The Native Fruit Geoffroea decorticans from Arid Northern Chile: Phenolic Composition, Antioxidant Activities and In Vitro Inhibition of Pro-Inflammatory and Metabolic Syndrome-Associated Enzymes

 ,
,  and
and
Abstract
1. Introduction
2. Results and Discussion
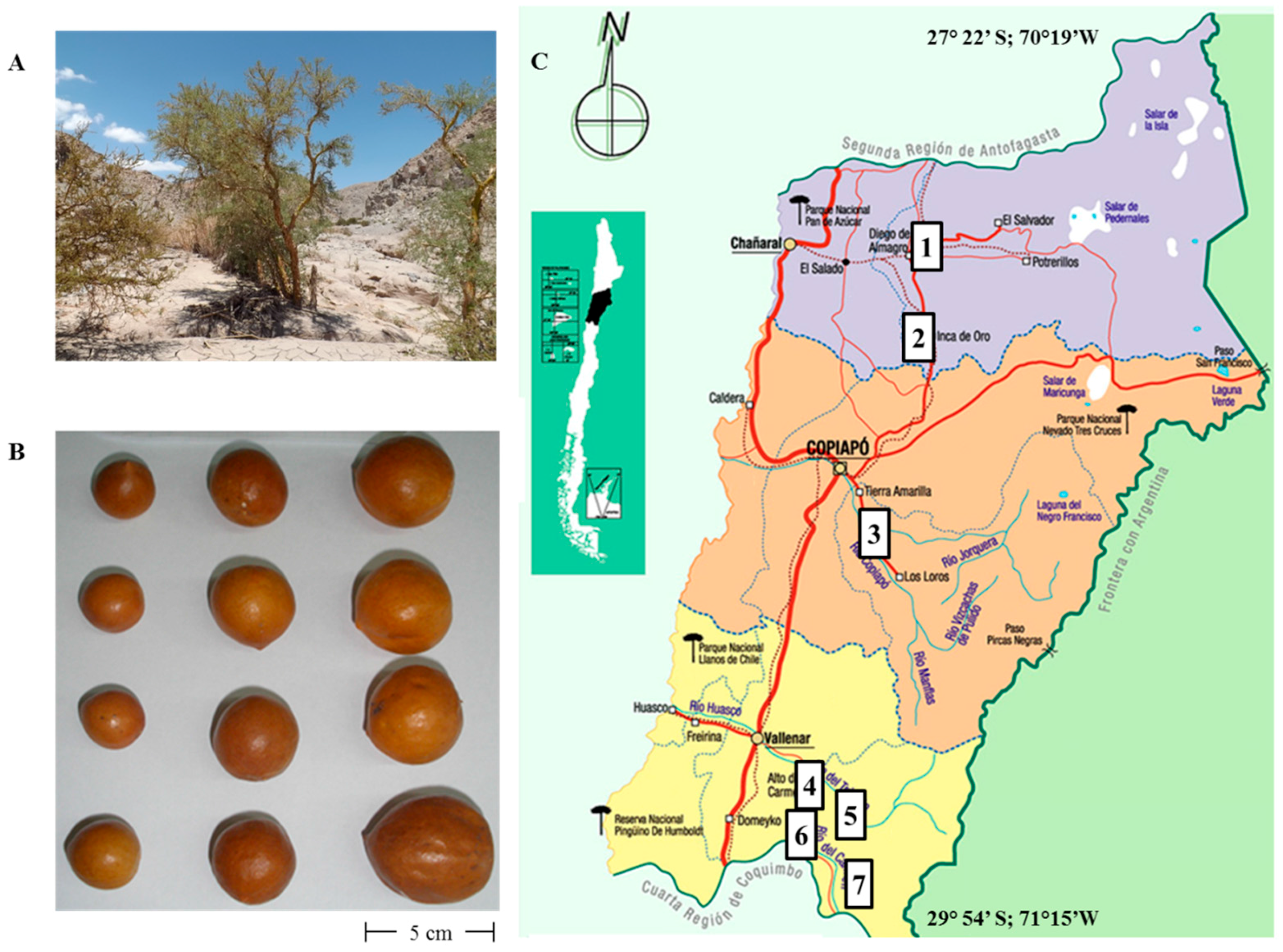
2.1. General Analysis of the Fruits
2.2. Total Phenolic (TP), Total Flavonoid (TF) and Total Proanthocyanidin (TPAC) Content
2.3. Antioxidant Activity
2.4. Inhibition of Pro-Inflammatory Enzymes
2.5. Inhibition of Metabolic Syndrome Associated-Enzymes
2.6. Tentative Identification of Phenolic Compounds in G. decorticans Fruits by HPLC-DAD and HPLC-ESI-MS/MS
2.6.1. Flavonoid Derivatives
2.6.2. Flavan-3-ol Monomers and Polymers
2.6.3. Phenolic Acids
2.7. Effect of Fractionation by Sephadex LH-20 on DPPH and α-Glucosidase Activity
2.8. Cytotoxicity Assay
2.9. Statistical Analyses
3. Materials and Methods
3.1. Chemicals
3.2. Sample Preparation
3.3. Total Phenolic (TP), Total Flavonoid (TF) and Total Proanthocyanidin (TPAC) Content
3.4. Antioxidant Activity
3.4.1. Reducing Power
3.4.2. Scavenging of Free Radicals
3.5. Inhibition of Pro-Inflammatory Enzymes
3.5.1. Lipooxygenase (LOX)
3.5.2. Cyclooxygenase (COX)
3.5.3. Secretory Phospholipase A2 (sPLA2)
3.6. Inhibition of Metabolic Syndrome Associated Enzymes
3.6.1. α-Glucosidase Inhibition Assay
3.6.2. α-Amylase Inhibition Assay
3.6.3. Lipase Inhibition Assay
3.7. HPLC-DAD and HPLC-MS Analysis
3.8. Fractionation of PEEs in Sephadex LH-20
3.9. Cytotoxicity Assay
3.10. Statistical Analyses
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Burkart, A. Las Leguminosas Argentinas Silvestres Y Cultivadas; ACME: Buenos Aires, Argentina, 1952; pp. 1–569. [Google Scholar]
- Rodríguez, R.; Matthei, O.; Quezada, M. Flora Arbórea De Chile; Editorial Universidad de Concepción: Concepción, Chile, 1983; pp. 1–408. [Google Scholar]
- Reynoso, M.A.; Vera, N.; Aristimuño, M.E.; Daud, A.; Sánchez Riera, A. Antinociceptive activity of fruits extracts and “arrope” of Geoffroea decorticans (chañar). J Ethnopharmacol. 2013, 145, 355–362. [Google Scholar] [CrossRef] [PubMed]
- Costamagna, M.S.; Zampini, I.C.; Alberto, M.R.; Cuello, S.; Torres, S.; Perez, J.; Quispe, C.; Schmeda-Hirschmann, G.; Isla, M.I. Polyphenol rich fraction from Geoffroea decorticans fruits flour affects key enzymes involved in metabolic syndrome, oxidative stress and inflammatory process. Food Chem. 2016, 190, 392–402. [Google Scholar] [CrossRef] [PubMed]
- Lamarque, A.L.; Maestri, D.M.; Zygadlo, J.A.; Guzmán, C.A. Chemical evaluation of Geoffroea decorticans seeds as source of oil and protein. Grasas Y Aceites 2000, 51, 241–243. [Google Scholar] [CrossRef]
