
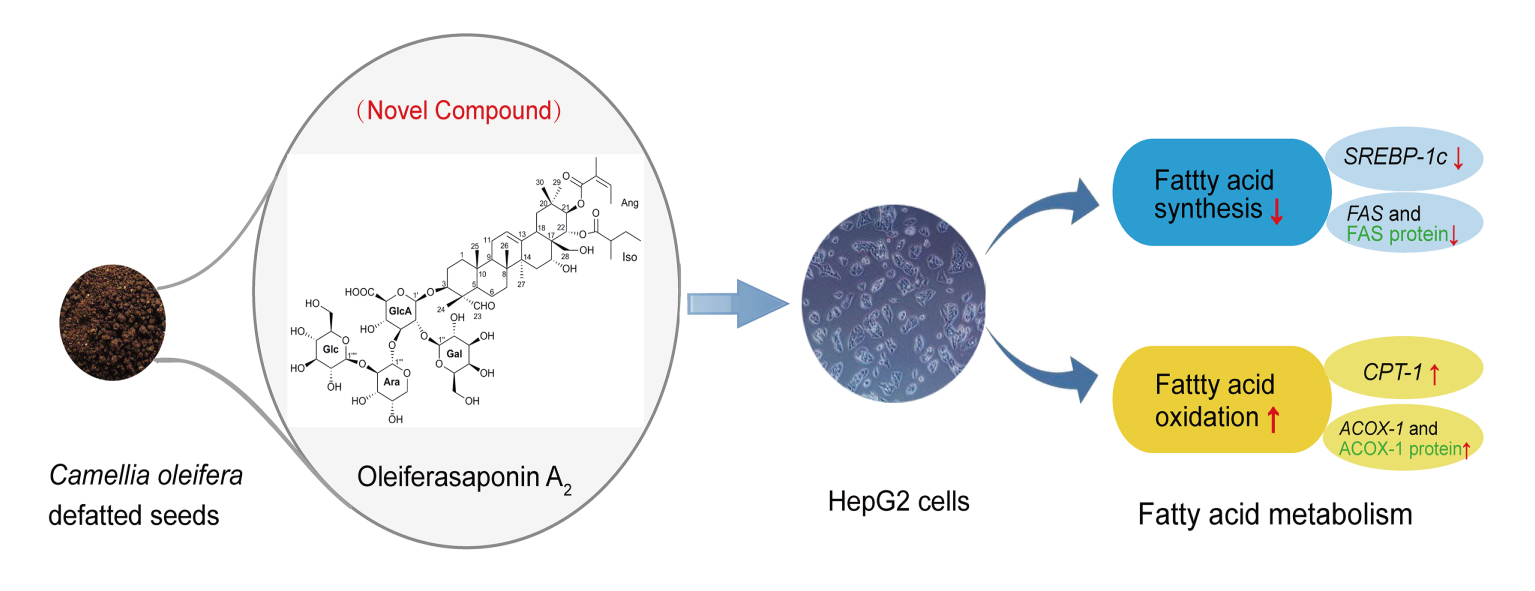
Oleiferasaponin A2, a Novel Saponin from Camellia oleifera Abel. Seeds, Inhibits Lipid Accumulation of HepG2 Cells Through Regulating Fatty Acid Metabolism

 and
and
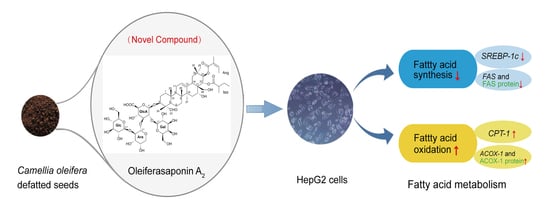
Abstract
:
1. Introduction
2. Results and Discussion
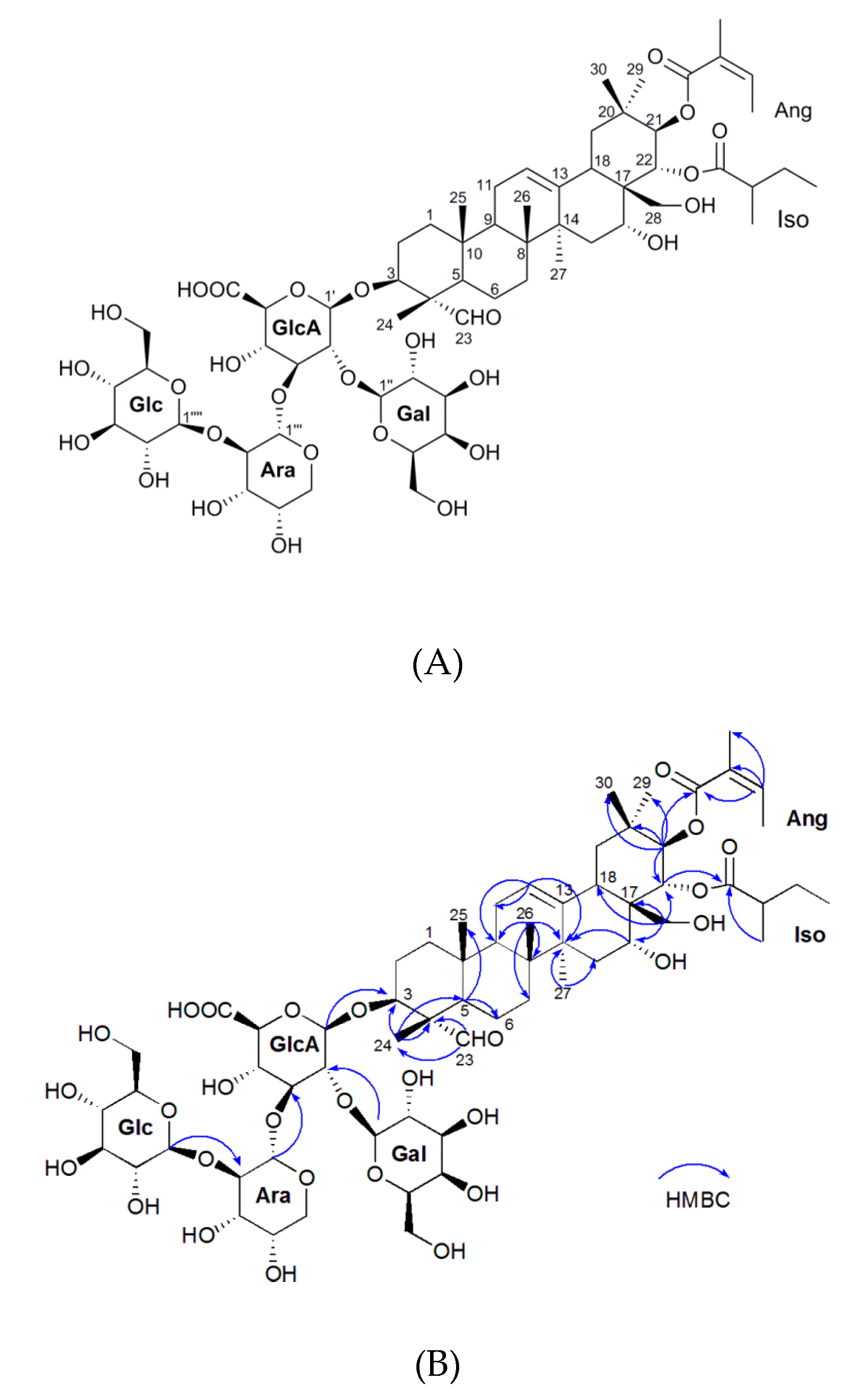
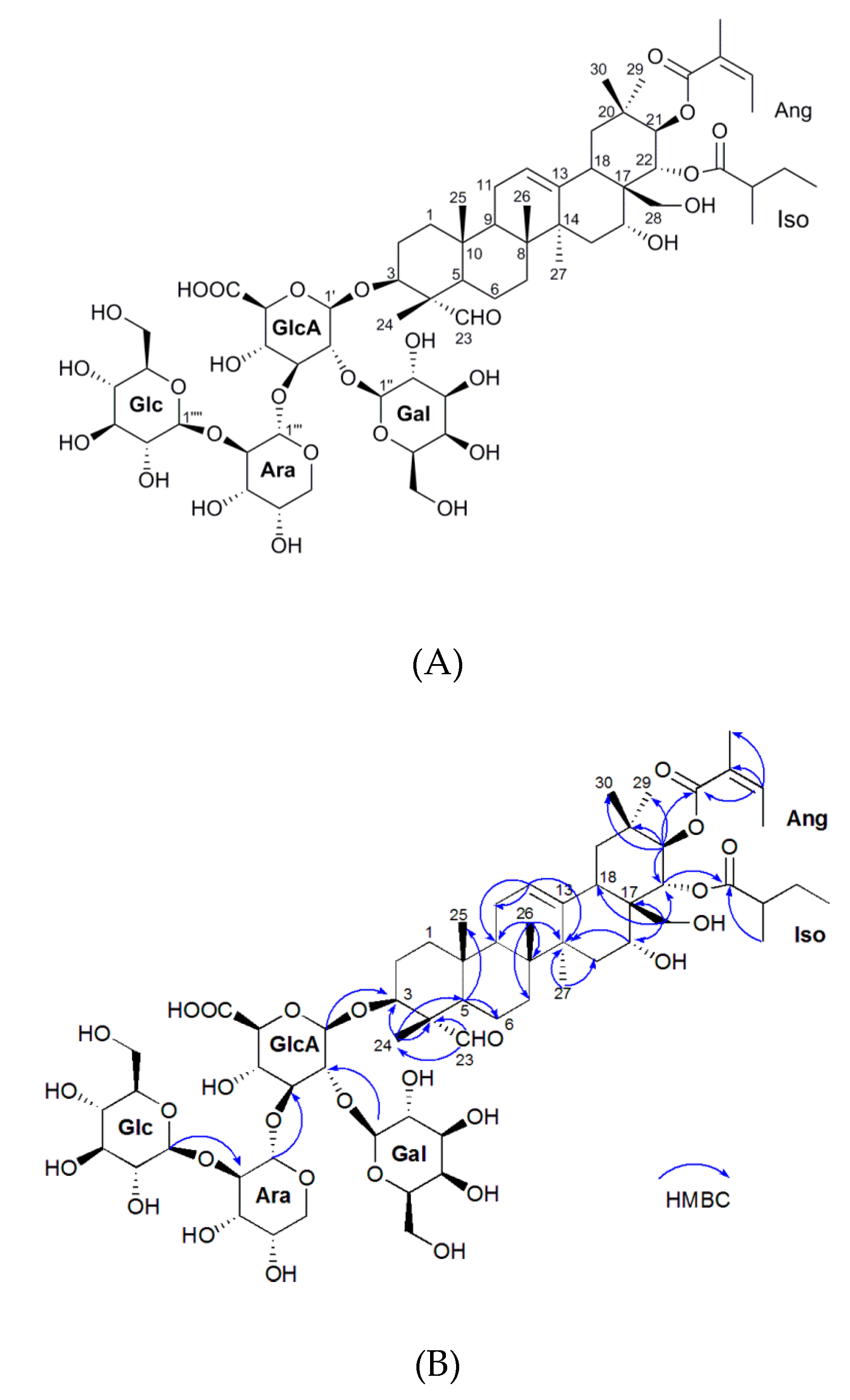
2.1. Isolation and Characterization of the Oleiferasaponin A2
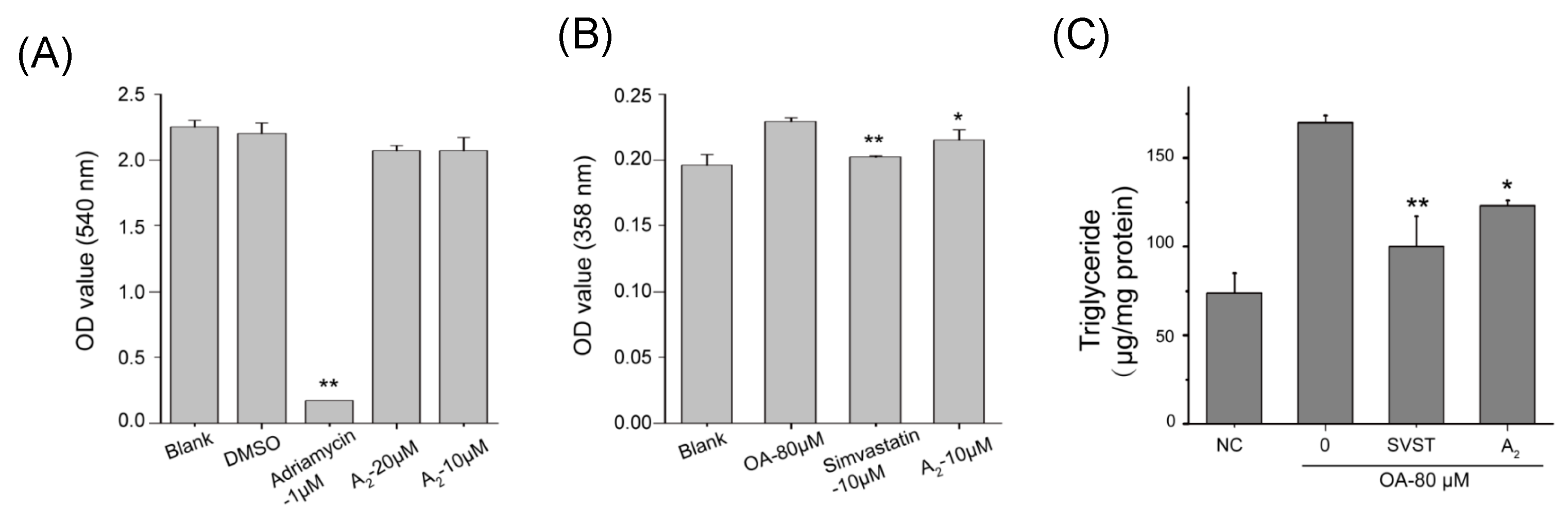
2.2. Hypolipidemic Activity of the Oleiferasaponin A2
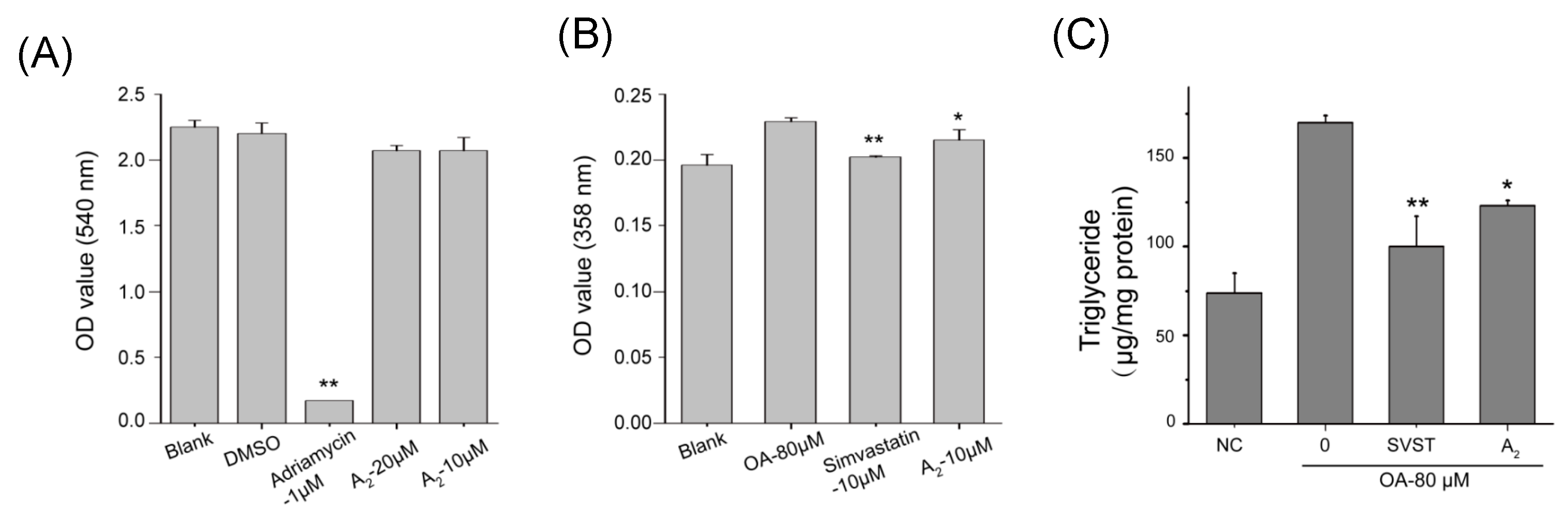
2.2.1. Oleiferasaponin A2 Exhibited Hypolipidemic Activity on HepG2 Cell Lines
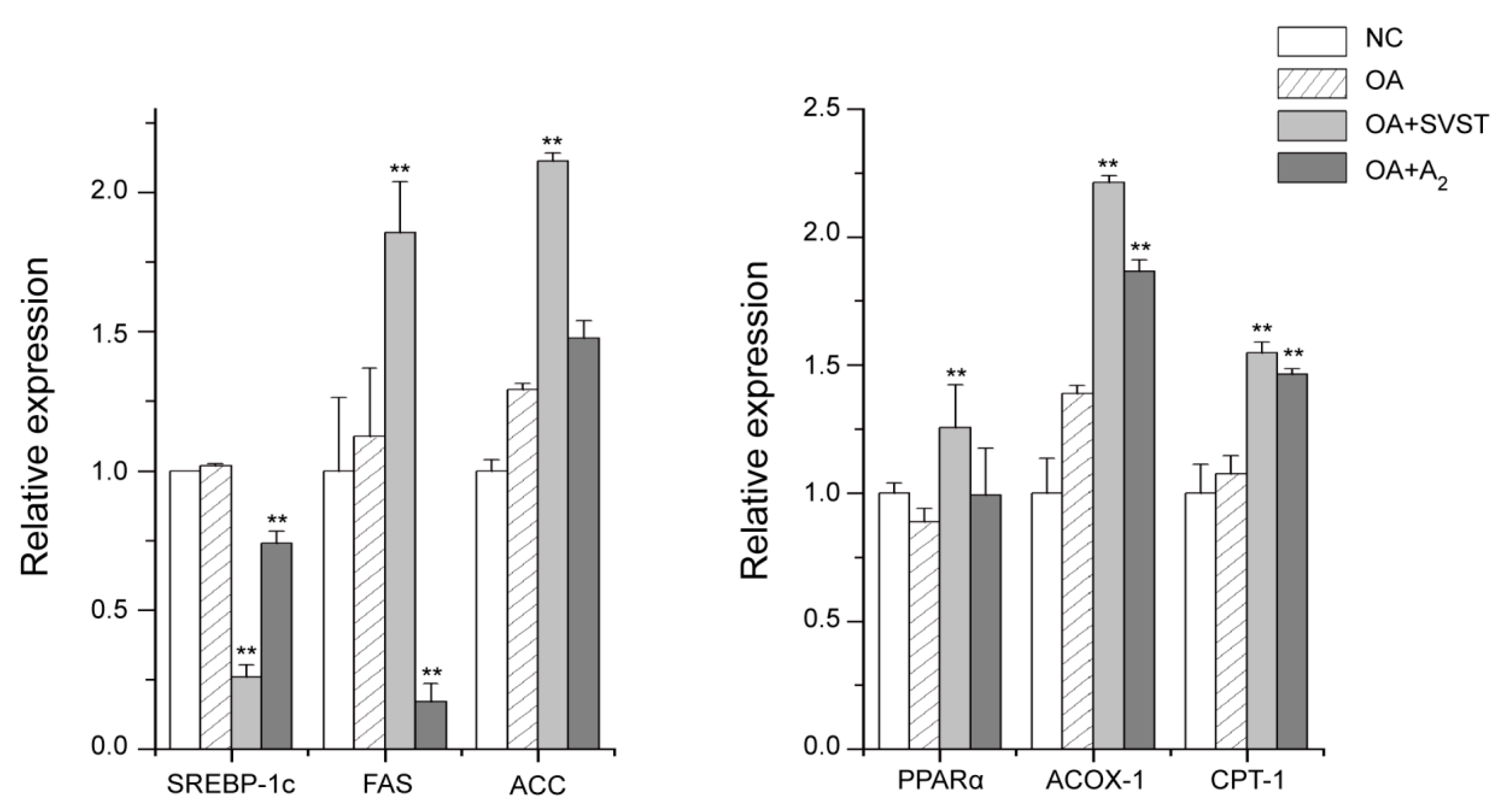
2.2.2. Oleiferasaponin A2 Affected the Expression of Genes Related to Fatty Acid Metabolism
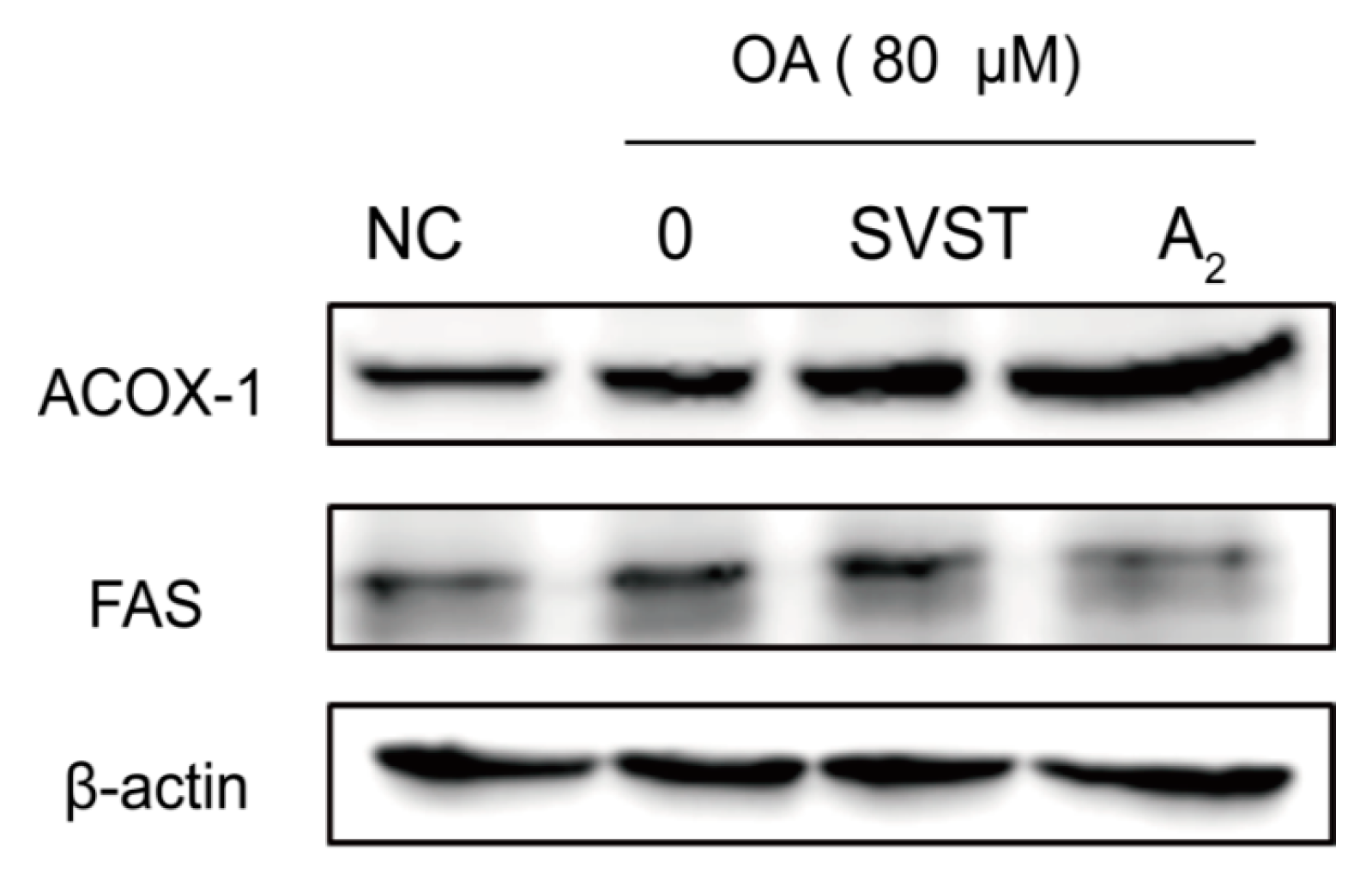
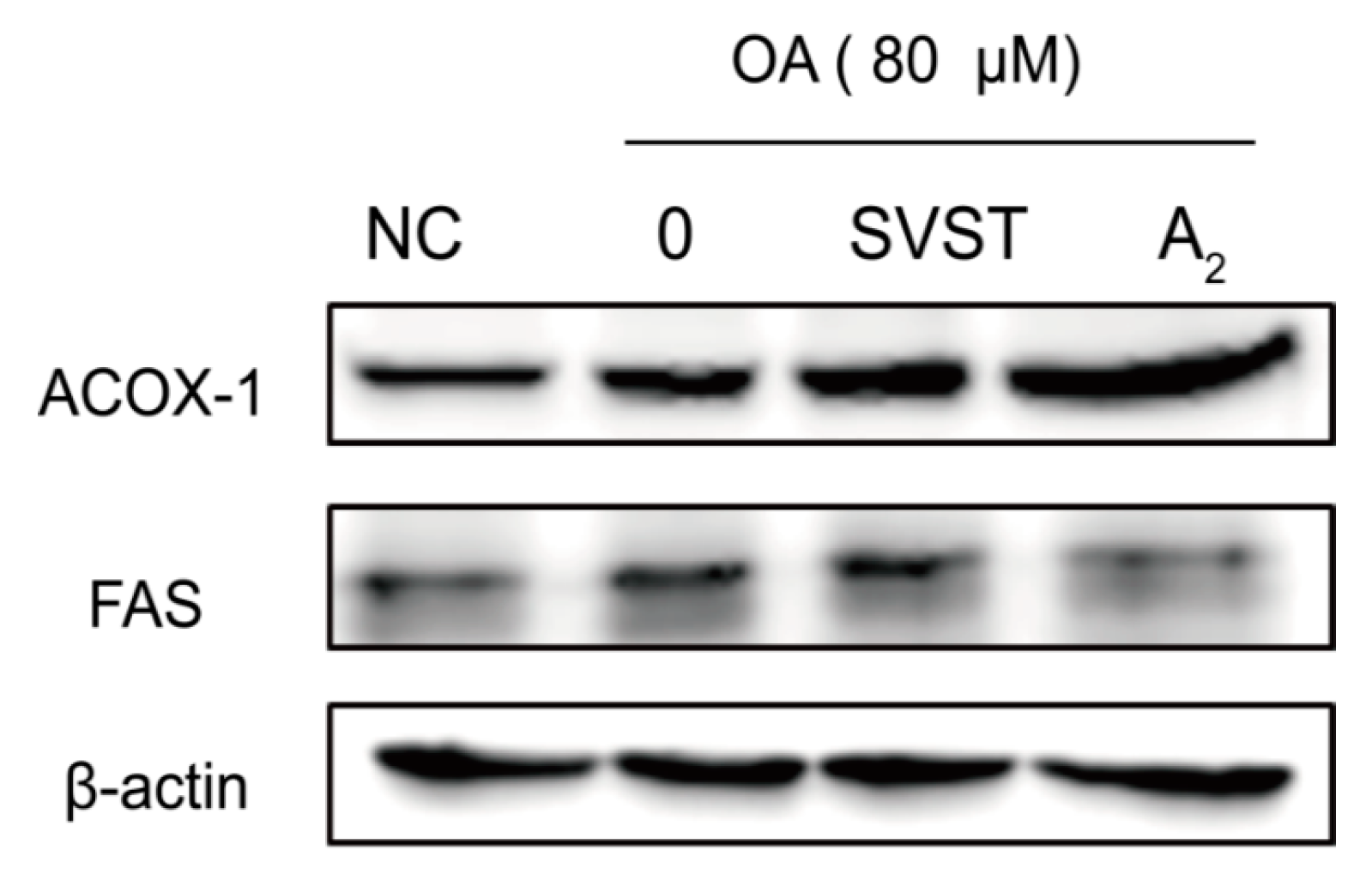
2.2.3. Oleiferasaponin A2 Affected the Expression of Proteins Related to Fatty Acid Metabolism
3. Materials and Methods
3.1. General
3.2. Plant Material
3.3. Extraction and Isolation
3.4. Acid Hydrolysis and GC-MS Analysis
3.5. Cell Viability Assay
3.6. Preliminary Screening of Hypolipidemic Activity
3.7. Triglyceride Test
3.8. Fluorescent Quantitative PCR
3.9. Western Blot Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Melanie, N.; Townsend, N.; Scarnorough, P.; Rayner, M. Cardiovascular disease in Europe 2014: Epidemiological update. Eur. Heart J. 2014, 35, 2950–2959. [Google Scholar]
- Program, L.C. The lipid research clinics coronary primary prevention trial results. I. Reduction in incidence of coronary heart disease. JAMA 1985, 253, 635–636. [Google Scholar]
- Shrestha, S.; Bhattarai, B.R.; Lee, K.H.; Cho, H. Mono- and disalicylic acid derivatives: Ptp1b inhibitors as potential anti-obesity drugs. Bioorgan. Med. Chem. 2007, 15, 6535–6548. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.B.; Nasir, A.; Prabhu, K.M.; Murthy, P.S.; Dev, G. Hypoglycemic and hypolipidemic effect of ethanolic extract of seeds of eugenia jambolana in alloxan induced diabetic rabbits. J. Ethnopharmacol. 2003, 85, 201–206. [Google Scholar] [CrossRef]
