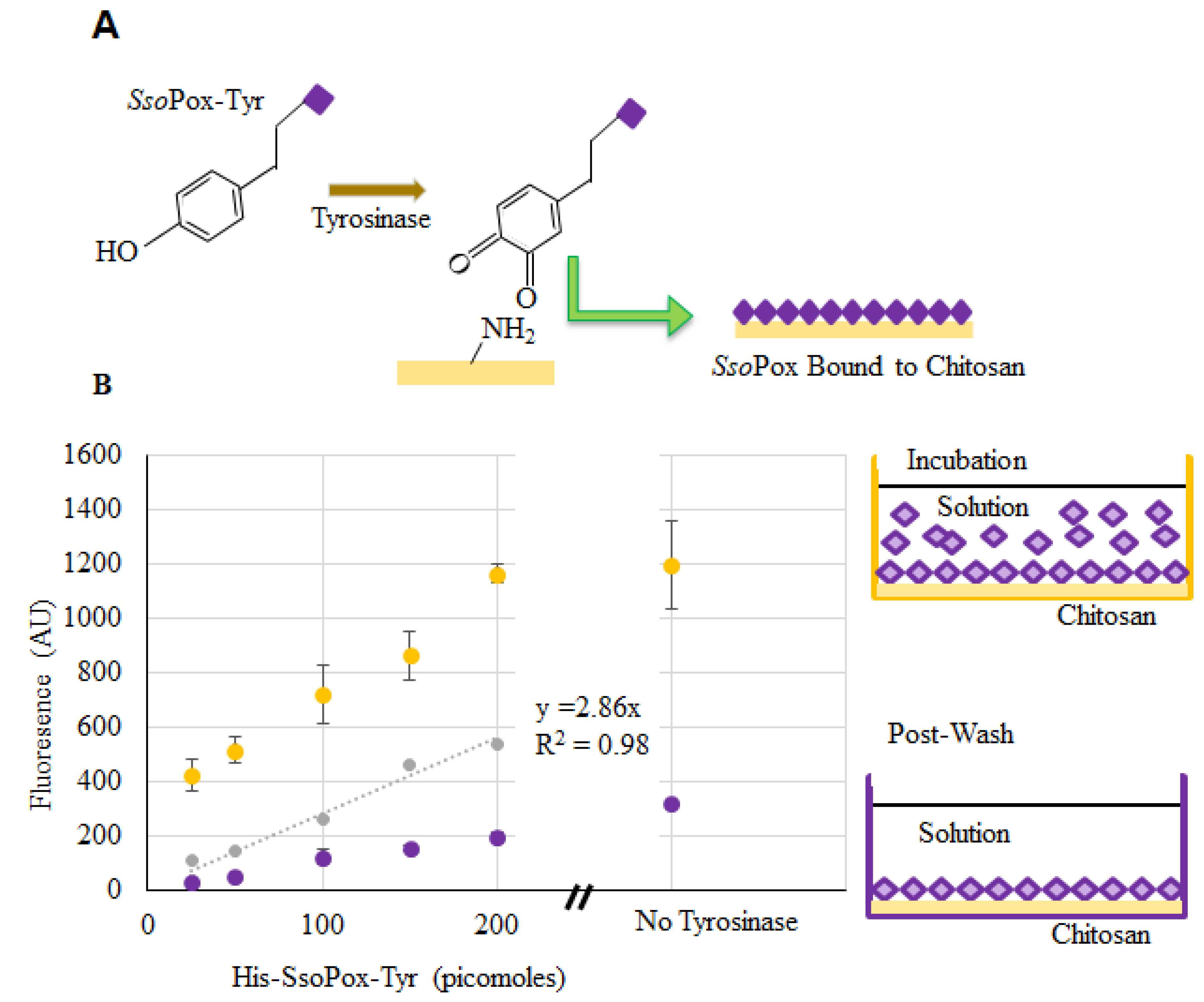
4.5. Binding SsoPox-Tyr to Chitosan
4.6. Binding SsoPox-Gln to Chitosan
4.7. Capsule Construction Materials
4.8. Calculating Bound SsoPox
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- World Health Organization (WHO). Media Centre: Fact Sheet: Antibiotic Resistance. Available online: http://www.who.int/mediacentre/factsheets/antibiotic-resistance/en/ (accessed on 30 December 2017).