- Vila, J.; Balderrama, L.; Bravo, J.L.; Almanza, G.; Codina, C.; Bastida, J.; Connolly, J. Prenylisoflavanones from Geoffroea decorticans. Phytochemistry 1998, 49, 2525–2528. [Google Scholar] [CrossRef]
- Jiménez-Aspee, F.; Quispe, C.; Soriano, M.D.P.C.; Fuentes Gonzalez, J.; Hüneke, E.; Theoduloz, C.; Schmeda-Hirschmann, G. Antioxidant activity and characterization of constituents in Copao fruits (Eulychnia acida Phil., Cactaceae) by HPLC-DAD-MS/MSn. Food Res. Int. 2014, 62, 286–298. [Google Scholar] [CrossRef]
- Schmeda-Hirschmann, G.; Quispe, C.; Soriano, M.D.P.C.; Theoduloz, C.; Jiménez-Aspee, F.; Pérez, M.J.; Cuello, A.S.; Isla, M.I. Chilean Prosopis mesocarp flour: Phenolic profiling and antioxidant activity. Molecules 2015, 20, 7017–7033. [Google Scholar] [CrossRef] [PubMed]
- Costamagna, M.S.; Ordoñez, R.M.; Zampini, I.C.; Sayago, J.E.; Isla, M.I. Nutritional and antioxidant properties of Geoffroea decorticans, an Argentinean fruit, and derived products (flour, arrope, decoction and hydroalcoholic beverage). Food Res. Int. 2013, 54, 160–168. [Google Scholar] [CrossRef]
- Godevac, D.; Zdunic, G.; Savikin, K.; Vajs, V.; Menkovic, N. Antioxidant activity of nine Fabaceae species growing in Serbia and Montenegro. Fitoterapia 2008, 79, 185–187. [Google Scholar] [CrossRef] [PubMed]
- Elmarakby, A.A.; Sullivan, J.C. Relationship between oxidative stress and inflammatory cytokines in diabetic nephropathy. Cardiovasc. Ther. 2012, 30, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Smith, W.L.; Garavito, R.M.; Dewitt, D.L. Prostaglandin endoperoxide H synthase (cyclooxygenase)-1 and -2. J. Biol. Chem. 1996, 271, 33157–33160. [Google Scholar] [CrossRef] [PubMed]
- Steele, V.E.; Holmes, C.A.; Hawk, E.T.; Kopelovich, L.; Lubet, R.A.; Crowell, J.A.; Sigman, C.C.; Kelloff, G.J. Lipoxygenase inhibitors as potential cancer chemopreventives. Cancer Epidemiol. Biomark. Prev. 1999, 8, 467–483. [Google Scholar]
- Fiorucci, S.; Antonelli, E.; Burgaud, J.L.; Morelli, A. Nitric oxide-releasing NDAIDs: A review of their current status. Drug. Saf. 2001, 24, 801–811. [Google Scholar] [CrossRef] [PubMed]
- Yin, Z.; Zhang, W.; Feng, F.; Zhang, Y.; Kang, W. α-Glucosidase inhibitors isolated from medicinal plants. Food Sci. Hum. Wellness 2014, 3, 136–174. [Google Scholar] [CrossRef]
- Kadouh, H.C.; Sun, S.; Zhu, W.; Zhou, K. α-Glucosidase inhibiting activity and bioactive compounds of six red wine grape pomace extracts. J. Funct. Foods 2016, 26, 577–584. [Google Scholar] [CrossRef]