- Zou, B.; Li, C.M.; Chen, J.Y.; Dong, X.Q.; Zhang, Y.; Du, J. High molecular weight persimmon tannin is a potent hypolipidemic in high-cholesterol diet fed rats. Food Res. Int. 2012, 48, 970–977. [Google Scholar] [CrossRef]
- Xia, D.Z.; Yu, X.F.; Wang, H.M.; Ren, Q.Y.; Chen, B.M. Anti-obesity and hypolipidemic effects of ethanolic extract from alpinia officinarum hance (zingiberaceae) in rats fed high-fat diet. J. Med. Food 2010, 13, 785–791. [Google Scholar] [CrossRef]
- Jian, H.L.; Liao, X.X.; Zhu, L.W.; Zhang, W.M.; Jiang, J.X. Synergism and foaming properties in binary mixtures of a biosurfactant derived from camellia oleifera abel and synthetic surfactants. J. Coll. Interface Sci. 2011, 359, 487–492. [Google Scholar] [CrossRef]
- Chen, Y.Z.; Li, M.A.; Liu, Z.C.; Peng, S.F.; Chen, X.L.; Chen, L.S.; Wang, X.N.; Wang, R. Molluscicidal effect of camellia oleifera saponin. J. Cent. South Univ. Technol. 2011, 31, 147–150. [Google Scholar]
- Chen, Y.F.; Yang, C.H.; Chang, M.S.; Ciou, Y.P.; Huang, Y.C. Foam properties and detergent abilities of the saponins from camellia oleifera. Int. J. Mol. Sci. 2010, 11, 4417–4425. [Google Scholar] [CrossRef]
- Kuo, P.C.; Lin, T.C.; Yang, C.W.; Lin, C.L.; Chen, G.F.; Huang, J.W. Bioactive saponin from tea seed pomace with inhibitory effects against rhizoctonia solani. J. Agric. Food Chem. 2010, 58, 8618–8622. [Google Scholar] [CrossRef]
- Chen, L.; Chen, J.; Xu, H. Sasanquasaponin from camellia oleifera abel. Induces cell cycle arrest and apoptosis in human breast cancer mcf-7 cells. Fitoterapia 2013, 84, 123–129. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.H.; Wu, H.Y.; Bingchung, L.; Chang, C.M.J.; Tingting, J.; Wu, L.C. Identification and evaluation of antioxidants defatted camellia oleifera seeds by isopropanol salting-out pretreatment. Food Chem. 2010, 121, 1246–1254. [Google Scholar] [CrossRef]