- Uroz, S.; Dessaux, Y.; Oger, P. Quorum sensing and quorum quenching: The yin and yang of bacterial communication. Chembiochem 2009, 10, 205–216. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.H.; Wang, L.H.; Xu, J.L.; Zhang, H.B.; Zhang, X.F.; Zhang, L.H. Quenching quorum-sensing-dependent bacterial infection by an N-acyl homoserine lactonase. Nature 2001, 411, 813–817. [Google Scholar] [CrossRef] [PubMed]
- Roy, V.; Smith, J.A.; Wang, J.; Stewart, J.E.; Bentley, W.E.; Sintim, H.O. Synthetic analogs tailor native AI-2 signaling across bacterial species. J. Am. Chem. Soc. 2010, 132, 11141–11150. [Google Scholar] [CrossRef] [PubMed]
- Roy, V.; Fernandes, R.; Tsao, C.Y.; Bentley, W.E. Cross species quorum quenching using a native AI-2 processing enzyme. ACS Chem. Biol. 2010, 5, 223–232. [Google Scholar] [CrossRef] [PubMed]
- Roy, V.; Meyer, M.T.; Smith, J.A.; Gamby, S.; Sintim, H.O.; Ghodssi, R.; Bentley, W.E. AI-2 analogs and antibiotics: A synergistic approach to reduce bacterial biofilms. Appl. Microbiol. Biotechnol. 2013, 97, 2627–2638. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, S.; Gerasopoulos, K.; Guo, M.; Sintim, H.O.; Bentley, W.E.; Ghodssi, R. Autoinducer-2 analogs and electric fields—An antibiotic-free bacterial biofilm combination treatment. Biomed. Microdevices 2016, 18, 95. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.H.; Zhang, L.H. Quorum sensing and quorum-quenching enzymes. J. Microbiol. 2005, 43, 101–109. [Google Scholar] [PubMed]
- Bzdrenga, J.; Daude, D.; Remy, B.; Jacquet, P.; Plener, L.; Elias, M.; Chabriere, E. Biotechnological applications of quorum quenching enzymes. Chem. Biol. Interact. 2017, 267, 104–115. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.H.; Gusti, A.R.; Zhang, Q.; Xu, J.L.; Zhang, L.H. Identification of quorum-quenching N-acyl homoserine lactonases from bacillus species. Appl. Environ. Microbiol. 2002, 68, 1754–1759. [Google Scholar] [CrossRef] [PubMed]
- Barrios, A.F.; Covo, V.; Medina, L.M.; Vives-Florez, M.; Achenie, L. Quorum quenching analysis in Pseudomonas aeruginosa and Escherichia coli: Network topology and inhibition mechanism effect on the optimized inhibitor dose. Bioprocess Biosyst. Eng. 2009, 32, 545–556. [Google Scholar] [CrossRef] [PubMed]
- Brackman, G.; Coenye, T. Quorum sensing inhibitors as anti-biofilm agents. Curr. Pharm. Des. 2015, 21, 5–11. [Google Scholar] [CrossRef] [PubMed]
- WHO. Exposure to highly hazardous pesticides: A major public health conern. In Preventing Disease Through Healthy Environments; WHO: Geneva, Switzerland, 2010. [Google Scholar]
- Raushel, F.M. Bacterial detoxification of organophosphate nerve agents. Curr. Opin. Microbiol. 2002, 5, 288–295. [Google Scholar] [CrossRef]
- López-Granero, C.; Cardona, D.; Giménez, E.; Lozano, R.; Barril, J.; Aschner, M.; Sánchez-Santed, F.; Cañadas, F. Comparative study on short- and long-term behavioral consequences of organophosphate exposure: Relationship to AChE mRNA expression. Neurotoxicology 2014, 40, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Costa, L.G. Current issues in organophosphate toxicology. Clin. Chim. Acta 2006, 366, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Aubert, S.D.; Li, Y.; Raushel, F.M. Mechanism for the hydrolysis of organophosphates by the bacterial phosphotriesterase. Biochemistry 2004, 43, 5707–5715. [Google Scholar] [CrossRef] [PubMed]