- Li, X.M.; Sui, Y.; Li, S.; Xie, B.; Sun, Z. A-type procyanidins from litchi pericarp ameliorate hyperglycaemia by regulating hepatic and muscle glucose metabolism in streptozotocin (STZ)-induced diabetic mice fed with high fat diet. J. Funct. Foods 2016, 27, 711–722. [Google Scholar] [CrossRef]
- Zhang, B.; Deng, Z.; Ramdath, D.D.; Tang, Y.; Chen, P.X.; Liu, R.; Liu, Q.; Tsao, R. Phenolic profiles of 20 Canadian lentil cultivars and their contribution to antioxidant activity and inhibitory effects on α-glucosidase and pancreatic lipase. Food Chem. 2015, 172, 862–872. [Google Scholar] [CrossRef] [PubMed]
- Bellesia, A.; Tagliazucchi, D. Cocoa brew inhibits in vitro α-glucosidase activity: The role of polyphenols and high molecular weight compounds. Food Res. Int. 2014, 63, 439–445. [Google Scholar] [CrossRef]
- Calle, E.; Thun, M.J.; Petrelli, J.M.; Rodriguez, C.; Heath, C.W. Body mass index and mortality in a prospective cohort of US adults. N. Engl. J. Med. 1999, 341, 1097–1105. [Google Scholar] [CrossRef] [PubMed]
- Birari, R.B.; Bhutani, K.K. Pancreatic lipase inhibitors from natural sources: Unexplored potential. Drug Discov. Today 2007, 12, 879–889. [Google Scholar] [CrossRef] [PubMed]
- Rahim, A.T.M.A.; Takahashi, Y.; Yamaki, K. Mode of pancreatic lipase inhibition activity in vitro by some flavonoids and non-flavonoid polyphenols. Food Res. Int. 2015, 75, 289–294. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.X.; Liu, J.; Cai, S.B.; Wang, O.; Ji, B.P. Synergistic interactions of apigenin, naringin, quercetin and emodin on inhibition of 3T3-L1 preadipocyte differentiation and pancreas lipase activity. Obes. Res. Clin. Pract. 2016, 10, 327–339. [Google Scholar] [CrossRef] [PubMed]
- Abu-Reidah, I.M.; Arráez-Román, D.; Segura-Carretero, A.; Fernández-Gutiérrez, A. Profiling of phenolic and other polar constituents from hydro-methanolic extract of watermelon (Citrullus lanatus) by means of accurate-mass spectrometry (HPLC-ESI-QTOF-MS). Food Res. Int. 2013, 51, 354–362. [Google Scholar] [CrossRef]
- Fabre, N.; Rustan, I.; de Hoffmann, E.; Quetin-Leclercq, J. Determination of flavone, flavonol, and flavanone aglycones by negative ion liquid chromatography electrospray ion trap mass spectrometry. J. Am. Soc. Mass Spectrom. 2001, 12, 707–715. [Google Scholar] [CrossRef]
- Surco-Laos, F.; Dueñas, M.; González-Manzano, S.; Cabello, J.; Santos-Buelga, C.; González-Paramás, A.M. Influence of catechins and their methylated metabolites on lifespan and resistance to oxidative and thermal stress of Caenorhabditis elegans and epicatechin uptake. Food Res. Int. 2012, 46, 514–521. [Google Scholar] [CrossRef]