- Zhang, X.F.; Han, Y.Y.; Bao, G.H.; Ling, T.J.; Zhang, L.; Gao, L.P.; Xia, T. A new saponin from tea seed pomace (camellia oleifera abel) and its protective effect on pc12 cells. Molecules 2012, 17, 11721. [Google Scholar] [CrossRef] [PubMed]
- Zong, J.; Wang, R.; Bao, G.; Ling, T.; Zhang, L.; Zhang, X.; Hou, R. Novel triterpenoid saponins from residual seed cake of camellia oleifera abel. Show anti-proliferative activity against tumor cells. Fitoterapia 2015, 104, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Zong, J.F.; Peng, Y.R.; Bao, G.H.; Hou, R.Y.; Wan, X.C. Two new oleanane-type saponins with anti-proliferative activity from camellia oleifera abel. Seed cake. Molecules 2016, 21, 188–195. [Google Scholar] [CrossRef] [PubMed]
- Zong, J.; Wang, D.; Jiao, W.; Zhang, L.; Bao, G.; Ho, C.T.; Hou, R.; Wan, X. Oleiferasaponin c6 from the seeds of camellia oleifera abel.: A novel compound inhibits proliferation through inducing cell-cycle arrest and apoptosis on human cancer cell lines in vitro. RSC Adv. 2016, 6, 91386–91393. [Google Scholar] [CrossRef]
- Zhou, H.; Wang, C.Z.; Ye, J.Z.; Chen, H.X. New triterpene saponins from the seed cake of camellia oleifera and their cytotoxic activity. Phytochem. Lett. 2014, 8, 46–51. [Google Scholar] [CrossRef]
- Di, T.M.; Yang, S.L.; Du, F.Y.; Zhao, L.; Xia, T.; Zhang, X.F. Cytotoxic and hypoglycemic activity of triterpenoid saponins from camellia oleifera abel. Seed pomace. Molecules 2017, 22, 1562. [Google Scholar] [CrossRef]
- Wei, Y.Y.; Cai, H.Y.; Wang, L.S.; Xia, T.; Xian-Feng, D.U. Production of multi-enzyme enriched bio-feed from camellia oleifera seed cake in solid state fermentation by aspergillus niger. J. Anhui Agric. Univ. 2012, 39, 41–46. [Google Scholar]
- Hu, J.L.; Nie, S.P.; Huang, D.F.; Chang, L.; Xie, M.Y.; Yin, W. Antimicrobial activity of saponin—rich fraction from camellia oleifera cake and its effect on cell viability of mouse macrophage raw 264.7. J. Sci. Food Agric. 2012, 92, 2443–2449. [Google Scholar] [CrossRef]
