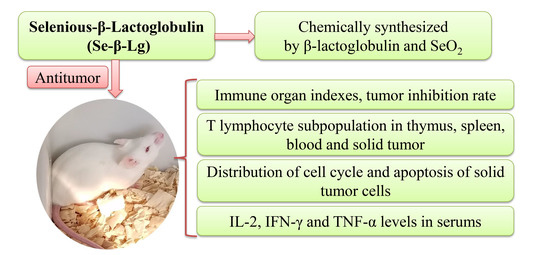
Antitumor and Immunoregulatory Activities of Seleno-β-Lactoglobulin on S180 Tumor-Bearing Mice


Abstract
:
1. Introduction
2. Results and Discussion
2.1. Immune Organ Indexes and Tumor Inhibition Rate of Se-β-lg
2.2. T Lymphocyte Subpopulation of Thymus and Spleen
2.3. T Lymphocyte Subpopulation of Tumor and Peripheral Blood
2.4. Analysis of Cell Cycle Detection in Tumor Cells
2.5. Cytokine Detection
3. Materials and Methods
3.1. Animals and Cells
- Group 1: Normal (N)
- Group 2: Model (M)
- Group 3: β-lg
- Group 4: SeO2
- Group 5: Low Se-β-lg (L-Se-β-lg)
- Group 6: Middle Se-β-lg (M-Se-β-lg)
- Group 7: High Se-β-lg (H-Se-β-lg)
3.2. Thymus Index and Spleen Index
3.3. Measurement of Tumor Weight and Tumor Inhibition Rate
3.4. PI Staining
3.5. Detection of T Lymphocyte Subpopulation
3.6. Detection of IL-2, IFN-γ and TNF-α in Peripheral Blood
3.7. Statistics Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Palomo, M.; Gutierrez, A.M.; Perez-Conde, M.C.; Camara, C.; Madrid, Y. Se metallomics during lactic fermentation of Se-enriched yogurt. Food Chem. 2014, 164, 371–379. [Google Scholar] [CrossRef] [PubMed]
- Skalickova, S.; Milosavljevic, V.; Cihalova, K.; Horky, P.; Richtera, L.; Adam, V. Selenium nanoparticles as a nutritional supplement. Nutrition 2017, 33, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Yao, Z.; Zhang, Y.; Li, H.; Deng, Z.; Zhang, X. Synergistic effect of Se-methylselenocysteine and vitamin E in ameliorating the acute ethanol-induced oxidative damage in rat. J. Trace Elem. Med. Biol. 2015, 29, 182–187. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.R.; Li, Y.; Zhang, X.H.; Wang, M.; Guo, H.R.; Ruan, W.J. Anticancer effect and mechanism of a Se-modified porphyrin Au(III) complex. Bioorg. Med. Chem. Lett. 2015, 25, 3592–3596. [Google Scholar] [CrossRef] [PubMed]
- Avila, F.W.; Yang, Y.; Faquin, V.; Ramos, S.J.; Guilherme, L.R.; Thannhauser, T.W.; Li, L. Impact of selenium supply on Se-methylselenocysteine and glucosinolate accumulation in selenium-biofortified Brassica sprouts. Food Chem. 2014, 165, 578–586. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Sun, S.; Cai, D.; Kong, G. Induction of mitochondrial-dependent apoptosis in T24 cells by a selenium (Se)-containing polysaccharide from Ginkgo biloba L. leaves. Int. J. Biol. Macromol. 2017, 101, 126–130. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, Y.; Liu, Y.; Chen, X.; Wei, X. Extraction, characterization and antioxidant activities of Se-enriched tea polysaccharides. Int. J. Biol. Macromol. 2015, 77, 76–84. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chen, J.; Zhang, D.; Zhang, Y.; Wen, Y.; Li, L.; Zheng, L. Tumoricidal effects of a selenium (Se)-polysaccharide from Ziyang green tea on human osteosarcoma U-2 OS cells. Carbohydr. Polym. 2013, 98, 1186–1190. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Sun, J.; Rao, S.; Su, Y.; Li, J.; Li, C.; Xu, S.; Yang, Y. Antidiabetic activity of mycelia selenium-polysaccharide from catathelasma ventricosum in STZ-induced diabetic mice. Food Chem. Toxicol. 2013, 62, 285–291. [Google Scholar] [CrossRef] [PubMed]