- Lin, L.Z.; Sun, J.; Chen, P.; Monagas, M.J.; Harnly, J.M. UHPLC-PDA-ESI/HRMSn profiling method to identify and quantify oligomeric proanthocyanidins in plant products. J. Agric. Food Chem. 2014, 62, 9387–9400. [Google Scholar] [CrossRef] [PubMed]
- Li, H.J.; Deinzer, M.L. The mass spectral analysis of isolated hops A-type proanthocyanidins by electrospray ionization tandem mass spectrometry. J. Mass Spectrom. 2008, 43, 1353–1363. [Google Scholar] [CrossRef] [PubMed]
- Clifford, M.N.; Johnston, K.L.; Knight, S.; Kuhnert, N. Hierarchical scheme for LC-MSn identification of chlorogenic acids. J. Agric. Food Chem. 2003, 51, 2900–2911. [Google Scholar] [CrossRef] [PubMed]
- Montagut, G.; Baiges, I.; Valls, J.; Terra, X.; del Bas, J.M.; Vitrac, X.; Richard, T.; Mérillon, J.M.; Arola, L.; Blay, M.; et al. A trimer plus a dimer-gallate reproduce the bioactivity described for an extract of grape seed procyanidins. Food Chem. 2009, 116, 265–270. [Google Scholar] [CrossRef]
- Hughey, C.A.; Straut, C.; Howell, A.; Dai, Z.; Kaletunc, G. Inhibition of α-amylase and α-glucoamylase by tannins extracted from cocoa, pomegranates, cranberries, and grapes. J. Agric. Food Chem. 2013, 61, 1477–1486. [Google Scholar]
- Zhou, H.C.; Lin, Y.M.; Wei, S.D.; Tam, N.F. Structural diversity and antioxidant activity of condensed tannins fractionated from mangosteen pericarp. Food Chem. 2011, 129, 1710–1720. [Google Scholar] [CrossRef]
- Wallace, T.C.; Giusti, M.M. Evaluation of parameters that affect the 4-dimethylaminocinnamaldehyde assay for flavonols and proanthocyanidinds. J. Food Sci. 2010, 75, C619–C625. [Google Scholar] [CrossRef] [PubMed]
- Apak, R.; Güçlü, K.; Özyürek, M.; Bektaşoǧlu, B.; Bener, M. Cupric ion reducing antioxidant capacity assay for food antioxidants: Vitamins, polyphenolic, and flavonoids in food extracts. In Advanced Protocols in Oxidative Stress I; Armstrong, D., Ed.; Humana Press: New York, NY, USA, 2008; pp. 163–193. [Google Scholar]
- Cheel, J.; Theoduloz, C.; Rodríguez, J.A.; Caligari, P.D.S.; Schmeda-Hirschmann, G. Free radical scavenging activity and phenolic content in achenes and thalamus from Fragaria chiloensis spp. chiloensis, F. vesca and F. x ananassa cv. Chandler. Food Chem. 2007, 102, 36–44. [Google Scholar] [CrossRef]
- Wangensteen, H.; Samuelsen, A.B.; Malterud, K.E. Antioxidant activity in extracts from coriander. Food Chem 2004, 88, 293–297. [Google Scholar] [CrossRef]