- Zhang, X.F.; Yang, S.L.; Han, Y.Y.; Zhao, L.; Lu, G.L.; Xia, T.; Gao, L.P. Qualitative and quantitative analysis of triterpene saponins from tea seed pomace (camellia oleifera abel) and their activities against bacteria and fungi. Molecules 2014, 19, 7568–7580. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, H.; Nakamura, S.; Fujimoto, K.; Moriuchi, R.; Kimura, Y.; Ikoma, N.; Hata, Y.; Muraoka, O.; Yoshikawa, M. Medicinal flowers. Xxxi. Acylated oleanane-type triterpene saponins, sasanquasaponins i-v, with antiallergic activity from the flower buds of camellia sasanqua. Chem. Pharm. Bull. 2010, 58, 1617–1621. [Google Scholar] [CrossRef]
- Hu, J.; Nie, S.; Huang, D.; Li, C.; Xie, M. Extraction of saponin from camellia oleifera cake and evaluation of its antioxidant activity. Int. J. Food Sci. Technol. 2012, 47, 1676–1687. [Google Scholar] [CrossRef]
- Mu, L.H.; Huang, C.L.; Zhou, W.B.; Guo, D.H.; Liu, P. Methanolysis of triterpenoid saponin from ardisia gigantifolia stapf. And structure–activity relationship study against cancer cells. Bioorgan. Med. Chem. Lett. 2013, 23, 6073–6078. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.; Akhtar, M.S.; Khan, B.A.; Braga, V.D.A.; Reich, A. Antiobesity, hypolipidemic, antioxidant and hepatoprotective effects of achyranthes aspera seed saponins in high cholesterol fed albino rats. Arch. Med. Sci. Ams 2015, 11, 1261–1271. [Google Scholar] [CrossRef] [PubMed]
- Weng, Y.; Yu, L.; Cui, J.; Zhu, Y.R.; Guo, C.; Wei, G.; Duan, J.L.; Yin, Y.; Guan, Y.; Wang, Y.H. Antihyperglycemic, hypolipidemic and antioxidant activities of total saponins extracted from aralia taibaiensis in experimental type 2 diabetic rats. J. Ethnopharmacol. 2014, 152, 553–560. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, M.; Morikawa, T.; Yamamoto, K.; Kato, Y.; Akifumi Nagatomo, A.; Matsuda, H. Floratheasaponins a–c, acylated oleanane-type triterpene oligoglycosides with anti-hyperlipidemic activities from flowers of the tea plant (camellia sinensis)1. J. Nat. Prod. 2005, 68, 1360–1365. [Google Scholar] [CrossRef] [PubMed]