- Afriat-Jurnou, L.; Jackson, C.J.; Tawfik, D.S. Reconstructing a missing link in the evolution of a recently diverged phosphotriesterase by active-site loop remodeling. Biochemistry 2012, 51, 6047–6055. [Google Scholar] [CrossRef] [PubMed]
- Tsai, P.-C.; Fox, N.; Bigley, A.N.; Harvey, S.P.; Barondeau, D.P.; Raushel, F.M. Enzymes for the homeland defense: Optimizing phosphotriesterase for the hydrolysis of organophosphate nerve agents. Biochemistry 2012, 51, 6463–6475. [Google Scholar] [CrossRef] [PubMed]
- Elias, M.; Tawfik, D.S. Divergence and convergence in enzyme evolution: Parallel evolution of paraoxonases from quorum-quenching lactonases. J. Biol. Chem. 2012, 287, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Del Vecchio, P.; Elias, M.; Merone, L.; Graziano, G.; Dupuy, J.; Mandrich, L.; Carullo, P.; Fournier, B.; Rochu, D.; Rossi, M.; et al. Structural determinants of the high thermal stability of ssopox from the hyperthermophilic archaeon sulfolobus solfataricus. Extremophiles 2009, 13, 461–470. [Google Scholar] [CrossRef] [PubMed]
- Del Giudice, I.; Coppolecchia, R.; Merone, L.; Porzio, E.; Carusone, T.M.; Mandrich, L.; Worek, F.; Manco, G. An efficient thermostable organophosphate hydrolase and its application in pesticide decontamination. Biotechnol. Bioeng. 2015, 113, 724–734. [Google Scholar] [CrossRef] [PubMed]
- Hiblot, J.; Gotthard, G.; Chabriere, E.; Elias, M. Characterisation of the organophosphate hydrolase catalytic activity of SsoPox. Sci. Rep. 2012, 2. [Google Scholar] [CrossRef] [PubMed]
- Hiblot, J.; Gotthard, G.; Elias, M.; Chabriere, E. Differential active site loop conformations mediate promiscuous activities in the lactonase SsoPox. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [PubMed]
- Elias, M.; Dupuy, J.; Merone, L.; Mandrich, L.; Porzio, E.; Moniot, S.; Rochu, D.; Lecomte, C.; Rossi, M.; Masson, P.; et al. Structural basis for natural lactonase and promiscuous phosphotriesterase activities. J. Mol. Biol. 2008, 379, 1017–1028. [Google Scholar] [CrossRef] [PubMed]
- Ng, F.S.W.; Wright, D.M.; Seah, S.Y.K. Characterization of a phosphotriesterase-like lactonase from sulfolobus solfataricus and its immobilization for disruption of quorum sensing. Appl. Environ. Microbiol. 2011, 77, 1181–1186. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.-C.; Shi, X.-W.; Tsao, C.-Y.; Lewandowski, A.T.; Fernandes, R.; Hung, C.-W.; DeShong, P.; Kobatake, E.; Valdes, J.J.; Payne, G.F.; et al. Biofabrication of antibodies and antigens via IgG-binding domain engineered with activatable pentatyrosine pro-tag. Biotechnol. Bioeng. 2009, 103, 231–240. [Google Scholar] [CrossRef] [PubMed]