- Khanum, S.A.; Murari, S.K.; Vishwanth, B.S.; Shashikanth, S. Synthesis of benzoyl phenyl benzoates as effective inhibitors for phospholipase A2 and hyaluronidase enzymes. Bioorg. Med. Chem. Lett. 2005, 15, 4100–4104. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.; Chang, S.K.C.; Zhang, Y. Comparison of α-amylase, α-glucosidase and lipase inhibitory activity of the phenolic substances in two black legumes of different genera. Food Chem. 2017, 214, 259–268. [Google Scholar] [CrossRef] [PubMed]
- McDougall, G.J.; Kulkarni, N.N.; Stewart, D. Berry polyphenols inhibit pancreatic lipase activity in vitro. Food Chem. 2009, 115, 193–199. [Google Scholar] [CrossRef]
- Cheli, F.; Baldi, A. Nutrition-based health: Cell-based bioassays for food antioxidant activity evaluation. J. Food Sci. 2011, 76, R197–R205. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the polyphenol-enriched extracts are available from the authors. |



{kind=link}
{kind=link}
| Collection Place | Pulp (%) | MeOH (%) | PEE (%) | TP (g GAE/kg PEE) | TF (g CE/kg PEE) | TPAC (g CE/kg PEE) | FRAP (mmol TE/g PEE) | CUPRAC (mmol TE/g PEE) | DPPH SC50 (μg/mL) | O2 Scavenging SC50 (μg/mL) # |
|---|---|---|---|---|---|---|---|---|---|---|
| Provincia de Chañaral | ||||||||||
| Diego de Almagro | 78.1 | 43.6 | 1.7 | 389.1 ± 2.0 a | 222.1 ± 1.7 a | 24.5 ± 2.7 a | 1.8 ± 0.1 a | 4.1 ± 0.1 a | 12.1 ± 0.1 a | 36.6 ± 0.9 a |
| Inca de Oro | 58.3 | 29.1 | 0.8 | 639.2 ± 11.1 b | 260.5 ± 8.9 b | 84.1 ± 7.4 b | 3.1 ± 0.1 b | 6.7 ± 0.3 b | 5.3 ± 0.7 b | 18.1 ± 1.0 b |
| Provincia de Copiapó | ||||||||||
| Copiapó (turning) | 66.6 | 51.7 | 1.8 | 450.0 ± 4.6 c | 55.8 ± 2.1 c,d | 90.1 ± 4.7 b | 2.9 ± 0.1 b | 7.4 ± 0.1 c | 4.9 ± 0.9 b | 15.2 ± 1.2 b,c |
| Copiapó (ripe) | 67.4 | 52.7 | 1.6 | 446.3 ± 2.8 c | 50.3 ± 2.4 c | 90.2 ± 4.1 b | 2.8 ± 0.2 b | 6.3 ± 0.1 b | 3.9 ± 0.5 b | 30.1 ± 0.9 d |
| Provincia de Huasco | ||||||||||
| Alto del Carmen | 59.0 | 43.9 | 4.0 | 508.3 ± 4.1 d | 37.8 ± 1.6 e | 123.4 ± 1.1 c | 3.0 ± 0.2 b | 6.7 ± 0.1 b | 3.9 ± 0.7 b | 14.5 ± 0.3 c |
| El Transito (1) | 73.2 | 45.9 | 1.2 | 236.4 ± 2.2 e | 163.5 ± 1.4 f | 11.7 ± 1.8 d | 1.6 ± 0.1 a,c | 3.4 ± 0.1 d | 10.4 ± 0.5 a | 44.2 ± 0.4 e |
| El Transito (2) | 67.7 | 53.8 | 0.9 | 361.0 ± 0.0 f | 77.1 ± 2.5 g | 17.7 ± 3.4 a,d | 1.4 ± 0.1 c | 1.3 ± 0.1 e | 19.3 ± 0.9 c | 45.1% ± 3.5 # |
| Pinte | 74.3 | 69.2 | 2.0 | 369.1 ± 0.0 f | 65.1 ± 2.6 d,h | 48.3 ± 1.2 e | 1.9 ± 0.1 a | 5.5 ± 0.2 f | 5.2 ± 0.6 b | 32.5 ± 2.1 d |
| Conay | 77.0 | 59.4 | 0.7 | 196.2 ± 1.5 g | 116.0 ± 3.0 i | BDL | 0.9 ± 0.1 d | 2.5 ± 0.1 g | 24.3 ± 0.3 d | n.d. |
| Catechin * | 5.4 ± 0.1 | 13.4 ± 0.3 | 11.4 ± 1.6 | 8.7 ± 0.1 |
| Collection Place | LOX IC50 (μg PEE/mL) | COX-1% Inhibition | COX-2% Inhibition | sPLA2% Inhibition or IC50 (μg PEE/mL) | α-Glucosidase IC50 (µg PEE/mL) | Lipase% of Inhibition or IC50 (µg PEE/mL) |