- Ijeh, I.; Inyang, E.; Egedigwe, A.; Ahaiwe, U. Hypoglycaemic and hypolipidemic effects of crude saponins extracted from vernonia colorata in normoglycaemic albino rats (259.6). FASEB J. [CrossRef]
- Song, C.; Yu, Q.; Li, X.; Jin, S.; Li, S.; Zhang, Y.; Jia, S.; Chen, C.; Xiang, Y.; Jiang, H. The hypolipidemic effect of total saponins from kuding tea in high-fat diet-induced hyperlipidemic mice and its composition characterized by uplc-qtof-ms/ms. J. Food Sci. 2016, 81, H1313–H1319. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Vermeer, M.A.; Bos, W. Molecular structures of citrus flavonoids determine their effects on lipid metabolism in hepg2 cells by primarily suppressing apob secretion. J. Agric. Food Chem. 2011, 59, 4496–4503. [Google Scholar] [CrossRef] [PubMed]
- Sangkitikomol, W.; Rocejanasaroj, A.; Tencomnao, T. Effect of moringa oleifera on advanced glycation end-product formation and lipid metabolism gene expression in hepg2 cells. Genet. Mol. Res. 2014, 13, 723–735. [Google Scholar] [CrossRef]
- Niu, Y.; Lü, N.; Li, Y.; Zhao, D.; Sun, C. Establishment of a model for evaluating hypolipidemic effect in hepg2 cells. J. Hyg. Res. 2010, 39, 155–158. [Google Scholar]
- Kim, K.Y.; Park, K.I.; Lee, S.G.; Baek., S.Y.; Lee, E.H.; Kim, S.C.; Kim, S.H.; Park, S.G.; Yu, S.N.; Oh, T.W.; et al. Deoxypodophyllotoxin in anthriscus sylvestris alleviates fat accumulation in the liver via amp-activated protein kinase, impeding srebp-1c signal. Chem.-Biol. Interact. 2018, 294, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Higuchi, N.; Kato, M.; Shundo, Y.; Tajiri, H.; Tanaka, M.; Yamashita, N.; Kohjima, M.; Kotoh, K.; Nakamuta, M.; Takayanagi, R.; et al. Liver x receptor in cooperation with srebp-1c is a major lipid synthesis regulator in nonalcoholic fatty liver disease. Hepatol. Res. 2008, 38, 1122–1129. [Google Scholar] [CrossRef] [PubMed]