- Mao, G.H.; Ren, Y.; Li, Q.; Wu, H.Y.; Jin, D.; Zhao, T.; Xu, C.Q.; Zhang, D.H.; Jia, Q.D.; Bai, Y.P.; et al. Anti-tumor and immunomodulatory activity of selenium (Se)-polysaccharide from Se-enriched Grifola frondosa. Int. J. Biol. Macromol. 2016, 82, 607–613. [Google Scholar] [CrossRef] [PubMed]
- Yuan, C.; Wang, C.; Wang, J.; Kumar, V.; Anwar, F.; Xiao, F.; Mushtaq, G.; Liu, Y.; Kamal, M.A.; Yuan, D. Inhibition on the growth of human MDA-MB-231 breast cancer cells in vitro and tumor growth in a mouse xenograft model by Se-containing polysaccharides from Pyracantha fortuneana. Nutr. Res. 2016, 36, 1243–1254. [Google Scholar] [CrossRef] [PubMed]
- Indyk, H.E.; Hart, S.; Meerkerk, T.; Gill, B.D.; Woollard, D.C. The β-lactoglobulin content of bovine milk: Development and application of a biosensor immunoassay. Int. Dairy J. 2017, 73, 68–73. [Google Scholar] [CrossRef]
- He, Z.; Zhu, H.; Xu, M.; Zeng, M.; Qin, F.; Chen, J. Complexation of bovine β-lactoglobulin with malvidin-3-O-glucoside and its effect on the stability of grape skin anthocyanin extracts. Food Chem. 2016, 209, 234–240. [Google Scholar] [CrossRef] [PubMed]
- Murphy, R.W.; Farkas, B.E.; Jones, O.G. Effect of crosslinking on the physical and chemical properties of beta-lactoglobulin (Blg) microgels. J. Colloid Interface Sci. 2017, 505, 736–744. [Google Scholar] [CrossRef] [PubMed]
- Jia, J.; Gao, X.; Hao, M.; Tang, L. Comparison of binding interaction between β-lactoglobulin and three common polyphenols using multi-spectroscopy and modeling methods. Food Chem. 2017, 228, 143–151. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Harris, P.; Kaur, A.; Pastrana, L.; Jauregi, P. Characterisation of β-lactoglobulin nanoparticles and their binding to caffeine. Food Hydrocoll. 2017, 71, 85–93. [Google Scholar] [CrossRef]
- Livney, Y.D. Milk proteins as vehicles for bioactives. Curr. Opin. Colloid Interface Sci. 2010, 15, 73–83. [Google Scholar] [CrossRef]
- Ren, Y.; Zhao, T.; Mao, G.; Zhang, M.; Li, F.; Zou, Y.; Yang, L.; Wu, X. Antitumor activity of hyaluronic acid–selenium nanoparticles in heps tumor mice models. Int. J. Biol. Macromol. 2013, 57, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Hou, D.; Lu, J.; Zhu, L.; Zhang, P.; Zhou, N.; Chen, K. Anti-tumor activity of exopolysaccharide from rhizopus nigricans ehrenb on s180 tumor-bearing mice. Bioorg. Med. Chem. Lett. 2016, 26, 2098–2104. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Park, G.; Lee, S.; Hwang, S.W.; Min, N.; Lee, K.M. Single wall carbon nanotube electrode system capable of quantitative detection of CD4+ T cells. Biosens. Bioelectron. 2017, 90, 238–244. [Google Scholar] [CrossRef] [PubMed]