|---|---|---|---|---|---|---|
| Provincia de Chañaral | ||||||
| Diego de Almagro | 60.1 ± 3.2 a,b | 61.8 ± 1.3 a | 60.5 ± 1.2 a | 98.9 ± 0.8 a | 4.7 ± 0.0 a | 17.1 ± 0.9 # |
| Inca de Oro | 61.2 ± 4.4 a,b | 80.9 ± 1.2 b | 18.9 ± 1.2 b | 142.9 ± 4.9 b | 4.5 ± 0.3 a | 9.9 ± 0.7 a |
| Provincia de Copiapó | ||||||
| Copiapó (turning) | 51.8 ± 1.6 a,c | 74.8 ± 1.5 c | 12.9 ± 0.2 c | 38.1 ± 1.9 # | 2.1 ± 0.1 b | 66.0 ± 1.9 b |
| Copiapó (ripe) | 53.1 ± 1.7 a,b | 92.1 ± 1.3 d | 55.5 ± 0.6 d | 42.8 ± 1.5 # | 0.8 ± 0.0 c | 29.7 ± 3.0 # |
| Provincia de Huasco | ||||||
| Alto del Carmen | 43.6 ± 3.2 c | Inactive | 25.9 ± 0.6 e | 156.0 ± 1.2 c | 0.7 ± 0.1 c | 14.2 ± 0.1 c |
| El Transito (1) | 74.0 ± 3.6 d | Inactive | 51.3 ± 0.4 f | 39.9 ± 0.1 # | 5.0 ± 0.3 a | 33.6 ± 3.4 # |
| El Transito (2) | >100 | 70.1 ± 1.2 e | 31.2 ± 0.6 g | 34.7 ± 0.1 # | 7.3 ± 0.4 d | 34.9 ± 3.5 # |
| Pinte | 96.8 ± 1.8 e | 87.3 ± 1.3 f | Inactive | 34.6 ± 0.3 # | 4.9 ± 0.1 a | 0.0 # |
| Conay | 76.8 ± 2.8 d,e | 80.8 ± 1.1 d,f | 76.0 ± 0.9 h | n.d. | n.d. | n.d. |
| Caffeic acid * | 37.2 ± 2.0 | |||||
| Nimesulide * | 100 | 100 | ||||
| Ursolic acid * | 26.7 ± 0.7# | |||||
| Acarbose * | 120.9 ± 2.0 | |||||
| Orlistat * | 0.04 ± 0.00 |
| Peak | Rt (min) | λ Max (nm) | [M − H]− (m/z) | MS/MS (m/z) | Tentative Identification |
|---|---|---|---|---|---|
| 1 | 22.1 | 307sh, 280 | 463 | 301 | Ellagic acid hexoside 1 |
| 2 | 26.0 | 280 | 433 | 301 | Ellagic acid pentoside 1 |
| 3 | 27.2 | 325, 298sh, 280 | 421 | 287, 151 | Eriodictyol pentoside |
| 4 | 34.7 | 280 | 451 | 289 | (epi)-catechin hexoside 1 |
| 5 | 41.1 | 280 | 451 | 289 | (epi)-catechin hexoside 2 |
| 6 | 43.9 | 274 | 577 | 559, 451, 425, 407, 289 | Procyanidin B-type dimer 1 [27] |
| 7 | 46.1 | 327sh, 280 | 465 | 303 | Taxifolin hexoside 1 |
| 8 | 46.3 | 280 | 451 | 289 | (epi)-catechin hexoside 3 |
| 9 | 47.6 | 283 | 329 | 167, 123 | Vanillic acid hexoside |
| 10 | 50.2 | 324sh, 298sh, 280 | 461 | 419, 401, 341, 299, 209, 167 | Vanillic acid hexoside pentoside |
| 11 | 52.0 | ND | 615 | 493, 405, 327, 285 | Kaempferol derivative |
| 12 | 56.0 | 279 | 577 | 471, 451, 425, 289 | Procyanidin B-type dimer 2 [27] |
| 13 | 56.1 | 279 | 863 | 711, 573, 451, 289 | Procyanidin A-type trimer 1 [28] |
| 14 | 57.4 | 327sh, 280 | 465 | 303 | Taxifolin hexoside 2 |
| 15 | 59.8 | 306sh, 280 | 449 | 287, 151 | Eriodictyol hexoside |
| 16 | 61.6 | 274 | 863 | 711, 695, 573, 451 | Procyanidin A-type trimer 2 [28] |
| 17 | 64.1 | 279 | 863 | 711, 573, 451 | Procyanidin A-type trimer 3 [28] |