- Bhokisham, N.; Pakhchanian, H.; Quan, D.; Tschirhart, T.; Tsao, C.-Y.; Payne, G.F.; Bentley, W.E. Modular construction of multi-subunit protein complexes using engineered tags and microbial transglutaminase. Metab. Eng. 2016, 38, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Lewandowski, A.T.; Small, D.A.; Chen, T.; Payne, G.F.; Bentley, W.E. Tyrosine-based “activatable pro-tag”: Enzyme-catalyzed protein capture and release. Biotechnol. Bioeng. 2006, 93, 1207–1215. [Google Scholar] [CrossRef] [PubMed]
- Lewandowski, A.T.; Yi, H.; Luo, X.; Payne, G.F.; Ghodssi, R.; Rubloff, G.W.; Bentley, W.E. Protein assembly onto patterned microfabricated devices through enzymatic activation of fusion pro-tag. Biotechnol. Bioeng. 2008, 99, 499–507. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Deng, S.; Wei, H.; Wang, B.; Huang, J.; Yu, G. Activated carbons and amine-modified materials for carbon dioxide capture—A review. Front. Environ. Sci. Eng. 2013, 7, 326–340. [Google Scholar] [CrossRef]
- Zargar, A.; Quan, D.N.; Abutaleb, N.; Choi, E.; Terrell, J.L.; Payne, G.F.; Bentley, W.E. Constructing “quantized quorums” to guide emergent phenotypes through quorum quenching capsules. Biotechnol. Bioeng. 2017, 114, 407–415. [Google Scholar] [CrossRef] [PubMed]
- Sarmento, B.; das Neves, J. Chitosan-Based Systems for Biopharmaceuticals: Delivery, Targeting and Polymer Therapeutics; John Wiley & Sons: Hoboken, NJ, USA, 2012. [Google Scholar]
- Williams, P.; Cámara, M. Quorum sensing and environmental adaptation in Pseudomonas aeruginosa: A tale of regulatory networks and multifunctional signal molecules. Curr. Opin. Microbiol. 2009, 12, 182–191. [Google Scholar] [CrossRef] [PubMed]
- Schuster, M.; Urbanowski, M.L.; Greenberg, E.P. Promoter specificity in Pseudomonas aeruginosa quorum sensing revealed by DNA binding of purified LasR. Proc. Natl. Acad. Sci. USA 2004, 101, 15833–15839. [Google Scholar] [CrossRef] [PubMed]
- Borlee, B.R.; Geske, G.D.; Blackwell, H.E.; Handelsman, J. Identification of synthetic inducers and inhibitors of the quorum-sensing regulator LasR in Pseudomonas aeruginosa by high-throughput screening. Appl. Environ. Microbiol. 2010, 76, 8255–8258. [Google Scholar] [CrossRef] [PubMed]
- Alagely, A.; Rajamani, S.; Teplitski, M. Luminescent reporters and their applications for the characterization of signals and signal-mimics that alter LasR-mediated quorum sensing. Methods Mol. Biol. 2011, 692, 113–130. [Google Scholar] [PubMed]
- Lindsay, A.; Ahmer, B.M. Effect of sdia on biosensors of N-acylhomoserine lactones. J. Bacteriol. 2005, 187, 5054–5058. [Google Scholar] [CrossRef] [PubMed]
- Rhoads, M.K.; Hauk, P.; Terrell, J.; Tsao, C.Y.; Oh, H.; Raghavan, S.R.; Mansy, S.S.; Payne, G.F.; Bentley, W.E. Incorporating LsrK AI-2 quorum quenching capability in a functionalized biopolymer capsule. Biotechnol. Bioeng. 2017, 115, 278–289. [Google Scholar] [CrossRef] [PubMed]
- Bhokisham, N.; Liu, Y.; Pakhchanian, H.; Payne, G.F.; Bentley, W.E. A facile two-step enzymatic approach for conjugating proteins to polysaccharide chitosan and an electrode interface. Cell. Mol. Bioeng. 2017, 10, 134–142. [Google Scholar] [CrossRef]