- Cabruja, M.; Momdino, S.; Tsai, Y.T.; Lara, J.; Gramajo, H.; Gago, G. A conditional mutant of the fatty acid synthase unveils unexpected cross talks in mycobacterial lipid metabolism. Open Biol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Greenwood, J.; Bhat, S.; Huang, X.; Wang, R.; Paul, D.; Tong, L.; Saha, A.; Westlin, W.; Kapeller, R.; Harwood, H. Acetyl-coa carboxylase inhibition by nd-630 reduces hepatic steatosis, improves insulin sensitivity, and modulates dyslipidemia in rats. Proc. Nat. Acad. Sci. USA 2016, 113, E1796–E1805. [Google Scholar]
- Ge, J.; Miao, J.J.; Sun, X.Y.; Yu, J.Y. Huangkui capsule, an extract from Abelmoschus manihot (L.) medic, improves diabetic nephropathy via activating peroxisome proliferator-activated receptor (ppar)-α/γ and attenuating endoplasmic reticulum stress in rats. J. Ethnopharmacol. 2016, 189, 238–249. [Google Scholar] [CrossRef]
- Jung, Y.Y.; Lee, Y.K.; Koo, J.S. Expression of lipid metabolism-related proteins in breast phyllodes tumors. Neoplasma 2016, 63, 254–262. [Google Scholar] [CrossRef]
- Liu, X.J.; Duan, N.N.; Liu, C.L.; Niu, C.; Liu, X.P.; Wu, J. Characterization of a murine nonalcoholic steatohepatitis model induced by high fat high calorie diet plus fructose and glucose in drinking water. Lab. Investing. J. Tech. Methods Pathol. 2018, 98, 1184–1199. [Google Scholar] [CrossRef]
- Yoshikawa, M.; Harada, E.; Murakami, T.; Matsuda, H.; Yamahara, J.; Murakami, N. Camelliasaponins b1, b2, c1 and c2, new type inhibitors of ethanol absorption in rats from the seeds of Camellia japonica L. Chem. Pharm. Bull. 1994, 42, 742–744. [Google Scholar] [CrossRef]
- Wang, P.; Ownby, S.; Zhang, Z.; Yuan, W.; Li, S. Cytotoxicity and inhibition of DNA topoisomerase i of polyhydroxylated triterpenoids and triterpenoid glycosides. Bioorgan. Med. Chem. Lett. 2010, 20, 2790–2796. [Google Scholar] [CrossRef]