| 18 | 64.4 | 339, 273 | 337 | 191 | 5-p-coumaroylquinic acid [29] |
| 19 | 66.2 | 279 | 1153 | 863, 575, 451, 289 | Procyanidin B-type tetramer [27] |
| 20 | 69.1 | 347, 280 | 771 | 609, 301 | Quercetin dihexoside rhamnoside 1 |
| 21 | 71.3 | 279 | 939 | 863, 573, 411, 289 | Procyanidin trimer derivative |
| 22 | 72.6 | 347, 280 | 741 | 579, 447, 285 | Kaempferol dihexoside pentoside |
| 23 | 73.3 | 343, 280 | 625 | 463, 301 | Quercetin dihexoside 2 |
| 24 | 73.5 | 350, 280 | 771 | 609, 463, 301 | Quercetin dihexoside rhamnoside |
| 25 | 73.8 | 373, 250 | 479 | 317 | Myricetin hexoside |
| 26 | 75.4 | 279 | 577 | 539, 449, 289 | Procyanidin B-type dimer 3 [28] |
| 27 | 75.6 | 347, 277 | 755 | 593, 575, 285 | Kaemperol hexoside rutinoside |
| 28 | 76.2 | 317sh, 280 | 565 | 433, 271 | Naringenin hexoside pentoside |
| 29 | 77.5 | 279 | 577 | 539, 423, 289 | Procyanidin B-type dimer 4 [28] |
| 30 | 78.2 | 317sh, 280 | 433 | 271 | Naringenin hexoside 1 |
| 31 | 78.4 | 316sh, 280 | 623 | 301 | Ellagic acid derivative |
| 32 | 79.1 | 355, 317sh, 280 | 595 | 301 | Quercetin pentoside hexoside 1 |
| 33 | 80.3 | 355, 255 | 771 | 609, 463, 301 | Quercetin dihexoside rhamnoside 2 |
| 34 | 80.5 | 365, 265 | 609 | 447, 429, 285 | Kaempferol dihexoside |
| 35 | 80.8 | 365, 280 | 623 | 285 | Kaempferol hexoside glucuronide |
| 36 | 81.6 | 365, 300sh, 260 | 609 | 429, 285 | Kaempferol dihexoside 2 |
| 37 | 81.6 | 300sh, 288 | 433 | 271 | Naringenin hexoside 2 |
| 38 | 84.1 | ND | 579 | 447, 285 | Kaempferol pentoside hexoside |
| 39 | 84.3 | 355, 255 | 609 | 463, 301 | Rutin a |
| 40 | 85.8 | 340, 267, 254 | 755 | 285, 241, 175 | Luteolin rhamnoside dihexoside |
| 41 | 86.5 | ND | 463 | 301 | Quercetin hexoside |
| 42 | 87.0 | 342, 268, 250 | 609 | 477, 315 | Isorhamnetin pentoside hexoside |
| 43 | 87.6 | 355, 255 | 477 | 301 | Quercetin glucuronide |
| 44 | 88.6 | ND | 579 | 447, 285 | Kaempferol hexoside pentoside 2 |
| 45 | 88.7 | 342, 268, 250 | 755 | 623, 315 | Isorhamnetin rutinoside pentoside |
| 46 | 89.0 | 342, 268, 250 | 609 | 477, 315 | Isorhamnetin pentoside hexoside 2 |
| 47 | 90.8 | 350, 268, 247 | 785 | 623, 315 | Isorhamnetin dihexoside rhamnoside |
| 48 | 93.4 | 320, 280 | 515 | 353, 191, 135 | 3,5-dicaffeoylquinic acid [29] |
| 49 | 93.1 | 343, 268 | 609 | 477, 315, 301 | Isorhamnetin pentoside hexoside 3 |
| 50 | 94.6 | 343, 270 | 623 | 315 | Isorhamnetin rutinoside |
| 51 | 96.9 | 340, 300sh, 280 | 447 | 285 | Kaempferol hexoside |
| 52 | 97.5 | ND | 461 | 285 | Kaempferol glucuronide |
| 53 | 98.4 | 330, 245 | 515 | 353, 173 | 4,5-dicaffeoylquinic acid [29] |