- Hiraoka, N. Tumor-infiltrating lymphocytes and hepatocellular carcinoma: Molecular biology. Int. J. Clin. Oncol. 2010, 6, 544–552. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Sarkar, M.; Ghosh, T.; Guha, I.; Bhuniya, A.; Biswas, J.; Mallick, A.; Bose, A.; Baral, R. Absence of CD4+ T cell help generates corrupt CD8+ effector T cells in sarcoma-bearing Swiss mice treated with NLGP vaccine. Immunol. Lett. 2016, 175, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Godet, Y.; Fabre, E.; Dosset, M.; Lamuraglia, M.; Levionnois, E.; Ravel, P.; Benhamouda, N.; Cazes, A.; Le Pimpec-Barthes, F.; Gaugler, B.; et al. Analysis of spontaneous tumor-specific CD4 T-cell immunity in lung cancer using promiscuous hla-dr telomerase-derived epitopes: Potential synergistic effect with chemotherapy response. Clin. Cancer Res. 2012, 18, 2943–2953. [Google Scholar] [CrossRef] [PubMed]
- Oh, J.G.; Hwang, D.J.; Heo, T.H. Direct regulation of IL-2 by curcumin. Biochem. Biophys. Res. Commun. 2017, 495, 300–305. [Google Scholar] [CrossRef] [PubMed]
- Ruan, B.Y.; Chen, S.N.; Hou, J.; Huang, B.; Laghari, Z.A.; Li, L.; Nie, P. Two type II IFN members, IFN-γ and IFN-γ related (rel), regulate differentially IRF1 and IRF11 in zebrafish. Fish Shellfish Immunol. 2017, 65, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Umapathy, D.; Krishnamoorthy, E.; Mariappanadar, V.; Viswanathan, V.; Ramkumar, K.M. Increased levels of circulating (TNF-α) is associated with (-308G/A) promoter polymorphism of TNF-α gene in Diabetic Nephropathy. Int. J. Biol. Macromol. 2017, in press. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Not available. |

{kind=link}
| Treatment | Dose | Thymus Index | Spleen Index | Tumor Weight | Inhibitory Rate |
|---|---|---|---|---|---|
| (μg/kg) | (mg/g) | (mg/g) | (g) | (%) | |
| N | - | 3.21 ± 0.36 | 4.89 ± 0.31 | - | - |
| M | - | 1.63 ± 0.14 a | 8.16 ± 0.21 a | 2.79 ± 0.28 | - |
| β-lg | 150 | 1.98 ± 0.14 b | 6.47 ± 0.31 b | 2.19 ± 0.12 b | 21.51 |
| SeO2 | 100 | 1.33 ± 0.11 b | 3.17 ± 0.13 b | 1.31 ± 0.11 b | 53.05 |
| L-Se-β-lg | 50 | 1.94 ± 0.12 b | 6.48 ± 0.41 b | 2.21 ± 0.19 b | 20.79 |
| M-Se-β-lg | 100 | 2.76 ± 0.28 b | 5.38 ± 0.42 b | 1.51 ± 0.10 b | 45.88 |
| H-Se-β-lg | 150 | 3.18 ± 0.36 b | 4.72 ± 0.21 b | 1.44 ± 0.12 b | 48.38 |
| Treatment | Spleen | Thymus | ||
|---|---|---|---|---|
| CD4/(%) | CD8/(%) | CD4/(%) | CD8/(%) | |
| N | 67.01 ± 6.19 | 15.13 ± 1.21 | 17.07 ± 1.19 | 1 1.21 ± 1.23 |
| M | 50.21 ± 3.25 a | 8.52 ± 0.88 a | 28.21 ± 2.24 a | 21.52 ± 1.88 a |