- Bhokisham, N.; Pakhchanian, H.; Quan, D.; Tschirhart, T.; Tsao, C.Y.; Payne, G.F.; Bentley, W.E. Data on biochemical fluxes generated from biofabricated enzyme complexes assembled through engineered tags and microbial transglutaminase. Data Brief 2016, 8, 1031–1035. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Terrell, J.L.; Fernandes, R.; Dowling, M.B.; Payne, G.F.; Raghavan, S.R.; Bentley, W.E. Encapsulated fusion protein confers “sense and respond” activity to chitosan–alginate capsules to manipulate bacterial quorum sensing. Biotechnol. Bioeng. 2013, 110, 552–562. [Google Scholar] [CrossRef] [PubMed]
- Takka, S.; Gürel, A. Evaluation of chitosan/alginate beads using experimental design: Formulation and in vitro characterization. AAPS PharmSciTech 2010, 11, 460–466. [Google Scholar] [CrossRef] [PubMed]
- González-Rodríguez, M.L.; Holgado, M.A.; Sánchez-Lafuente, C.; Rabasco, A.M.; Fini, A. Alginate/chitosan particulate systems for sodium diclofenac release. Int. J. Pharm. 2002, 232, 225–234. [Google Scholar] [CrossRef]
- Servinsky, M.D.; Terrell, J.L.; Tsao, C.Y.; Wu, H.C.; Quan, D.N.; Zargar, A.; Allen, P.C.; Byrd, C.M.; Sund, C.J.; Bentley, W.E. Directed assembly of a bacterial quorum. ISME J. 2016, 10, 158–169. [Google Scholar] [CrossRef] [PubMed]
- Hentzer, M.; Givskov, M. Pharmacological inhibition of quorum sensing for the treatment of chronic bacterial infections. J. Clin. Investig. 2003, 112, 1300–1307. [Google Scholar] [CrossRef] [PubMed]
- Hentzer, M.; Riedel, K.; Rasmussen, T.B.; Heydorn, A.; Andersen, J.B.; Parsek, M.R.; Rice, S.A.; Eberl, L.; Molin, S.; Høiby, N.; et al. Inhibition of quorum sensing in Pseudomonas aeruginosa biofilm bacteria by a halogenated furanone compound. Microbiology 2002, 148, 87–102. [Google Scholar] [CrossRef] [PubMed]
- Chambers, C.E.; Visser, M.B.; Schwab, U.; Sokol, P.A. Identification of N-acylhomoserine lactones in mucopurulent respiratory secretions from cystic fibrosis patients. FEMS Microbiol. Lett. 2005, 244, 297–304. [Google Scholar] [CrossRef] [PubMed]
- Telford, G.; Wheeler, D.; Williams, P.; Tomkins, P.T.; Appleby, P.; Sewell, H.; Stewart, G.S.A.B.; Bycroft, B.W.; Pritchard, D.I. The Pseudomonas aeruginosaquorum-sensing signal moleculen-(3-oxododecanoyl)-l-homoserine lactone has immunomodulatory activity. Infect. Immun. 1998, 66, 36–42. [Google Scholar] [PubMed]
Sample Availability: Samples of the compounds (chitosan, plasmids, cells) are available from the authors. |



© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rhoads, M.K.; Hauk, P.; Gupta, V.; Bookstaver, M.L.; Stephens, K.; Payne, G.F.; Bentley, W.E. Modification and Assembly of a Versatile Lactonase for Bacterial Quorum Quenching. Molecules 2018, 23, 341. https://doi.org/10.3390/molecules23020341
Rhoads MK, Hauk P, Gupta V, Bookstaver ML, Stephens K, Payne GF, Bentley WE. Modification and Assembly of a Versatile Lactonase for Bacterial Quorum Quenching. Molecules. 2018; 23(2):341. https://doi.org/10.3390/molecules23020341
Chicago/Turabian StyleRhoads, Melissa K., Pricila Hauk, Valerie Gupta, Michelle L. Bookstaver, Kristina Stephens, Gregory F. Payne, and William E. Bentley. 2018. "Modification and Assembly of a Versatile Lactonase for Bacterial Quorum Quenching" Molecules 23, no. 2: 341. https://doi.org/10.3390/molecules23020341