- Lee, K.I.; Choi, S.U.; Lee, K.R. Triterpene saponins from pleurospermum kamtschaticum and their biological activity. Chem. Pharm. Bull. 2012, 60, 1011–1018. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, H.; Hamao, M.; Nakamura, S.; Kon’I, H.; Murata, M.; Yoshikawa, M. Medicinal flowers. Xxxiii. Anti-hyperlipidemic and anti-hyperglycemic effects of chakasaponins i-iii and structure of chakasaponin iv from flower buds of chinese tea plant (Camellia sinensis). Chem. Pharm. Bull. 2012, 60, 674–680. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Singh, H.; Jagavelu, K.; Wahajuddin, M.; Hanif, K. Fatty acid synthase modulates proliferation, metabolic functions and angiogenesis in hypoxic pulmonary artery endothelial cells. Eur. J. Pharmacol. 2017, 815, 462–469. [Google Scholar] [CrossRef] [PubMed]
- NHammer, C.; El-Shabrawi, Y.; Schauer, S.; Hiden, M.; Berger, J.; Forss-Petter, S.; Winter, E.; Eferl, R.; Zechner, R.; Hoefler, G. Cdna cloning and analysis of tissue-specific expression of mouse peroxisomal straight-chain acyl-coa oxidase. Eur. J. Biochem. 2010, 267, 1254–1260. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are not available from the authors. |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Position | δC | δH | Position | δC | δH |
|---|---|---|---|---|---|
| 1 | 38.0 | 1.14 m, 1.73 m | 21-O-Ang | ||
| 2 | 24.3 | 1.81 m, 2.05 m | Ang-1 | 167.7 | |
| 3 | 84.9 | 3.89 m | Ang-2 | 127.7 | |
| 4 | 55.0 | Ang-3 | 139.0 | 6.18 m | |
| 5 | 47.5 | 1.38 m | Ang-4 | 14.8 | 1.94 m |
| 6 | 19.8 | 0.96 m | Ang-5 | 19.7 | 1.87 s |
| 7 | 31.8 | 1.29 m, 1.64 m | 22-O-Iso | ||
| 8 | 39.9 | Iso-1 | 177.4 | ||
| 9 | 46.5 | 1.81 m | Iso-2 | 41.3 | 2.33 m |
| 10 | 35.6 | Iso-3 | 26.3 | 1.43 m, 1.65 m | |
| 11 | 23.3 | 1.98 m | Iso-4 | 10.8 | 0.89 m |
| 12 | 123.6 | 5.41, t, 3.7 | Iso-5 | 15.6 | 1.05, d, 6.8 |
| 13 | 141.6 | GlcA-1′ | 103.4 | 4.42, d, 7.7 | |
| 14 | 41.0 | GlcA-2′ | 76.8 | 3.77 m | |