| β-lg | 54.36 ± 4.12 b | 11.22 ± 1.73 b | 24.89 ± 2.12 b | 19.22 ± 1.73 b |
| SeO2 | 44.02 ± 3.07 b | 6.22 ± 0.82 b | 14.02 ± 1.07 b | 13.15 ± 1.22 b |
| L-Se-β-lg | 53.98 ± 4.16 | 12.44 ± 1.12 b | 23.98 ± 2.23 b | 21.44 ± 2.16 |
| M-Se-β-lg | 61.68 ± 3.41 b | 13.27 ± 1.14 b | 21.68 ± 2.41 b | 17.28 ± 1.23 b |
| H-Se-β-lg | 63.25 ± 5.25 b | 13.21 ± 1.19 b | 20.31 ± 2.25 b | 16.23 ± 1.48 b |
| Treatment | Peripheral Blood | Solid Tumor | ||
|---|---|---|---|---|
| CD4/(%) | CD8/(%) | CD4/(%) | CD8/(%) | |
| N | 42.22 ± 3.17 | 23.01 ± 2.19 | - | - |
| M | 28.63 ± 2.11 a | 18.52 ± 1.12 a | 0.09 ± 0.003 | 41.52 ± 3.12 |
| β-lg | 32.79 ± 3.25 b | 19.88 ± 1.14 b | 0.12 ± 0.003 | 40.24 ± 3.11 |
| SeO2 | 24.02 ± 2.11 b | 16.04 ± 1.12 b | 0.14 ± 0.002 | 38.41 ± 2.21 |
| L-Se-β-lg | 31.98 ± 2.20 b | 21.11 ± 1.16 b | 0.12 ± 0.004 | 39.87 ± 3.29 |
| M-Se-β-lg | 38.68 ± 3.21 b | 21.28 ± 2.19 b | 0.13 ± 0.003 | 41.01 ± 2.41 |
| H-Se-β-lg | 44.20 ± 3.24 b | 23.19 ± 1.27 b | 0.11 ± 0.004 | 38.82 ± 3.34 |
| Groups | G0/G1/(%) | S/(%) | G2/M/(%) | Apoptosis/(%) |
|---|---|---|---|---|
| M | 31.44 ± 2.35 | 54.51 ± 3.12 | 14.52 ± 1.63 | 2.24 ± 0.32 |
| β-lg | 42.32 ± 2.32 b | 44.63 ± 3.42 b | 14.34 ± 1.24 | 13.21 ± 1.05 b |
| SeO2 | 50.21 ± 3.32 b | 34.32 ± 2.15 b | 15.26 ± 1.22 | 27.15 ± 2.53 b |
| L-Se-β-lg | 38.13 ± 2.46 b | 48.27 ± 4.13 b | 14.47 ± 1.45 | 11.32 ± 1.03 b |
| M-Se-β-lg | 43.23 ± 2.12 b | 41.32 ± 3.23 b | 13.51 ± 1.63 | 18.13 ± 1.24 b |
| H-Se-β-lg | 49.11 ± 3.43 b | 36.92 ± 3.05 b | 15.37 ± 1.35 | 24.13 ± 2.12 b |
| Treatment | IL-2 (pg/mL) | IFN-γ (ng/L) | TNF-α (ng/L) |
|---|---|---|---|
| N | 4.01 ± 0.19 | 119.83 ± 7.01 | 117.89 ± 4.07 |
| M | 2.52 ± 0.11 a | 112.34 ± 5.93 a | 91.09 ± 5.99 a |
| β-lg | 2.89 ± 0.15 b | 116.71 ± 4.97 | 97.77 ± 7.03 |
| SeO2 | 2.02 ± 0.11 b | 100.84 ± 4.93 | 80.90 ± 4.99 |
| L-Se-β-lg | 2.98 ± 0.16 b | 118.8 ± 5.98 | 101.86 ± 4.04 b |
| M-Se-β-lg | 3.68 ± 0.21 b | 124.5 ± 8.03 b | 111.56 ± 3.09 b |
| H-Se-β-lg | 3.81 ± 0.25 b | 133.03 ± 7.07 b | 120.09 ± 6.13 b |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, S.-j.; Feng, Y.-y.; Zhang, Y.; Ji, H.-y.; Yu, J.; Liu, A.-j. Antitumor and Immunoregulatory Activities of Seleno-β-Lactoglobulin on S180 Tumor-Bearing Mice. Molecules 2018, 23, 46. https://doi.org/10.3390/molecules23010046
Sun S-j, Feng Y-y, Zhang Y, Ji H-y, Yu J, Liu A-j. Antitumor and Immunoregulatory Activities of Seleno-β-Lactoglobulin on S180 Tumor-Bearing Mice. Molecules. 2018; 23(1):46. https://doi.org/10.3390/molecules23010046
Chicago/Turabian StyleSun, Su-jun, Ying-ying Feng, Yan Zhang, Hai-yu Ji, Juan Yu, and An-jun Liu. 2018. "Antitumor and Immunoregulatory Activities of Seleno-β-Lactoglobulin on S180 Tumor-Bearing Mice" Molecules 23, no. 1: 46. https://doi.org/10.3390/molecules23010046