| Sample | Mass (mg) | % Fraction | DPPH (SC50, µg/mL) | α-Glucosidase (IC50, µg/mL) |
|---|---|---|---|---|
| PEE | 2520 | 100 | 12.1 ± 0.1 | 4.7 ± 0.1 |
| Fractions | ||||
| 1–9 | 68.9 | 4.4 | 15.0 ± 0.0 # | 8.0 ± 0.4 |
| 10–14 | 217.1 | 14.0 | 16.1 ± 0.0 # | 7.3 ± 0.1 |
| 15–19 | 659.3 | 42.6 | 87.1 ± 0.1 | 2.6 ± 0.1 |
| 20–25 | 183.3 | 11.8 | 37.8 ± 0.0 | 0.7 ± 0.1 |
| 26–27 | 43.1 | 2.8 | 32.9 ± 0.0 | 0.6 ± 0.0 |
| 28–30 | 97.8 | 6.3 | 28.4 ± 0.0 | 0.7 ± 0.1 |
| 31–33 | 58.1 | 3.8 | 29.2 ± 0.0 | 0.6 ± 0.0 |
| 34–37 | 78.1 | 5.0 | 27.7 ± 0.0 | 0.4 ± 0.0 |
| 38–42 | 57.4 | 3.7 | 24.5 ± 0.0 | 0.5 ± 0.0 |
| 43–44 | 5.6 | 0.4 | 24.1 ± 0.0 | 0.9 ± 0.0 |
| 45–46 | 21.5 | 1.4 | 28.8 ± 0.1 | 0.6 ± 0.0 |
| 47–48 | 58.8 | 3.8 | 40.7 ± 0.1 | 0.5 ± 0.0 |
| Acarbose * | 120.9 ± 2.0 | |||
| Catechin * | 11.4 ± 1.6 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiménez-Aspee, F.; Theoduloz, C.; Soriano, M.D.P.C.; Ugalde-Arbizu, M.; Alberto, M.R.; Zampini, I.C.; Isla, M.I.; Simirgiotis, M.J.; Schmeda-Hirschmann, G. The Native Fruit Geoffroea decorticans from Arid Northern Chile: Phenolic Composition, Antioxidant Activities and In Vitro Inhibition of Pro-Inflammatory and Metabolic Syndrome-Associated Enzymes. Molecules 2017, 22, 1565. https://doi.org/10.3390/molecules22091565
Jiménez-Aspee F, Theoduloz C, Soriano MDPC, Ugalde-Arbizu M, Alberto MR, Zampini IC, Isla MI, Simirgiotis MJ, Schmeda-Hirschmann G. The Native Fruit Geoffroea decorticans from Arid Northern Chile: Phenolic Composition, Antioxidant Activities and In Vitro Inhibition of Pro-Inflammatory and Metabolic Syndrome-Associated Enzymes. Molecules. 2017; 22(9):1565. https://doi.org/10.3390/molecules22091565
Chicago/Turabian StyleJiménez-Aspee, Felipe, Cristina Theoduloz, Maria Del Pilar C. Soriano, Maider Ugalde-Arbizu, Maria Rosa Alberto, Iris Catiana Zampini, Maria Inés Isla, Mario J. Simirgiotis, and Guillermo Schmeda-Hirschmann. 2017. "The Native Fruit Geoffroea decorticans from Arid Northern Chile: Phenolic Composition, Antioxidant Activities and In Vitro Inhibition of Pro-Inflammatory and Metabolic Syndrome-Associated Enzymes" Molecules 22, no. 9: 1565. https://doi.org/10.3390/molecules22091565
APA StyleJiménez-Aspee, F., Theoduloz, C., Soriano, M. D. P. C., Ugalde-Arbizu, M., Alberto, M. R., Zampini, I. C., Isla, M. I., Simirgiotis, M. J., & Schmeda-Hirschmann, G. (2017). The Native Fruit Geoffroea decorticans from Arid Northern Chile: Phenolic Composition, Antioxidant Activities and In Vitro Inhibition of Pro-Inflammatory and Metabolic Syndrome-Associated Enzymes. Molecules, 22(9), 1565. https://doi.org/10.3390/molecules22091565