| 15 | 33.4 | 1.37 m, 1.70 m | GlcA-3′ | 81.7 | 3.88 overlap |
| 16 | 68.1 | 4.00 m | GlcA-4′ | 69.1 | 3.84 overlap |
| 17 | 47.2 | GlcA-5′ | 75.5 | 3.56 m | |
| 18 | 39.4 | 2.65 m | GlcA-6′ | 177.4 | |
| 19 | 46.3 | 1.21 m, 2.69 m | Gal-1” | 101.3 | 5.04, d, 7.3 |
| 20 | 35.6 | Gal-2” | 72.1 | 3.50 m | |
| 21 | 78.3 | 5.94, d, 10.1 | Gal-3” | 73.4 | 3.76 m |
| 22 | 73.0 | 5.55, d, 10.1 | Gal-4” | 69.1 | 3.84 m |
| 23 | 209.4 | 9.48 s | Gal-5” | 74.8 | 3.30 m |
| 24 | 9.5 | 1.19 s | Gal-6” | 61.1 | 3.69 m |
| 25 | 15.1 | 2.00 s | Ara-1‴ | 100.2 | 5.03, d, 7.3 |
| 26 | 15.9 | 0.96 s | Ara-2‴ | 82.4 | 3.68 m |
| 27 | 26.4 | 1.53 s | Ara-3‴ | 69.6 | 3.53 m |
| 28 | 63.0 | 2.95 m, 3.77 m | Ara-4‴ | 68.1 | 4.00 m |
| 29 | 28.3 | 0.88 s | Ara-5‴ | 65.9 | 3.21 m, 3.97 m |
| 30 | 19.0 | 1.10 s | Glc-1‴′ | 106.2 | 4.53, d, 7.7 |
| Glc-2‴′ | 75.1 | 3.65 m | |||
| Glc-3‴′ | 73.6 | 3.51 overlap | |||
| Glc-4‴′ | 69.6 | 3.53 overlap | |||
| Glc-5‴′ | 76.4 | 3.36 m | |||
| Glc-6‴′ | 61.2 | 3.80 m |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di, T.-M.; Yang, S.-L.; Du, F.-Y.; Zhao, L.; Li, X.-H.; Xia, T.; Zhang, X.-F. Oleiferasaponin A2, a Novel Saponin from Camellia oleifera Abel. Seeds, Inhibits Lipid Accumulation of HepG2 Cells Through Regulating Fatty Acid Metabolism. Molecules 2018, 23, 3296. https://doi.org/10.3390/molecules23123296
Di T-M, Yang S-L, Du F-Y, Zhao L, Li X-H, Xia T, Zhang X-F. Oleiferasaponin A2, a Novel Saponin from Camellia oleifera Abel. Seeds, Inhibits Lipid Accumulation of HepG2 Cells Through Regulating Fatty Acid Metabolism. Molecules. 2018; 23(12):3296. https://doi.org/10.3390/molecules23123296
Chicago/Turabian StyleDi, Tai-Mei, Shao-Lan Yang, Feng-Yu Du, Lei Zhao, Xiao-Han Li, Tao Xia, and Xin-Fu Zhang. 2018. "Oleiferasaponin A2, a Novel Saponin from Camellia oleifera Abel. Seeds, Inhibits Lipid Accumulation of HepG2 Cells Through Regulating Fatty Acid Metabolism" Molecules 23, no. 12: 3296. https://doi.org/10.3390/molecules23123296